坡向对高寒矿区渣山恢复植物生物量和养分繁殖分配的影响
2022-08-10蒋宏宇李长慧梁德飞
蒋宏宇,李长慧,孙 熠,梁德飞
(1. 青海大学省部共建三江源生态与高原农牧业国家重点实验室, 青海 西宁 810016;2. 青海大学生态环境工程学院,青海 西宁 810016;3. 青海大学农牧学院, 青海 西宁 810016)
繁殖分配作为植物关键生活史策略,是植物为了保证种群延续分配至繁殖体的资源比例[1]。不仅是种群、群落生态学研究中的核心,也是生物演替及生态系统功能维持的基础[2]。植物通过营养和繁殖体分配的权衡来提升自身竞争力以及优化当前和未来分配的权衡来达到种群最大化延续[3]。植物竞争优势度以及资源利用的有效性等过程都依赖于能量和养分的繁殖分配比例[4]。养分储备是决定生物量分配的前提[5],是植物生长适应生境的基础[6],然而养分繁殖分配过程得到的关注相对较少[7-8]。植物经过长期自然选择调节繁殖分配以最大化适应生境的胁迫[9-10]。尤其在气候条件恶劣的青藏高原,植物对能量和养分的权衡分配是适应区域生境的重要机制[11]。
地处祁连山南麓的青海江仓矿区是黄河重要支流大通河的发源地。然而经过多年的露天煤炭开采,大面积煤矸石堆积的渣山导致区域高寒草地严重受损,因此渣山生态恢复是保障区域生态系统安全以及黄河流域高质量发展的必要措施。矿山堆积、沉陷等过程会改变植物适应性,如禾本科植物短花针茅(Stipa breviflora)通过降低千粒重及繁殖分配比例来适应神木矿区的采煤沉陷[12]。青海江仓矿区由于地处高寒地区,恶劣的气候条件导致恢复植物因生长季短而难以完成正常的生活史[13]。植物向繁殖体养分分配增多有助于完成种子成熟等生活史过程[6],因此通过研究植物繁殖分配变异筛选适宜恢复植物可能是提升渣山生态恢复效率的有效措施。
同时渣山坡向的温度、水分和土壤等生境条件存在差异,决定了在恢复过程中需根据坡向变化来选择适宜的植物及措施。山西阳泉矿区植物物种多样性随煤矸石山坡向变化而变化[14],福建长汀矿区渣山阳坡的植物物种更丰富[15],青海江仓矿区阳坡和平地上植物平均高度和盖度高于阴坡[16]。植物还能通过繁殖体及营养体资源分配的权衡以适应坡向变化等生境变异[17-18],如祁连山霸王(Zygophyllum xanthoxylum)通过增大繁殖分配比例以适应北坡到南坡的生境变异[19]。植物将资源投入到繁殖体以适应低温[20],分配能量至叶片促进光合以应对光照条件不足[21],分配资源到根部以适应土壤养分的缺乏[22]。同时水分缺乏会使得禾草繁殖分配降低导致杂类草升高[23],而随着海拔升高,植物个体变小且繁殖分配升高[20]。资源向繁殖体分配的增大暗示了种子有更高的生存保障,向营养分配倾向意味着根部抗逆性的增强[24]。为此,本研究假设从渣山阴坡、平地到阳坡,光辐射增多、温度升高、风蚀效应和水分下降等生境变化使得恢复植物向繁殖体的生物量和养分分配增高,且不同植物对坡向有不同的响应。以3 种典型恢复植物老芒麦(Elymus sibiricus)、冷地早熟禾(Poa crymophila)和小花碱茅(Puccinellia tenuiflora)为研究对象,分析不同坡向下恢复植物生物量繁殖分配和养分繁殖分配的变化,为渣山植物引种筛选提供理论支撑,以提升高寒矿区渣山恢复效率。
1 材料与方法
1.1 研究位点概述
试验区位于青海省天峻县江仓矿区5 号井(38°03′32″N,99°26′57″E,海拔3 870 m),该区域属高原大陆性气候,年均气温−4.2 ℃,年均降水量477 mm,蒸发量1 049 mm,平均风速2.9 m·s-1。区域原生植被以矮生嵩草(Kobresia humilis)、华扁穗草(Blysmus sincompressus)和木里苔草(Carex muliensis)为优势种,伴有矮金莲花(Trollius farreri)、长毛风毛菊(Saussurea hieracioides)和湿生扁蕾(Gentianopsis paludosa)等常见种。土壤类型为沼泽草甸土,且常年冻土覆盖。近年来露天煤矿的大面积开采,大量煤矸石以及腐植表土、风化岩土的堆积形成渣山。
1.2 试验设计
2016 年6 月,在位于江仓矿区5 号井南渣山的山顶平地、阳坡(坡度13.8°,坡向南偏东30°)和阴坡(坡度18.3°,坡向北偏西12°)上分别布设试验,人工将地表较大的石砾捡除后将样地耙平;设置样地大小为10 m × 10 m,样地间距2 m,并重复3 次;播种草种为高寒区常见的恢复物种老芒麦、冷地早熟禾和小花碱茅;播种量为5 g·m−2,播种比例2 ∶ 1 ∶ 1,种子轻轻撒播于地表后轻耙地表并镇压踩实,以保证播种种子充分接触土壤;之后撒施有机肥(3 kg·m−2),再覆盖无纺布(20 g·m−2);播种完成后用围栏将样地封育以防止放牧破坏。
样地围封3 年后,于2019 年8 月在每个样地内分别随机选取每种恢复植物10 株,将地上部分剪下,装入冰盒后带回实验室。将每株植物的繁殖体、叶片和茎秆分开,并置于65 ℃烘箱中烘干至恒重。用万分之一天平(Sartorius BSA 224S,德国)称量每株植物繁殖体、叶片和茎秆的生物量,用以计算个体大小和生物量繁殖分配比例。同时在每个样地内随机选取3 个50 cm × 50 cm 样方,将样方中植物的地上部分齐地表剪下,并将3 种恢复植物区分开,分别称量每种植物的地上生物量(精确到0.000 1 g)。随后将繁殖体、叶片和茎秆分开,将其粉碎用以测定不同组织的氮、磷含量。植物取样完成后在每个样地用1 cm 直径的打孔器随机采集5 个0 − 5 cm表层渣山土壤并充分混合均匀后测定土壤理化性质。
植物全氮、全磷利用浓硫酸 + 双氧水消解,土壤全氮用浓硫酸 + 催化剂(硫酸钾 ∶ 硫酸铜 ∶ 硒粉 =100 ∶ 10 ∶ 1)消解,土壤全磷用浓硫酸 + 高氯酸消解,土 壤 速 效 氮 和 速 效 磷 用2 mol·L−1氯 化 钾 和0.5 mol·L−1碳酸氢钠浸提,之后均用流动分析仪测定(SEAL A++,德国)[25];土壤有机质采用浓硫酸 +重铬酸钾法测定[26]。
1.3 统计分析
生物量繁殖分配(biomass reproductive allocation,BRA)计算公式如下[27]:

式中:Cr表示特定物种繁殖体养分含量,Cl表示叶片养分含量,Cs表示茎秆养分含量。
单因素方差分析植物繁殖体、叶片、茎秆生物量和养分含量等功能性状之间的差异,相同物种在坡向间的差异,以及土壤理化性质在坡向间的差异;双因素方差分析坡向和物种对生物量繁殖分配、养分繁殖分配、个体大小和地上生物量的影响。线性回归分析繁殖分配之间以及与组织养分含量等功能性状之间的关系。使用SPSS 25.0 软件分析数据,Origin 2015 作图,显著性水平为P< 0.05。
2 结果与分析
2.1 土壤理化特征
矿区渣山阳坡土壤有机质含量显著高于阴坡(P<0.05);阳坡土壤全氮含量显著高于阴坡和平地(P<0.05);另外渣山坡向间土壤pH、全磷含量和速效养分含量无显著差异(P> 0.05) (表1)。
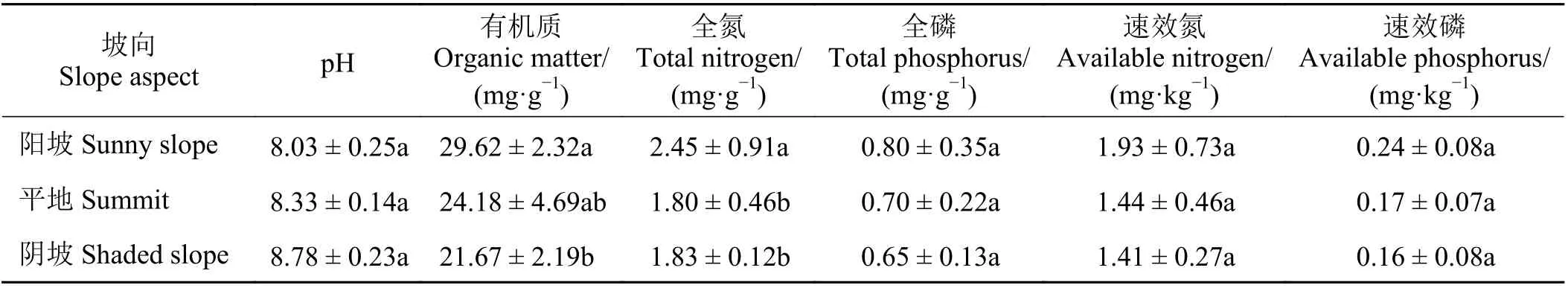
表1 不同坡向土壤理化特征Table 1 Soil physicochemical characteristics of different slope aspects
2.2 坡向对繁殖分配的影响
在阳坡上老芒麦和冷地早熟禾的繁殖体、叶片和茎秆生物量显著均高于阴坡(P< 0.05),而小花碱茅在阴坡上显著高于阳坡(P< 0.05);老芒麦和冷地早熟禾在阳坡的繁殖体氮含量显著高于阴坡(P<0.05);小花碱茅和冷地早熟禾在阳坡的繁殖体磷含量显著高于阴坡(P< 0.05) (表2)。另外,植物繁殖体氮含量高于叶片(P< 0.001),同时叶片氮含量显著高于茎秆(P= 0.008);繁殖体磷含量高于叶片和茎秆(P= 0.021)。
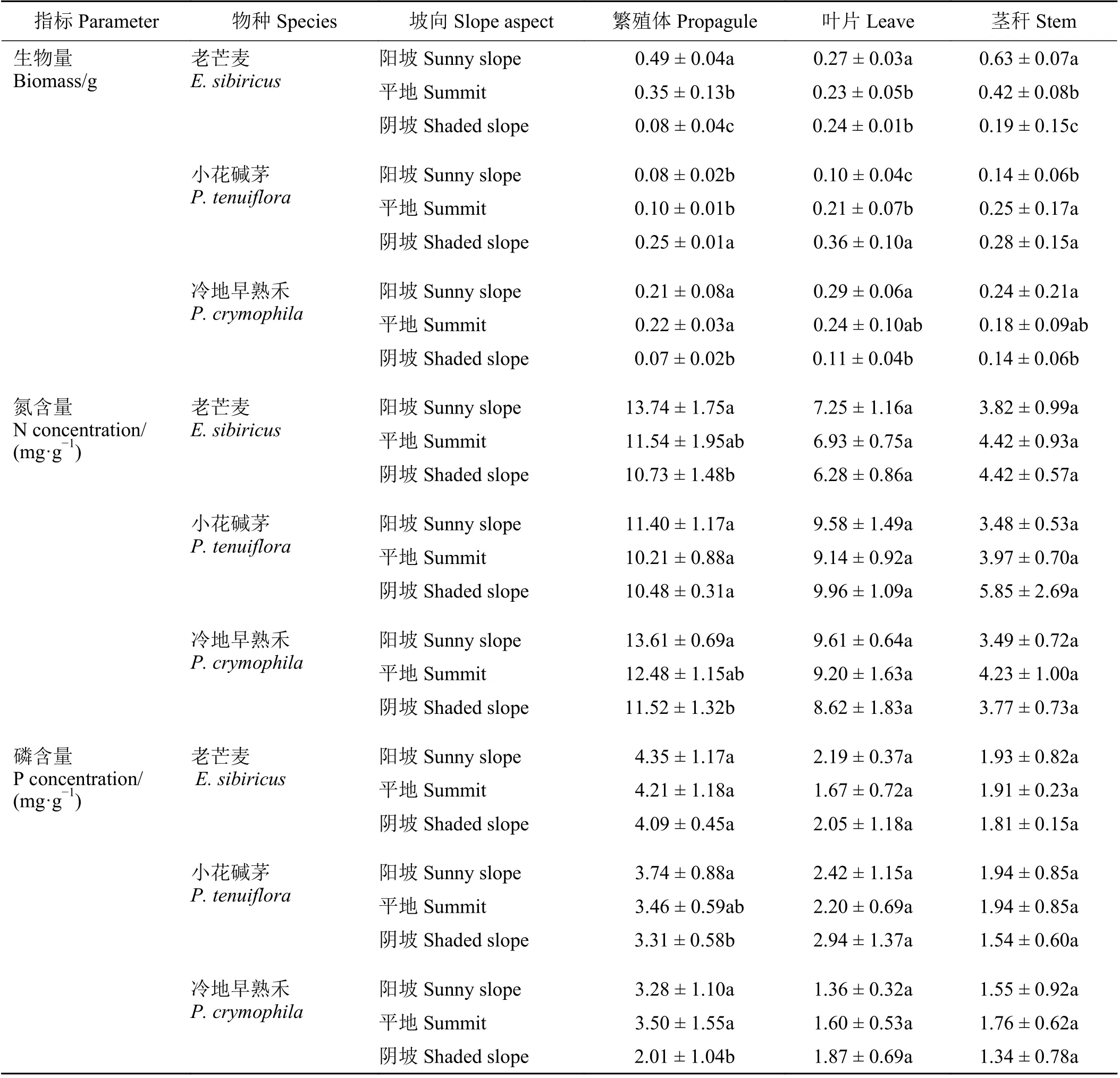
表2 恢复植物不同器官生物量和氮、磷在坡向间变化Table 2 Changes in biomass and concentrations of nitrogen and phosphorus of restoration plant tissues between different slope aspects
坡向对恢复植物生物量繁殖分配影响显著(P<0.05) (表3),阳坡平均生物量繁殖分配19.98%,显著高于阴坡13.25% (P< 0.05),平地介于二者之间(图1)。坡向对植物生物量繁殖分配的影响依赖于物种(表3),表现为在阳坡生物量繁殖分配从大到小依次为老芒麦、冷地早熟禾和小花碱茅,且彼此间差异显著(P< 0.05),平地上老芒麦和冷地早熟禾显著高于小花碱茅(P< 0.05),而在阴坡上小花碱茅显著高于老芒麦(P< 0.05) (图1)。

图1 坡向对恢复植物生物量繁殖分配的影响Figure 1 Effect of slope aspects on biomass reproductive allocation of restoration plants
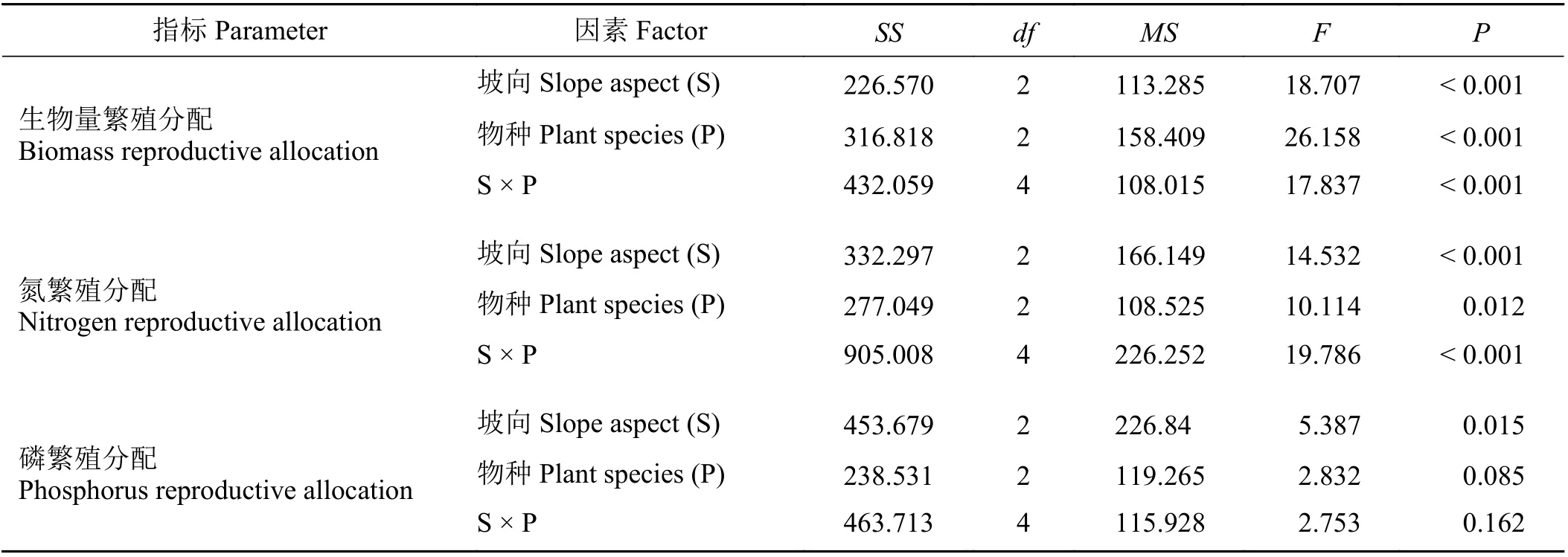
表3 渣山坡向和植物物种对生物量繁殖分配、氮繁殖分配和磷繁殖分配的影响Table 3 Two-way ANOVA analysis of the effect of slope aspect and plant species on biomass and nitrogen and phosphorus reproductive allocation
与生物量繁殖分配相似,坡向也改变了植物氮繁殖分配(表3),阳坡上平均氮繁殖分配33.96%,显著 高 于 阴 坡(25.51%) (P< 0.05),平 地 介 于 二 者 之间,且与二者无显著差异(P> 0.05) (图2);坡向、坡向及物种的交互作用对氮繁殖分配的影响显著(P<0.05) (表3),在阳坡和平地上,老芒麦和冷地早熟禾显著高于小花碱茅(P< 0.05),而在阴坡上小花碱茅显著高于老芒麦(P< 0.05) (图2)。
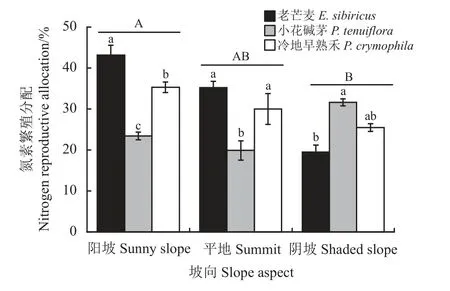
图2 坡向对恢复植物氮素繁殖分配的影响Figure 2 Effect of slope aspect on nitrogen reproductive allocation of restoration plants
坡向对植物磷繁殖分配影响显著(P< 0.05)(表3),从阳坡到阴坡,植物磷繁殖分配逐渐降低,且阳坡(34.83%)显著高于阴坡(25.40%) (P< 0.05)(图3)。

图3 坡向对恢复植物磷素繁殖分配的影响Figure 3 Effect of slope aspect on phosphorus reproductive allocation of restoration plants
从阳坡到阴坡,样方植物地上生物量显著下降(P= 0.031),且阳坡生物量显著高于阴坡和平地(P<0.05) (图4)。同时,坡向与物种对地上生物量的交互作用显著(P< 0.001),阳坡地上生物量以老芒麦最大,小花碱茅最小,冷地早熟禾介于二者之间,且与二者差异显著(P< 0.05);而在平地上地上生物量依次表现为冷地早熟禾 > 老芒麦 > 小花碱茅;在阴坡上冷地早熟禾和小花碱茅地上生物量显著高于老芒麦(P< 0.05) (图4)。
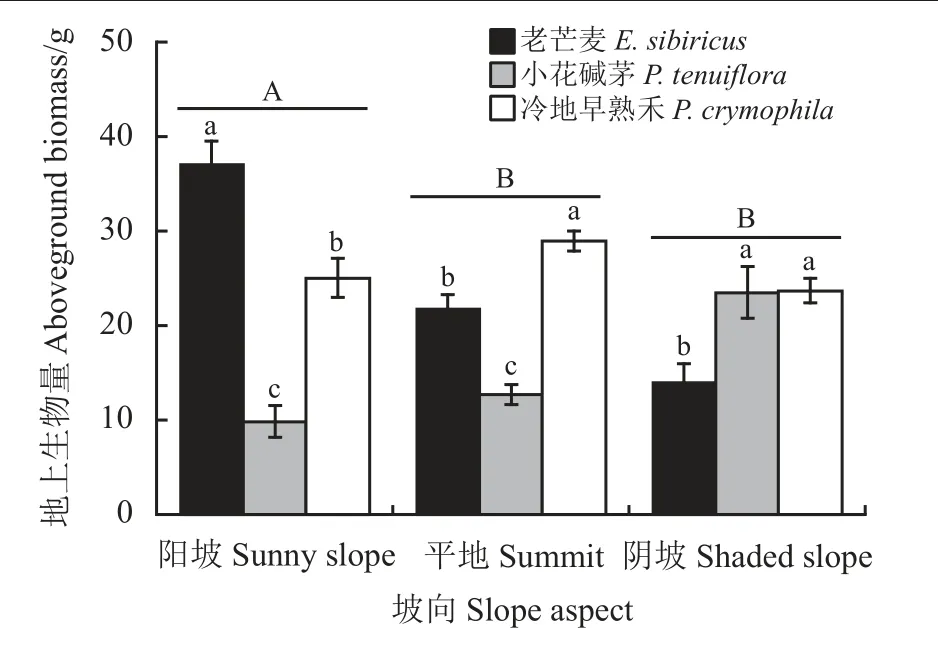
图4 坡向对恢复植物地上生物量的影响Figure 4 Effect of slope aspect on aboveground biomass of restoration plants
植物个体大小因坡向变化而显著变化(P=0.002),阴坡上植物个体最小,显著低于阳坡和平地(P< 0.05),阳坡和平地间无显著差异(P> 0.05);物种间个体大小存在不同差异,以老芒麦植株个体最大,显著高于小花碱茅和冷地早熟禾(P< 0.05)(图5)。
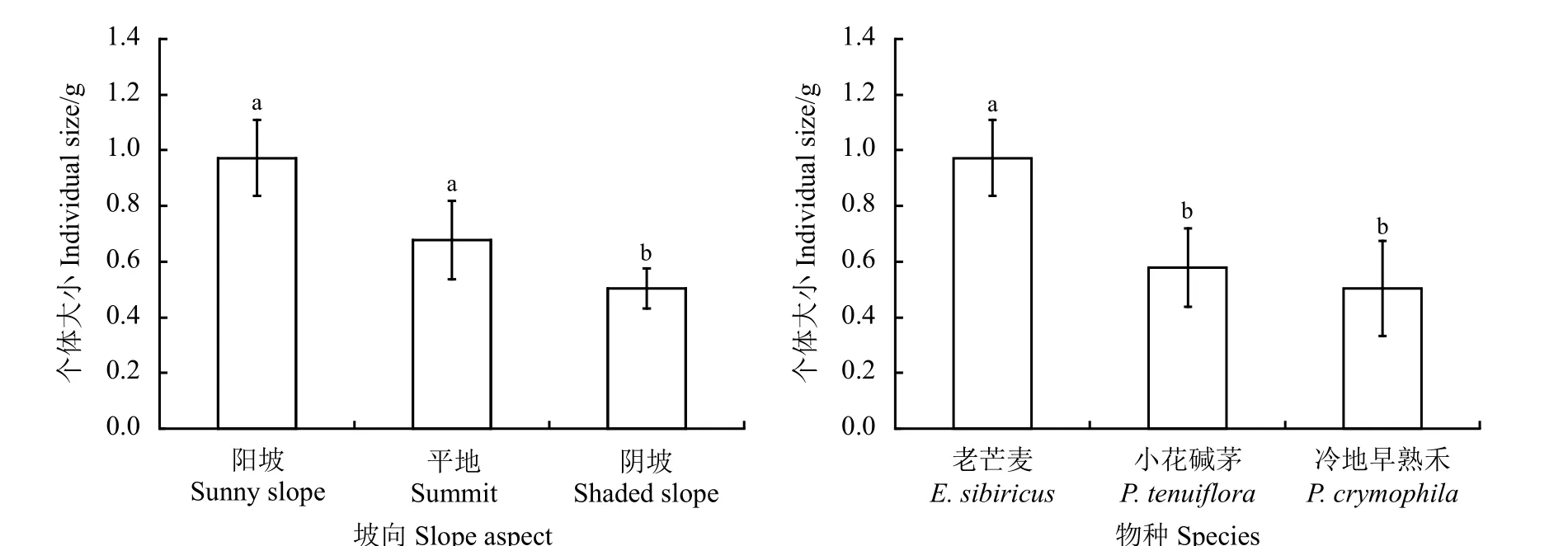
图5 坡向和物种对恢复植物个体大小的影响Figure 5 Effect of slope aspect and species on individual size of restoration plants
2.3 繁殖分配与植物功能性状的关系
植物繁殖体氮含量分别与地上生物量和氮素繁殖分配,以及繁殖体磷含量和磷繁殖分配呈显著正相关关系(图6)。表现为随着植物繁殖体含量的增高,生物量和氮素繁殖分配显著升高;随着繁殖体磷含量的升高,磷繁殖分配显著升高。另外植物养分繁殖分配比例均高于生物量繁殖分配,平均氮繁殖分配为29.27%,磷平均繁殖分配31.11%,二者均显著高于生物量繁殖分配的17.27% (P< 0.001)。
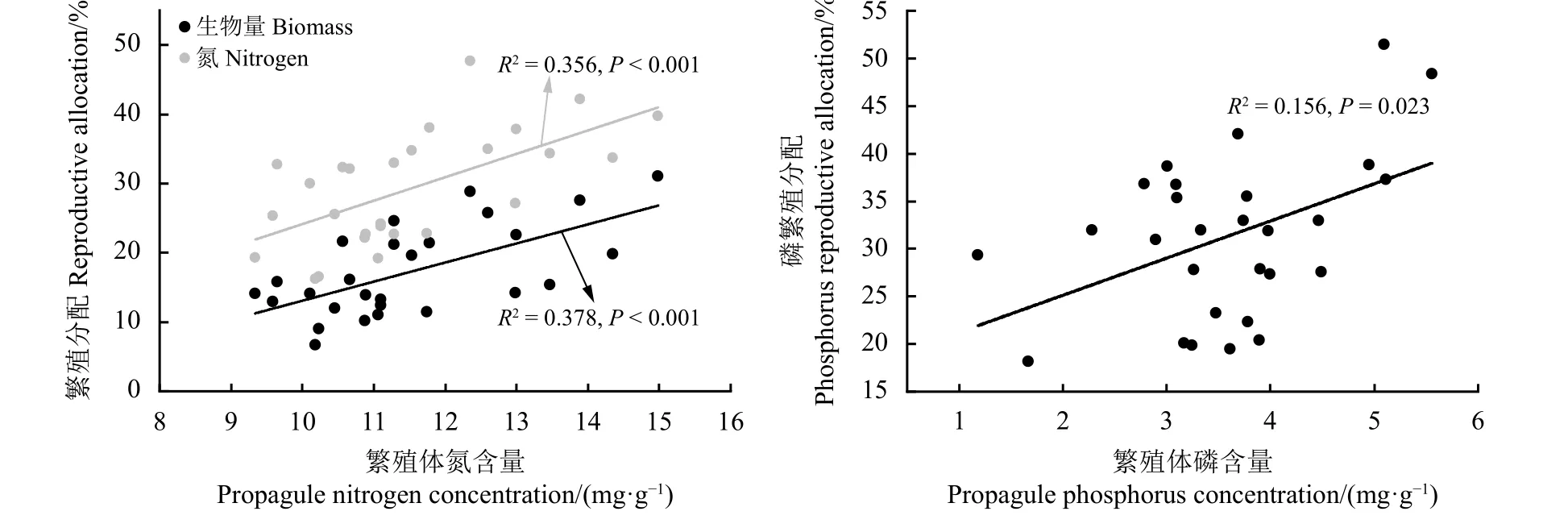
图6 繁殖体养分含量与繁殖分配的线性回归关系Figure 6 Linear regression relationships between propagule nutrient concentration and reproductive allocation
随着样方地上生物量的增加,生物量繁殖分配和氮繁殖分配显著升高,随着植物个体越大,生物量、氮和磷繁殖分配均显著升高(图7)。此外,随着生物量繁殖分配的增多,氮、磷繁殖分配显著升高,同时磷繁殖分配也随氮繁殖分配的升高而显著升高(图8)。

图7 繁殖分配与地上生物量及植物个体大小的线性回归关系Figure 7 Linear regression relationships between reproductive allocation and aboveground biomass and individual size
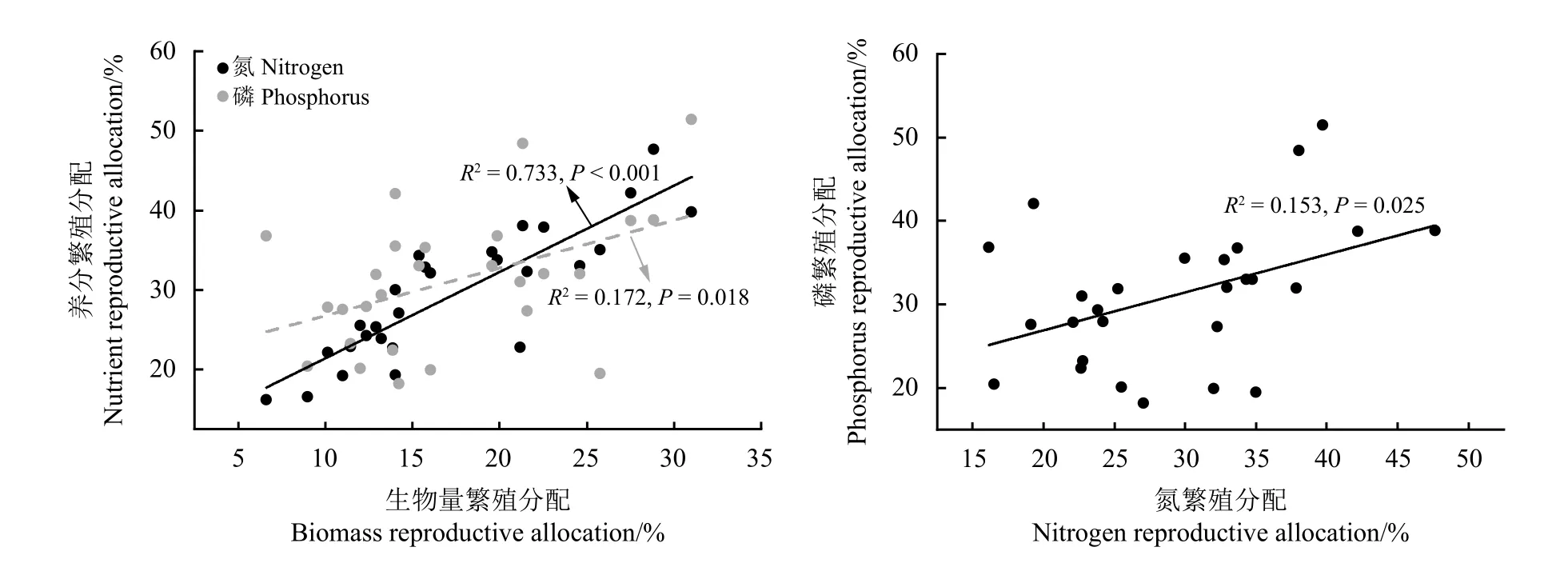
图8 生物量、氮和磷繁殖分配相互之间的线性回归关系Figure 8 Linear regression relationships between biomass and nitrogen and phosphorus reproduction allocation
3 讨论与结论
植物将资源向繁殖体和营养器官的分配权衡是适应生境的重要过程[29]。特别是高海拔区域高寒、低氧的气候特征,以及煤炭开采堆积形成的煤矸石渣山结构不稳定等生境条件决定了引种植物生活史极短,难以保证种子成熟及根部返青的资源储备[13],因此如何通过资源的合理分配以提高存活概率是植物在矿区渣山生长的重要机制。
恢复植物生物量和养分繁殖分配比例从阴坡到阳坡逐渐升高,可能是因为渣山坡向生境通过改变植物器官养分含量而改变了繁殖分配过程。从渣山阴坡、平地到阳坡,恢复植物繁殖体氮、磷含量逐渐升高,而叶片氮、磷含量没有显著变化。植物依据生境资源异质性变化而表现出多样的功能分化[30],组织养分储备丰富意味着生物量等资源分配倾向[31],通过权衡营养生长和繁殖分配比例,提升种群适合度等[32-33]。因此,随着植物繁殖体氮含量的升高生物量及氮繁殖分配也显著升高,且繁殖体磷含量的升高促进了磷繁殖分配的增高。虽然从阴坡到阳坡,渣山土壤总氮和有机质积累逐渐增多,但是在统计中与植物繁殖分配没有显著相关性,这可能是由于播种时间较短,恢复植物尚未能对土壤理化变异做出响应。另外从渣山阴坡、平地到阳坡,接收到的光辐射能量升高,植物光合效率增大,有助于个体生物量积累[34];同时也促进了养分积累及循环过程[35],植物生长的温度及养分限制的缓解提升了地上生产力及碳积累水平[36]。植物个体大小、地上生物量与繁殖分配显著的正相关关系也说明植物根据生境变异在保障当前生长的基础上才能进行繁殖体资源积累[37]。研究发现个体生长与繁殖体生长的异速关系,使得大个体比小个体植株能产生更多的繁殖体,甚至个体大小对繁殖分配的影响比生境及资源竞争等作用更大[38],因此渣山坡向通过改变植物个体大小而间接地促进繁殖分配。也有大量研究表明植物个体大小和繁殖分配呈显著的负相关性[39-40],和本研究结果不一致可能是因为矿区渣山是人工采矿的煤矸石堆积而成,形成时间短且结构不稳定,人工建立系统的能量流动和养分循环过程与历经千万年进化的自然系统不一致[41]。相对于生物因子,温度和水分等非生物因素可能是影响渣山植物繁殖分配等过程的关键因素[42]。个体大小和株丛数的地上生物量与生物量和氮素繁殖分配显著正相关(图7),说明渣山坡向引起的个体大小和密度变化改变了植物繁殖分配过程[5]。在胁迫较低的阳坡,植物将更多的资源分配至繁殖体,未来生长的权衡分配占比增大[4];相反的,在作为迎风坡的阴坡,温度低的同时风吹干扰也更剧烈,植物只能以牺牲繁殖体分配为代价,将资源更多的分配于适应当前生长的营养体[9]。
不同恢复植物对坡向的响应幅度不一致,表现在生境胁迫较弱的阳坡和平地上老芒麦和冷地早熟禾具有较高的生物量和氮繁殖分配,而在阴坡上小花碱茅具有更高的生物量和氮繁殖分配(图1、图2)。前人研究发现小花碱茅种子发芽率需保障足够的水分供应而对温度的要求不高,且随土壤pH的降低而降低[43],同时还具有较高的耐盐碱性[44]。以上特性可能是小花碱茅在渣山阴坡生长较好的原因,潜在的生理生化机制还需在后续研究中深入探讨。坡向与物种之间显著的交互作用说明在未来高寒矿区渣山恢复中,阳坡和平地上老芒麦和冷地早熟禾适宜引种,而在阴坡上小花碱茅有更高的适应性。另外坡向与物种间的交互作用对磷繁殖分配无显著影响说明复绿植物对氮更敏感。在高寒矿区渣山上,影响植物生长最主要营养元素可能是氮,这一结果也印证了中国北方草地生态系统植物生长主要受氮限制[45]。
引种植物平均生物量繁殖分配高寒区禾本科物种更低[46-47],可能是因为渣山周边是以莎草科物种为优势的沼泽化草甸,而引种植物是禾本科物种,植物与传粉者之间的不匹配致使协同进化关系紊乱[48],进而导致繁殖分配降低;同时多年生草本根茎繁殖比有性生殖更容易适应高寒、低氧且基质不稳定等生境胁迫[32]。因此在高寒低氧且养分匮乏的矿区渣山,恢复植物资源权衡分配优先供给当下,通过降低繁殖体分配而提升生境适宜性。另外本研究不足之处在于未能包含植物地下生物量和养分,未来研究不仅要考虑地下过程,还需将植物化学计量、生理生化等特征综合分析,为高寒矿区的生态恢复提供支撑。
综上,高寒矿区渣山坡向改变了恢复植物繁殖分配过程。从阴坡、平地到阳坡,繁殖体养分含量增大,植物个体大小、地上生物量以及繁殖分配比例均显著升高。且物种对坡向表现出不同的响应,在阳坡和平地上老芒麦和冷地早熟禾的生物量和氮繁殖分配更高,而在阴坡上小花碱茅更高。后续高寒矿区渣山恢复中,需根据渣山坡向差异以及恢复植物繁殖分配权衡特征选择适宜恢复措施,从而提升恢复效果。
