兰花型、蒲公英型还是郁金香型?环境与神经生物特征对儿童发展的影响
2022-08-10孙芯芸陈矜之计邹慧
王 燕 孙芯芸 陈矜之,2 计邹慧
(1 复旦大学心理学系,上海 200433)(2 台州学院心理健康教育与咨询中心, 台州 317000)
1 前言
传统的心理学通常聚焦于环境对个体发展的影响,大量文献论证了环境在塑造个体发展中的主导作用。然而,个体并不只是被动地被环境所左右,其自身特征也会在一定程度上左右环境所释放的影响力,个体同环境之间也存在着交互作用。
进入21世纪之后,随着表观遗传学和脑科学的发展,研究者开始日益关注个体自身的神经生物学变量对环境影响力的调节作用,并相继提出不同的模型假说,其中较有影响的有素质压力模型/双重风险模型、环境的生物敏感性理论、差别易感性模型以及优势敏感性假说。围绕这些模型,涌现出大量的实证研究,已经成为发展心理学领域最为活跃的研究热点。
2 素质压力模型和双重风险模型
环境影响力的研究早期大多集中于逆境对个体的影响。大量证据表明, 那些成长于逆境(如贫穷、父母拒绝或不敏感、低质量的照料等)中的个体, 在认知、社会适应、情感和健康方面出现问题的风险也更高(Boyce, 2007; Luthar, 1999; McLoyd, 1998; Shonkoff et al., 2009)。在此基础上, 发展心理学领域的学者提出了相互作用/双重风险模型(Lazarus, 1993), 精神病学领域的学者则提出了素质压力模型(Monroe & Simons, 1991; Zuckerman, 1999)。
两个模型都强调个体所面临的、相互之间存在交互影响的“双重风险”:其一是个体自身所携带的固有风险或素质——可能是行为特征(如困难型气质、冲动性)、生理或内表型(如对压力的高生化物质反应)或先天基因(如5-HTTLPR的短等位基因)(Caspi et al., 2003);另外则是环境中的压力因素(如不良的照料)。但是,因为个体自身携带的风险因素不同,在环境压力下, 那些具有心理脆弱性和心理弹性的(Masten & Obradovic, 2006)个体会表现出截然不同的发展路径。
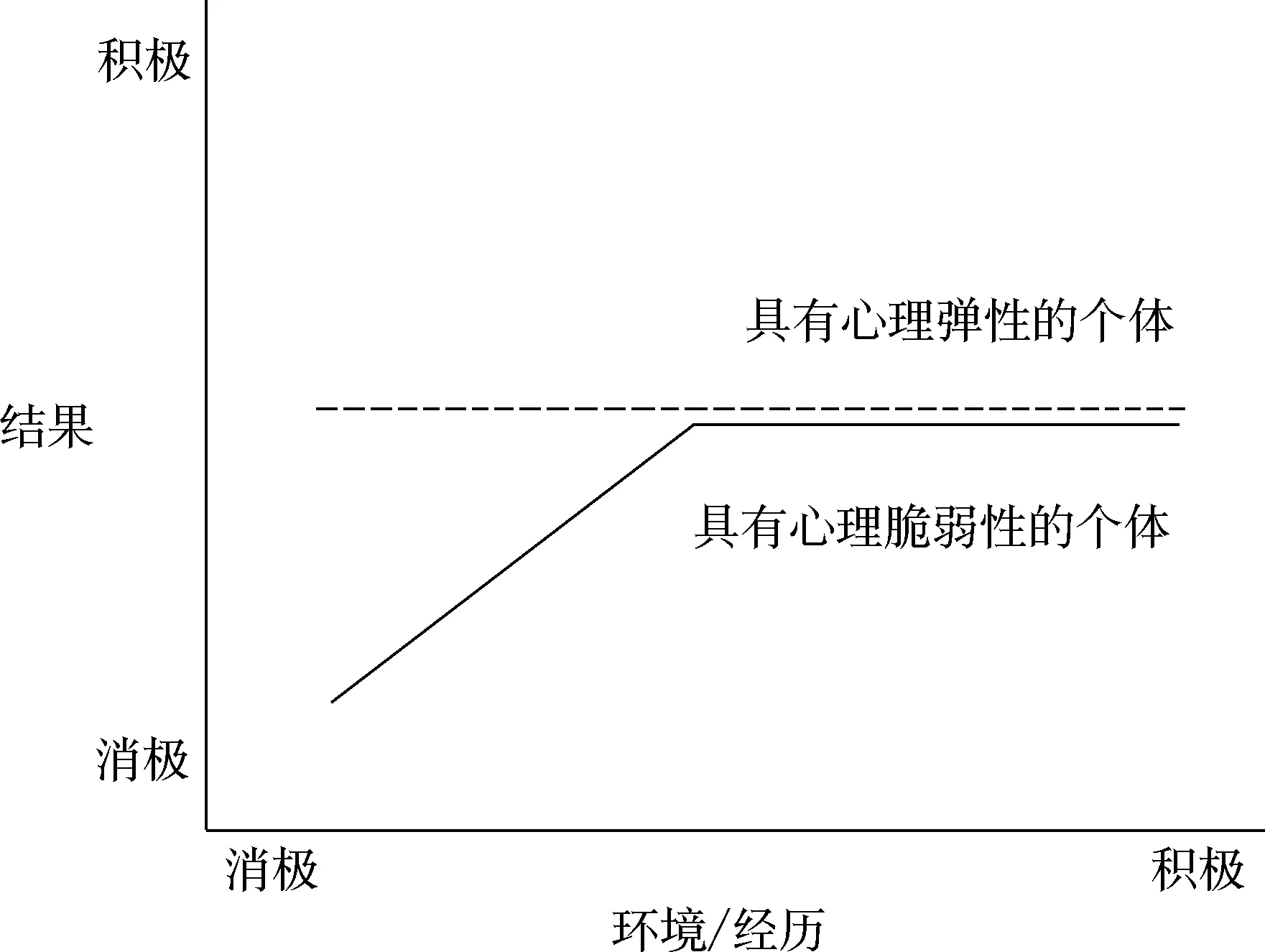
图1 素质压力模型/双重风险模型
如图1所示, 素质压力模型以及双重风险模型仅仅揭示了面对逆境时个体自身变量(心理脆弱还是富有弹性)同环境间的交互作用, 而没有涉及积极环境同个体自身特点间的相互影响。基于此局限性, 之后的研究者相继提出另外两个模型——环境的生物敏感性理论和差别易感性模型,从而更为全面地阐述环境同个体神经生物学特征之间的交互作用。
3 环境的生物敏感性理论
环境的生物敏感性理论(Boyce et al., 1995)指出, 儿童对环境影响的敏感性存在个体差异, 并且是以一种“更好和更坏”的方式同环境之间互动(这个“更坏”是从精神病理学和身体健康问题的角度来定义的, 不一定是更差的适应性结果)。
依据此理论, 高反应性儿童对积极和消极环境影响都表现出更强的敏感性, 被称为兰花型儿童, 他们对高压力和高支持的环境都特别敏感;相反, 反应性较弱的儿童被称为蒲公英型儿童, 他们对环境的感受较迟钝,在各种生存环境中基本都能完全适应;郁金香型儿童的反应性则介于两者之间,在情感反应和对环境的互动性(如冲动性、好奇心、社交状况等)上也处于中间水平(Lionetti et al., 2018)。
在此基础上,研究者(Boyce & Ellis, 2005)进一步设想, 早期环境的支持——压力水平和个体生物性反应特征的幅度之间存在一种U型关系(如图2),并提出以下假设:(1)幼年暴露在极度压力的环境中会增强个体对环境的生物敏感性, 提高其对环境中的危险或威胁的觉察和反应能力;(2)暴露在特别具有支持性的童年环境中也会提高个体对环境的生物敏感性, 提升其对社会资源和支持的敏感性;(3)相对而言, 大多数儿童成长于非极端的早期环境中, 从而降低了个体对环境的生物敏感性, 在一个既不是极度危险也不会一贯安全的世界里, 较低的生物敏感性会对环境中的慢性压力起到缓冲作用。之后的文献(Oshri et al., 2021; Shakiba et al., 2020)也进一步支持了环境的生物敏感性理论。
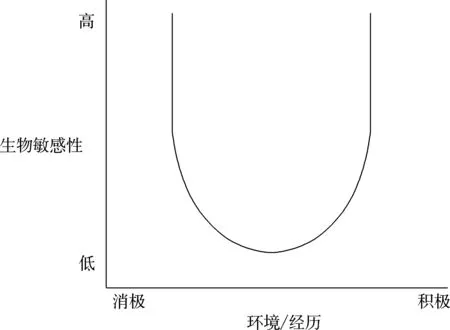
图2 环境的生物敏感性理论
4 环境的差别易感性理论
Belsky(1997)则在纯理论的基础上,提出了同环境的生物敏感性理论高度相似的理论——差别易感性理论。“为什么童年经历会影响个体之后的发展?”从进化角度来看, 只有当童年环境同后期环境有一定连续性或者前者会反复出现时, 利用童年经验来调整后期的心理和行为的个体才能获得可靠的收益, 即有条件的适应。然而, 未来在本质上是不确定的, 这就意味着有条件的适应在理论上具有局限性。
针对此问题, Belsky(2005)提出, 作为一种针对未来不确定性的风险对冲形式, 自然选择同时保留了以基因为基础的“条件性”和“替代性”的发展策略。所谓条件性发展策略, 是由环境因素所塑造的, 以使机体更好地适应未来环境。而替代性发展策略, 指行为取决于基因, 在很大程度上相对稳定、较少受环境影响(Rowe et al., 1997)。
之后,Belsky的合作者们(Ellis et al., 2011)进一步指出,对环境的易感性是在神经系统的生物反应中体现出来, 属于神经生物学的易感性,遗传易感性因素通过生物学过程起作用, 并且易感性的行为指标也是以神经生物学为基础。
从进化的角度来看, 神经生物易感性上的个体差异具有适应性。在不同的社会、物理和历史背景下, 不断波动的选择压力导致不同程度的适应度回报(至少在人类进化过程中是这样), 从而导致神经生物易感性的个体差异得以保留。
整体而言, 环境的差别易感性理论认为,对环境的神经生物易感性的不同,构成了人类发展中对可替代模式调控的中心机制。更确切地讲, 不同的易感性调节着环境对个体发展及生命轨迹的影响。从本质上看, 这就意味着即使经历和所处环境完全相同, 部分个体仍会比其他人更容易受到自身经历和所处环境的影响(Ellis et al., 2011)。
环境的差别易感性理论和环境的生物敏感性理论的相同之处在于, 两个模型都以人与环境的相互作用为核心, 认为在环境(充满压力或富有支持)对人类发展产生的影响中, 个体自身的神经生物学特征起到了调节作用。并且两者都假定在对环境影响的敏感性上存在个体差异。这两种理论都吸收并扩展了传统的素质压力模型的观点, 得出了非常关键性的结论:即那些最有可能受到消极环境不利影响(根据传统的心理健康标准)的人,也最有可能从支持性环境中受益。而且, 这些人受环境影响的表现模式是(遇到积极环境)变得更好(符合主流价值观所认同的如安全型依恋、幸福、高自尊、善于情绪调节、教育和职业成功、稳定的婚姻等)和(遇到消极环境)变得更糟糕(同样符合主流价值体系的如不安全型依恋、上瘾行为、社交问题、抑郁、学业失败、青少年怀孕等)。不过,目前在心理学领域的研究,通常围绕环境的差别易感性理论展开。
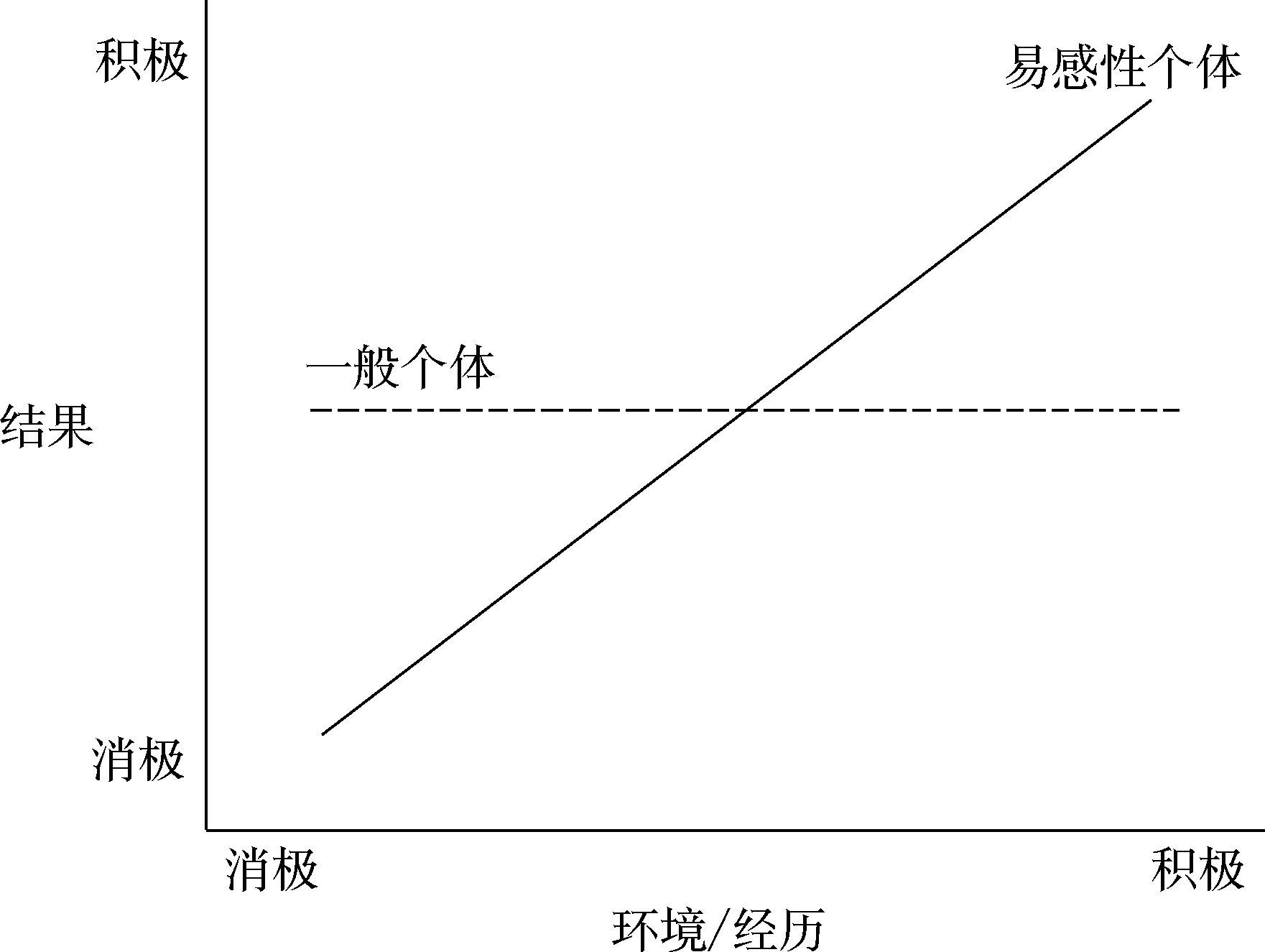
图3 差别易感性理论
5 优势敏感性假说
在差别易感性模型的基础上, 研究者(Pluess, 2017; Pluess & Belsky, 2013)又提出一个新概念——优势敏感性, 用来描述在极富支持性的环境下个体的不同反应, 这些反应也是源自个体自身固有特点的功能表现。优势敏感性概念重点强调个体对积极环境的反应性存在根本差异。Pluess和Belsky (2013)从以下角度详细描述了这一概念。首先, 优势敏感性反映了个体从环境中积极的、有利于促进个体幸福感和能力的因素中获益的一般倾向;其次, 优势敏感性的强弱表现为优势敏感因素(即促进性因素/promotive factors)所展现的功能;最后, 优势抵抗性描述的是个体未能从积极环境中获益, 类似于在素质压力模型中所描述的个体弹性对逆境的抵抗。进一步讲, 优势敏感因素增强了个体从积极因素中的获益程度, 而优势抵抗因素则降低或完全消除了积极因素的有利影响。目前文献(Iimura & Kibe, 2020; Rabinowitz & Drabick, 2017)进一步支持了优势敏感性假说。
优势敏感性假说虽然源自差别易感性模型, 但两者仍有明显不同。
首先, 优势敏感性假说认为, 鉴于有些人因为优势敏感因素的作用, 会对支持性或丰富性环境的好处更敏感, 但同样的个人属性可能不会使他们更容易受到逆境的消极影响。而差别易感性则意味着有些个体会对积极和消极的经历和环境都异常敏感。
优势敏感性理论认为个体的同一个内源性特征, 可能既对支持环境的反应性很强(即表现出增强的优势敏感性), 又对逆境的反应性不强并在逆境时起到保护作用(即表现出增强的弹性)。例如一个高智商的孩子, 可以从高质量的教育中更多获益, 但不管从理论还是经验上,人们都没有理由推断这样的学生也会从低质量教育中受到更多不利的影响。恰恰相反, 高智商儿童在面对逆境时往往更有弹性(Masten et al., 1999)。因此, 同样的个体特征(在这种情况下即高智商)可能在不利环境中起保护作用,但在支持性环境中却起促进作用,这正是优势敏感性假说所关注的现象。
其次,评价两者所需的实验条件不同。差别易感性的证据建立在一个更多样化的环境(范围从消极到积极)的基础上(Belsky & Pluess, 2009)。而优势敏感性则不同。事实上, 许多真正积极的环境暴露并不是从消极到积极的, 而只是从没有积极的到积极的, 如心理干预与没有心理干预。优势敏感性仅仅关注一个丰富或支持性体验所带来的积极益处。目前在心理干预领域,已经有大量文献支持了优势敏感性假说(闫玉鹏等,2016; Nocentini et al., 2018)。
最后, 差别易感性认为增加个体对逆境的脆弱性的因素也会增加个体对积极环境的优势敏感性, 而使个体对逆境具有弹性的因素也会使其对积极经验的反应性减少。而优势敏感性的概念与个体对积极情境条件的反应差异有关, 但这是促进因素的作用, 该假说并没有明确指出同一因素在逆境中会有潜在的保护作用。
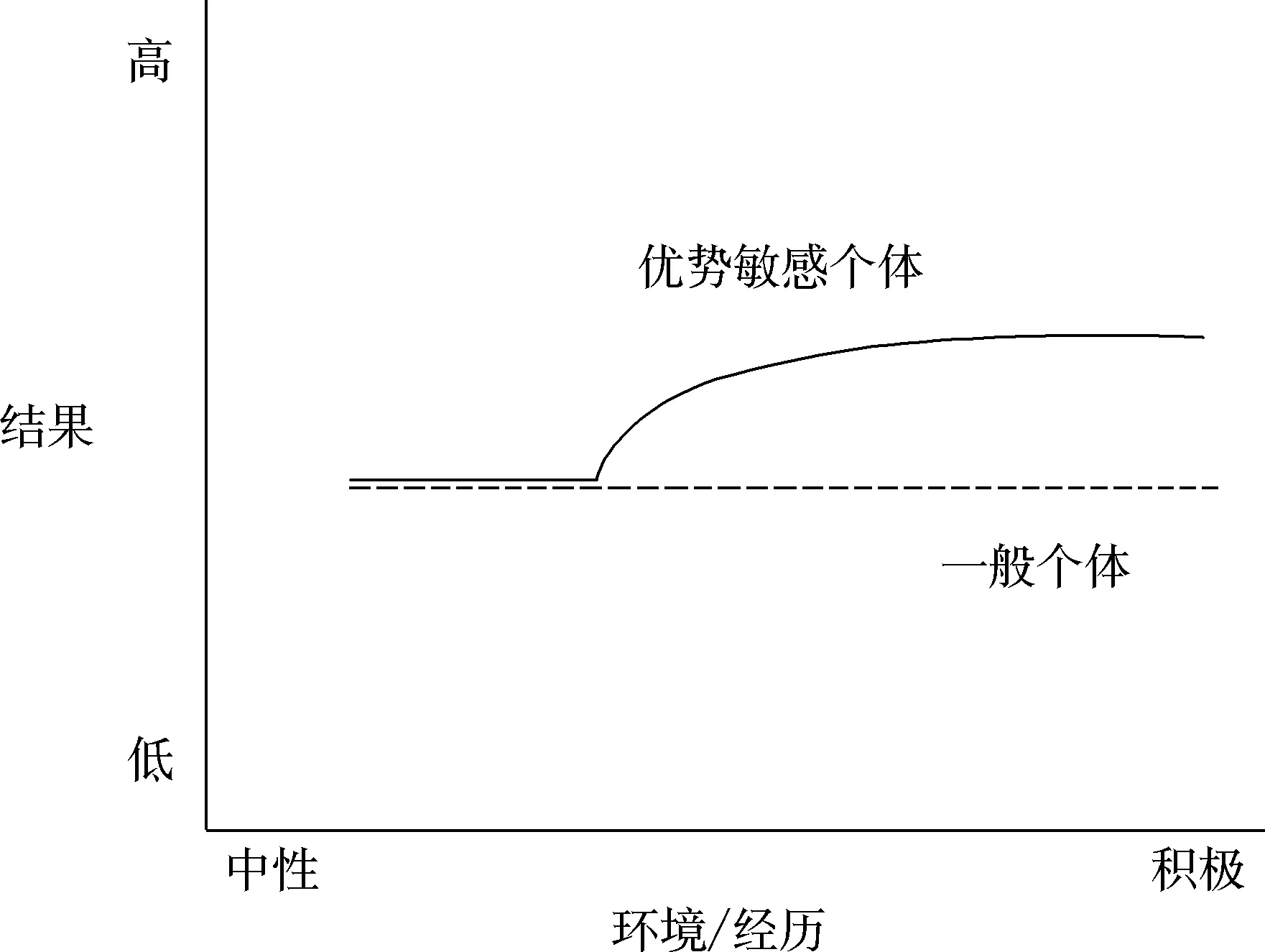
图4 优势敏感性模型
6 差别易感性理论的支持性证据
自环境的差别易感性理论和环境的生物敏感性理论提出以来, 研究者分别从表型特征、内表型机制和遗传多态性三个角度进行探讨, 得到了许多支持性的证据。
6.1 表型特征的研究证据
围绕表型特征的研究主要集中于个体的行为或表型特征, 鉴于这些个体的行为或表型特征对环境有调节作用, 因此可以作为可塑性(易感性)的潜在标记, 如困难型气质、冲动型和消极情绪。
大量文献支持了困难型气质在环境同儿童发展间的调节效应(Bradley & Corwyn, 2008; Pluess & Belsky, 2010)。一项追踪研究(Slagt et al., 2017)结果表明,同容易型气质的儿童相比,困难型儿童在消极养育模式之下会表现出更高水平的适应问题,但他们也从父母的积极养育模式中获益更多,进一步支持了环境的差异易感性理论。
在冲动行为上, Lengua等人(2000)对刚离婚的母亲和其9至12岁孩子的横断研究显示, 当母亲对他们要求的纪律是高度一致时, 高冲动的孩子表现出最少的抑郁症状, 但当母亲的纪律要求高度不一致时, 高冲动的孩子表现出最多的抑郁症状。此研究展现出一个以冲动性为中心的可塑性梯度:对于冲动性最强的那些孩子来说,教养方式和抑郁的关系最强, 对于冲动性弱的孩子来说, 两者关系最弱(并且不显著), 对于冲动性不太强也不太弱的孩子而言, 两者的关系强弱则介于中间。
此外, 对消极情绪的系列研究(高鑫等, 2018;Poehlmann et al., 2012;Slagt et al., 2017)表明,消极情绪高的孩子更容易受到教养方式和其他环境因素的影响, 这些儿童在父母教养质量较低时表现出更多的内化和外化行为问题, 社会和学业适应能力较差, 在父母教养质量较高时表现出较少的行为问题和较好的适应能力。
6.2 内表型机制的研究证据
内表型为介于基因和行为之间的个体属性, 表现为与基因成分和精神疾病临床症状相关的多种生物指标, 如血容量脉搏幅度、平均脉动压、皮肤导电水平等。来自内表型机制的大量文献为差异易感性理论进一步提供了有力证据。

此外, 对血容量脉搏幅度(Gannon et al., 1989)、 平均动脉压(Boyce et al., 1995)、儿童皮肤导电水平(El-Sheikh et al., 2007)、迷走神经(El-Sheikh et al., 2007)、呼吸性窦性心律不齐(Obradovic et al., 2010)、神经内分泌系统(Obradovic et al., 2010)及自主神经反应(Gannon et al.,1989)等的研究均有类似发现, 这些结果都支持了环境易感性理论。
6.3 遗传多态性的研究证据
进入21世纪以来, 基因领域的大量研究为差异易感性理论提供了更为有力的证据。现有文献主要集中于以下几种类型的基因。
一是围绕神经递质代谢酶单胺氧化酶A基因(MAOA)展开的系列研究。MAOA具有降解去甲肾上腺素、多巴胺以及五羟色胺等单胺类神经递质的作用,同反社会行为之间存在重要联系。中外大量研究(刘立敏等,2017;刘少将等,2017;Kim-Cohen et al., 2006; Mariz et al., 2022; Zhao et al., 2020;)支持了MAOA同环境在个体的反社会行为上存在着交互作用。
例如,张文新等人的研究(Zhang et al., 2016)指出, 具有MAOA基因T等位基因/TT纯合子的青少年在接受低程度积极养育时表现出更强的反应性攻击, 而在接受高程度积极养育时表现出更弱的反应性攻击。曹丛等人(2016)的研究则发现MAOA基因 rs6323多态性与母亲支持性教养行为对青少年抑郁具有交互作用, 母亲支持性教养行为可以显著负向预测 GG 基因型女青少年的抑郁水平, 但对 TT基因型女青少年抑郁的预测作用并不显著。另一项对MAOA 基因 rs6323 多态性与同伴关系对男青少年早期抑郁影响的研究(王美萍等, 2015)指出, 当同伴接纳水平较低时, G等位基因携带者的抑郁水平表现出高于T等位基因携带者的趋势, 当同伴接纳水平较高时, G基因携带者的抑郁水平显著低于T等位基因携带者, 研究结果进一步指出, 同伴接纳可以显著预测 G 等位基因携带者的抑郁, 但对 T 等位基因携带者的抑郁无显著预测作用。
二是围绕羟色胺相关基因的研究。针对羟色胺系统中血清素转运蛋白连接的多态性区域(5-HTTLPR)的相关研究主要集中在携带至少一个短等位基因(s/s, s/l)和长等位基因纯合子(l/l)的三个变异型上, 尽管5-HTTLPR已经鉴定出更多的变异型(Nakamura et al., 2000)。
尽管先前的大量研究认为(Brody et al., 2009; Caspi et al., 2003; Karg et al., 2011; Manuck et al., 2004),5-HTTLPR是抑郁症的风险因素,能与压力性生活事件相互作用,越来越多的证据表明,5-HTTLPR对抑郁症的影响十分有限(Border et al., 2019; Culverhouse et al., 2018)。尽管5-羟色胺能神经传递在HPA 轴调节中的重要作用(Porter et al., 2004),抑郁症状的发展依然无法由5-HTTLPR单个基因的影响进行解释(Kuhn et al., 2021)。
此外,早期生活压力可以通过包括 DNA甲基化和mRNA表达在内的表观遗传机制影响基因表达(Lopizzo et al., 2015)。研究表明,早期环境对HTR2A基因的甲基化及婴儿和儿童的行为结果相关(Eszlari et al., 2019; Paquette et al., 2013)。在对羟色胺系统中血清素受体基因HTR2A上, 芬兰早期展开的大型调查发现,携带一个或多个HTR2A T102C多态性的T等位基因的成年个体, 如果是在低社会经济地位的家庭长大, 那么在伤害回避气质评估上得分最高,即最具有伤害回避气质特质, 但如果在社会经济地位高的家庭中长大, 那么在伤害回避上得分最低(Jokela et al., 2007)。近期研究(Parade et al., 2017)发现,早期环境压力暴露与HTR2A-1420位点的甲基化相关,创伤后应激障碍和重度抑郁症症状与HTR2A-1420的甲基化呈负相关,但与HTR2A-1224的甲基化呈正相关。
三是对儿茶酚氧位甲基转移酶(COMT)Val158Met基因多态性的研究。这是一种常见的功能多态性,与不良行为和攻击性有关。
Caspi等人早期研究(2005)发现,COMT基因的功能多态性调节了青春期吸食大麻对成年期精神病发展的影响,那些携带COMT valine158等位基因的人如果使用大麻,最有可能表现出精神病症状,并发展成精神分裂症,但是,对于那些携带两个蛋氨酸等位基因的个体,使用大麻并没有起到这样的不利影响。
在近年文献中,Hygen等人(2015)观察到童年严重生活事件与COMT基因型的显著交互作用:与携带Met的配对儿童相比,面临许多严重生活事件且为Val纯合子的儿童表现出更多的侵略性(p=0.02)。值得注意的是,在没有严重生活事件的情况下,Val纯合子的攻击性得分显著低于Met携带者(p=0.03)。Val纯合子更容易受到严重生活事件对攻击性的影响。与此类似,曹衍淼等人(2017)对COMT 基因Val158Met 多态性的研究表明,在低同伴接纳或高同伴拒绝环境中, 携带ValVal基因型的青少年抑郁水平高于Met等位基因携带者;在高同伴接纳或低同伴拒绝环境中, 携带 ValVal基因型的青少年抑郁水平低于Met等位基因携带者。但是,与Hygen等人(2015)的结果不同,国内学者张文新等人(2016)发现,携带COMT基因Met等位基因的青少年在接受低程度积极养育时表现出更强的反应性攻击, 而在接受高程度积极养育时表现出更弱的反应性攻击。越来越多的研究证明(林小楠等, 2018),父母养育行为对青少年早期抑郁的影响与COMT基因密切相关。
最后,国内外学者还围绕多巴胺受体进行了相关探索。例如,国内学者张文新等人(2015)发现多巴胺D2受体多态性(DRD2 TaqIA)与母亲教养方式对青少年早期抑郁症状具有交互作用。最新的研究(李曦等, 2020)表明,DRD4基因还与积极的环境互动和亲社会行为存在密切关联。Van Ijzendoorn和Bakermans-Kranenburg(2006)对多巴胺受体D4(DRD4)多态性的研究发现,携带这种假定的风险等位基因的个体不仅更容易受到较差的环境条件的不利影响,而且比其他人更能从高质量的环境条件中获益。另一项研究(Sheese et al., 2007)指出,携带7-重复DRD4等位基因的幼儿与不携带此基因的幼儿相比,当教养行为质量较高时,幼儿寻求刺激的行为较少,而当教养行为质量较低时,幼儿寻求刺激的行为较多。其后的干预实验(Bakermans-Kranenburg & Juffer, 2008; Van Ijzendoorn et al., 2008)也有类似的发现。
除了上述四个领域之外, 研究者对儿童催产素受体基因(OXTR)rs53576(Hygen et al., 2017)、脑源性神经营养因子(BDNF)Val66Met基因多态性(张璠等, 2015)进行的研究均支持了Belsky的“差别易感性理论”。
7 差别易感性理论面临的挑战
虽然近十多年间的大量文献从不同角度支持了差别易感性假说,但目前为止,仍然有证据对此理论提出挑战,结果依然支持素质-压力模型。
近期针对135名36个月大婴儿同母亲间的互动研究(Vrantsidis et al., 2022)发现,母亲的消极回应水平越高,那些在多基因复合型(COMT, DAT1, DRD2, DRD4)得分上更高的孩子在自控方面表现越差,结果并不支持这些孩子在积极母亲养育下的更强自控力。此外,国内刘亚鹏等人(2019)对5-羟色胺转运体基因连锁多态性(5-HTTLPR)的研究发现,在母亲面临较大养育压力的家庭中,携带s/s和s/l型基因的男孩的攻击和反抗行为明显高于l/l型男孩,但是,在母亲养育压力水平较低的家庭中,两组男孩的行为表现并没有明显不同。另一方面,该研究发现当母亲承受的养育压力较大时,携带l/l型基因女孩的抑郁水平明显高于s/s和s/l型女孩,但在母亲养育压力水平较低的家庭中,两组女孩的抑郁水平差异不明显。这些研究的结论均支持了素质压力模型,而非差别易感性假设。
而张芸等人(2015)围绕单胺氧化酶A串联重复序列多态性研究显示,MAOA-VNTR基因型同童年期虐待并没有对女性青少年的冲动表现产生明显的交互作用。另外,中国学者近期围绕羟色胺系统中5-HTR1A 基因rs6295多态性的研究(王美萍等, 2015)指出,当父母表现出较高水平的积极教养行为时, 携带CC基因型的女青少年抑郁水平显著低于G等位基因携带者,但是,当父母积极教养行为水平较低时,携带不同基因型的女青少年的抑郁水平差异不显著。此外,该研究还指出,rs6295多态性与父母教养行为对男青少年的抑郁不存在交互作用。由此可见,此研究结果只是在女青少年中部分支持了差别易感假设。
另外一项针对幼儿园儿童气质和差别易感性的研究(MeikeSlagt et al., 2019)指出,幼儿在父母养育过程中的情绪反应与各项易感性特质均无关,对四个模型的假设均不支持。在一项日本高中生的纵向研究(Shuhei & Chieko, 2020)中,研究者同时检测了素质压力模型、差别易感性理论和优势敏感性模型,尽管在比较之下确实有拟合指数之间的差异,但仍没有一种模型能完全符合实验数据结果。面对争议,环境易感性理论依然缺乏有力的研究证据,因而该领域依然存在极大的研究空间。
8 结语
自提出至今的20多年时间里, 环境的差别易感性模型已经得到越来越多的研究证据支持。围绕个体神经生物学特征同环境对个体发展共同影响的研究目前仍是个相对活跃的领域, 国内外学者近年也围绕该领域进行了大量探讨,该领域还有很多问题亟待解决。
但是,毋庸置疑的是,在教育内卷化的当今社会,差别易感性理论为广大教育工作者及家长提供了崭新的教育视角,尤其是那些在内卷大浪中所谓的适应不良儿童,他们很有可能是蕴藏着巨大发展能量的“兰花型”儿童,当家长和学校提供适合的环境时,他们更有可能绽放出惊人的光彩。在教育实践中,当教育者以分类的视角重新审视发展中的儿童,从“兰花”“郁金香”和“蒲公英”的形象中捕获不同孩子的发展优势时,那么,“因材施教”的理念便可以在更大范围之内更为有效地落到实处。
