GhEIN3 基因对棉花枯萎病胁迫响应的功能分析
2022-08-08赵曾强张析李潇玲张薇
赵曾强,张析,李潇玲,张薇*
(1.石河子大学农学院,新疆 石河子 832000;2. 新疆农垦科学院生物技术研究所,新疆 石河子 832000)
乙烯(ethylene, ET)不敏感蛋白3(ethyleneinsensitive 3,EIN3)和EIN3-like(EIL)蛋白是乙烯信号转导途径中重要的核转录因子,其C 端保守性较低,N 端高度保守,包含酸性氨基酸区、脯氨酸富集区、碱性氨基酸区等,含有DNA 结合的重要结构域,能与初级乙烯响应元件(primary ethylene reaction element, PERE)结合调节相关基因表达,参与生物与非生物胁迫响应[1-3]。 目前关于该类转录因子基因的研究主要在拟南芥(Arabidopsis thaliana)[4](6 个成员)、 烟草(Nicotiana tabacum)[5-6](5 个成员)、 番茄(Lycopersicon esculentum)[7-8](4 个成员)、水稻(Oryza sativa)[9-10](6 个成员)等植物中,通过拟南芥建立的乙烯响应的线性信号转导模型研究表明,EIN3/EIL家族基因是乙烯信号通路中的关键基因[11-12]。
在拟南芥中,乙烯受体蛋白(ethylene receptor 1, ETR1; ethylene receptor 2, ETR2)、乙烯响应传感蛋白(ethylene response sensor 1, ERS1; ethylene response sensor 2, ERS2)、 乙烯不敏感蛋白4(ethylene-insensitive 4, EIN4) 等是膜相关受体蛋白的主要组成成分,能够感知乙烯信号并负调控该途径,膜相关受体的激活态或失活态与乙烯的结合有关[13-15]。 研究表明,无乙烯存在时,膜相关受体的激活态与负调控因子组成型三重响应蛋白1(constitutive triple response 1, CTR1)结合,通过促分裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)途径将信号传递至EIN2,从而抑制由EIN2 和EIN3 介导的信号通路应答[16-17]。 乙烯存在时,膜相关受体(如ETR1/2 等)失活,不能与CTR1 结合,CTR1 抑制下游乙烯响应开启,从而使EIN3 蛋白在细胞核快速积累,参与调控或激活下游其他转录因子基因的表达[16, 18-20]。
EIN3/EIL家族基因是否参与植物抗逆响应?据研究报道,拟南芥ein3-1和eil1-1双突变体对高浓度盐的耐受性显著降低[21]。 陈华民[22]发现EIN3 通过负调控水杨酸(salicylic acid, SA)信号通路调控植物的先天免疫反应; 在大豆(Glycine max)中过表达EIN3-1、EIN3-2能明显增强植株对大豆疫霉根腐病的抗性[23];从矮牵牛(Petunia hybrida)中分离得到的3 个转录因子PhEIL1~3中,只有PhEIL1受乙烯的诱导[24]。 研究表明,病程相关(pathogenesis-related, PR)蛋白在植物抗病反应中具有重要意义[25]。乙烯是重要的信号分 子,EIN2、EIN3、EIN5/AIN1、EIN6、EIN7 是 乙烯响应的正调控因子,负责乙烯信号转导;而定位于核内的DNA 结合蛋白EIN3,直接作用于乙烯响应因子1(Ethylene response factor 1, ERF1),调节乙烯响应基因的转录, 激活植物防御素和PR基因表达,激活植物的抗病性反应[26]。 棉花(Gossypium)基因组分析揭示,乙烯信号转导相关的基因涉及2 个途径[27],EIN3 和EIL 是相关的转录因子。韩利红[28]将GhEIN3基因导入拟南芥,并获得过表达该基因的拟南芥, 黑暗条件下用1- 氨基环丙烷-1- 羧酸(1-aminocyclopropane-1-carboxylic acid, ACC)对其进行处理,结果显示,过表达GhEIN3的拟南芥植株的下胚轴短于野生型和双突变体ein3eil1,表明GhEIN3在拟南芥中能够响应乙烯信号;Wang 等[29]发现GhEIN3参与棉花耐盐性调控。
棉花枯萎病是严重影响海岛棉(G.barbadense)品质及产量的土传真菌病害之一,而关于棉花中是否存在响应枯萎病菌胁迫表达的EIN3/EIL 转录因子基因的研究报道甚少。病毒诱导的基因沉默(virus-induced gene silencing, VIGS)技术不需遗传转化,能够在短时间内实现基因沉默,被开发为快速、高效、高通量的反向遗传学技术, 在棉花基因功能研究中得到广泛应用[30]。
VIGS 技术利用注射等方式接种棉花子叶, 操作简单,不受基因型限制,沉默效率高,约2 周便可实现靶基因沉默,可大幅缩短研究周期[31-33]。 因此,本研究通过分析枯萎病菌诱导棉花根部基因表达谱数据获得EIN3/EIL 转录因子基因GhEIN3,利用生物信息学工具、实时荧光定量聚合酶链式反应 (quantitative real time polymerase chain reaction, qRT-PCR)技术和VIGS 技术初步分析该基因参与调控棉花对枯萎病抗性的途径,以期为进一步研究其功能提供重要依据。
1 材料与方法
1.1 试验材料
棉花材料:抗枯萎病陆地棉(G.hirsutum)品种中棉所12;枯萎病菌:强致病力菌株7 号生理小种F430[34];VIGS 载体pTRV1 与pTRV2 载体由石河子大学农学院刘慧英老师惠赠;pTRV2∷GhCLA1 重组载体由本实验室保存。
1.2 材料种植
选籽粒饱满的种子,经10%(体积分数)过氧化氢浸泡3 h,无菌水冲洗5~6 次后种植于无菌蛭石中,待棉苗根部生长至3 cm 左右,移入霍格兰氏(Hogland's)溶液,置于25 ℃、光照16 h/ 黑暗8 h 的培养箱中继续培养,并每周更换溶液[35]。
1.3 材料处理
枯萎病接菌处理:种植棉苗400 余株,在生长过程中去除弱苗与长势不一致的棉苗,待棉苗第1 片真叶完全展开,选取长势一致的幼苗伤根后浸泡于孢子含量为1×107mL-1的枯萎病菌孢子悬浮液中40 min, 而后转入霍格兰氏溶液培养[36]。分别取未处理(CK)和处理后1 h、2 h、3 h、6 h、12 h、24 h、48 h 的棉苗完整根部组织,同时设置平行对照(Mock),即相同生长环境下相同取样时间的清水处理棉苗根部组织,共处理棉苗150 余株,样品经液氮冷冻后保存于-80 ℃冰箱备用[3]。
激素处理: 待棉苗第1 片真叶完全展开,选取长势一致的幼苗分别浸泡于含有1×10-3mol·L-1乙烯、5×10-5mol·L-1SA、1×10-3mol·L-1茉莉酸(jasmonic acid,JA)和不添加任何激素的清水(Mock)中40 min,然后转移至新鲜霍格兰氏溶液中,分别取未处理(CK)和处理后1 h、2 h、3 h、6 h、12 h、24 h、48 h 的棉苗完整根部组织,共处理棉苗200 余株, 样品经液氮冷冻后保存于-80 ℃冰箱备用[3]。
1.4 GhEIN3 基因克隆及表达特征分析
1.4.1GhEIN3 基因克隆及序列分析。 棉花根部总RNA 的提取采用改良的CTAB 方法[37],cDNA第1 链的合成参考TaKaRa 反转录试剂盒说明书操作。GhEIN3基因扩增程序:94 ℃预变性5 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸4 min,32 个循环;72 ℃延伸10 min。 将阳性菌株送至生工生物工程(上海) 股份有限公司进行基因测序。 扩增引物为GhEIN3-F:GCTCTAGAATGATGATGTTTGACGAGATGGG,GhEIN3-R:CGAGCTCCTGGAACCAGATTGAAACATCC。
利用NCBI(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)ORF finder 分 析 开 放 阅 读 框(open reading frame, ORF);利用SMART(http://smart.embl-heidelberg.de/smart/set_mode.cgi?GENOMIC=1) 进行蛋白保守结构域分析; 利用GOR4(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html)进行蛋白二级结构分析;利用SWISS-MODEL(http://swissmodel.expasy.org/)进行蛋白结构域三级结构的同源建模分析;利用ExPaSy ProtScale(http://web.expasy.org/cgi-bin/protscale/protscale.pl) 进行蛋白亲/疏水性分析;利用ExPaSy ProtParam(http://web.expasy.org/protparam/)进行蛋白理化性质分析;利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) 对GhEIN3基 因 上 游2 000 bp序列进行启动子分析; 利用NCBI 中的Blast 程序(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM = blastp & PAGE _ TYPE = Blast-Search &LINK_LOC=blasthome) 进行同源性比对,数据库选择非冗余蛋白质数据库(Nonredundant protein sequence, NR), 参数为程序默认设置,获得的同源序列登录号见表1。获得的序列利用Mega 7.0 软件进行系统进化分析, 利用DNAMAN 6.0 软件进行同源性比较。

表1 GhEIN3 基因同源序列Table 1 Homologous sequences of GhEIN3 gene
1.4.2GhEIN3 基因表达特征分析。 选取在棉花根和叶中表达稳定的GhUBQ7(DQ116441)作为内参基因[38],qRT-PCR 反应体系及程序参见TaKaRa 公司(大连)TB GreenPremix EX TaqTM说明书;设3 个重复,采用2-ΔΔCt 法计算基因相对表达量[39]。 所用引物为qGhEIN3-F:GCCCTTACAGTGAATTTCGTT,qGhEIN3-R:TACTGACATGAGTTCGCCGAT;GhUBQ7-F:GAAGACCTACACCAAGCCCAA,GhUBQ7-R:CGGACT CTACTC AATCCCCACC。
1.5 VIGS 载体的构建
根据GhEIN3基因序列设计特异性引物,利用PCR 扩增目标片段, 采用酶切连接法进行VIGS 载体构建, 将构建好的pTRV2∷GhEIN3重组载体通过电击法转入根癌农杆菌GV3101,对菌液采用PCR 技术筛选鉴定阳性菌株。 所用引物为GhEIN3-V-XbaI-F:GCTCTAGATGATCCTCCTCAAAGGCGAT,GhEIN3-V-KpnI-R:GGGGTACCCAAAG TTCGGCTCATCTTCAAC,下划线示限制性内切酶识别位点。
1.6 农杆菌介导的VIGS 基因沉默与沉默植株抗病观察及相关基因表达的检测
VIGS 试验操作参考棉花病毒诱导的基因沉默体系及优化方法[40]。GhEIN3沉默植株枯萎病抗病性鉴定:以注射pTRV2∷GhCLA1 的棉花叶片出现报告基因GhCLA1导致的白化表型后,对基因沉默的棉苗(记为TRV-GhEIN3)和对照棉苗,进行伤根处理,并对每株棉苗接种孢子数为(注射空载体pTRV2∷00 的棉花,记为mock)1×107mL-1的枯萎病菌孢子悬浮液10 mL,然后在温度25 ℃/23 ℃(光照/黑暗)、光照16 h/黑暗8 h,相对湿度75%条件下培养,设3 个重复,每个重复15 株,定期观察发病情况。
沉默株系中抗病相关基因表达检测方法:沉默植株接菌及培养条件同抗病性鉴定。 所检测的抗病相关基因包括GhERF1(XM_016876293.1)、GhACO( NM_001326965 . 1 ) 、GhPR1( XM_016882081.1)、GhPR2(XM_016813433.1)、Gh PR4(XM_016893098.1)、GhPR5(XM_016893098.1)。试验选取在棉花根和叶中表达稳定的GhUBQ7(DQ116441)设计内参引物[38],qTR-PCR 反应体系及程序参见TaKaRa 公司(大连)TB GreenPremix EX TaqTM说明书,设3 个重复,采用2-ΔΔCt法计算基因相对表达量[39],利用Olig 7 软件设计上述基因引物,其序列见表2。

表2 引物及其序列Table 2 Primer and their sequences
2 结果与分析
2.1 GhEIN3 基因克隆与序列分析
利用枯萎病菌诱导棉花根部基因表达谱和棉花数据库从陆地棉中筛选获得1 个ORF 完整且编码氨基酸序列含有典型EIN3 蛋白结构域的基因(图1A、B);以棉花根部组织cDNA 为模板,通过PCR 扩增及测序分析, 基因ORF 全长为1 842 bp(Genbank 登录号:KY744279,图1A),编码的蛋白相对分子质量为69 609.64,等电点为5.41,为亲水不稳定蛋白,该蛋白二级结构分析表明,无规则卷曲的比例较高,为55.79%,是其二级结构的主要组成元件(图1C);启动子序列分析表明,除启动核心元件和光响应元件外,还包括干旱响应元件,SA、脱落酸及赤霉素响应元件,低温响应元件,MYB 结合位点和DNA 结合蛋白位点(图1D)。 系统进化树分析表明,GhEIN3与澳洲棉KAA3468812.1亲缘关系较近(图2);同源比对分析表明,GhEIN3编码的氨基酸序列与同源基因编码的氨基酸序列的保守结构域主要集中在N 端(图3)。

图1 GhEIN3 基因克隆及序列分析Fig. 1 Clone and sequence analysis of GhEIN3 gene
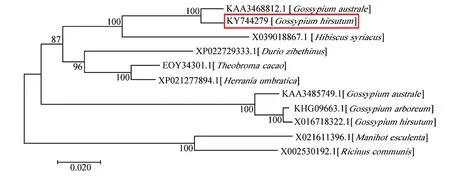
图2 GhEIN3 基因与其他植物EIN3/EIL 基因编码的氨基酸序列系统进化树Fig. 2 Phylogenetic tree of amino acid sequences encoded by GhEIN3 and EIN3/EIL genes in other plants

图3 GhEIN3 基因与其他EIN3 基因编码的氨基酸序列的同源比对结果Fig. 3 Alignment of deduced amino acid sequences of GhEIN3 and the other related EIN3 genes
2.2 GhEIN3 基因的诱导表达分析
枯萎病菌侵染后,GhEIN3在各处理时间点的相对表达量均高于Mock(图4A);其中处理后24 h, 该基因的相对表达量达到最大值, 是Mock 的3.72 倍(差异显著),显示该基因能被枯萎病菌诱导上调表达,推测该基因参与棉花枯萎病胁迫响应。
为了进一步检测GhEIN3基因是否对激素信号具有应答响应, 对抗病品种中棉所12 的棉苗分别用3 种激素(SA、JA 和ET)处理。 基因表达量检测结果显示:ET 处理后,该基因相对表达量呈现先上升后下降的变化趋势, 在处理后3 h达到峰值,且为同时期Mock 的5.43 倍,差异极显著(图4B);SA 与JA 处理后,该基因的表达模式相似,SA 处理后6 h 开始其表达量均低于Mock,JA 处理后其表达量均低于Mock, 在SA处理后48 h 与JA 处理后24 h 时,GhEIN3的表达量最低, 分别为Mock 的5.54%和2.07%(图4C、D)。 据此推测,ET 对GhEIN3基因的表达为正调控,SA 与JA 对GhEIN3基因的表达为负调控。

图4 不同胁迫处理后GhEIN3 基因的表达模式Fig. 4 Expression pattern of Ghein3 gene under different stresses
2.3 GhEIN3 基因VIGS 载体构建
以棉花根部组织的cDNA 为模板,利用特异性引物进行PCR 扩增,片段为400 bp(图5A),回收产物连接T 载体并进行双酶切验证(图5B)并进行测序,选择正确的重组质粒,采用酶切连接的方法进行VIGS 载体构建,对重组载体pTRV2∷GhEIN3 进行双酶切验证(图5C),结果均与预期相一致,表明成功构建了该基因沉默重组载体pTRV2 ∷GhEIN3。 然后用其转化农杆菌,于-80 ℃保存备用。

图5 pTRV2∷GhEIN3 载体的琼脂糖凝胶电泳结果Fig. 5 Agarose gel electrophoresis of pTRV2∷GhEIN3 vector
2.4 GhEIN3 基因沉默效率及沉默植株抗病性检测
注射后10 d,pTRV2∷GhCLA1 处理植株真叶出现白化现象(图6A),表明VIGS 体系可以成功抑制报告基因GhCLA1表达,且该体系能够用于棉花中基因功能研究。 在棉苗注射pTRV2∷GhEIN3 后10 d,GhEIN3基因表达水平检测结果表明,GhEIN3基因在沉默植株中的表达量为注射空载体植株的16.42%,由此可知,沉默效率约84%(图6B)。 伤根法接菌后14 d,沉默植株下部叶片快速萎蔫变黄然后变枯,与对照相比,沉默植株较感病(图6C~F)。

图6 GhEIN3 基因沉默效率及沉默植株抗病检测结果Fig. 6 Silencing efficiency of GhEIN3 gene and detection of disease resistance in silenced plants
2.5 沉默株系中抗病相关基因的表达分析
PR1、PR2、PR4、PR5基因表达分析结果 (图7)表明,在沉默株系中PR1、PR2、PR4的相对表达量均低于空载体对照,分别低50%、28%、30%,而PR5的相对表达量高于空载体对照46%;乙烯合成及信号通路相关基因ACO和ERF1在沉默植株中的相对表达量均显著低于空载体对照。 由以上结果推测,GhEIN3基因通过调控病程相关基因与乙烯信号通路关键基因的表达调控棉花对枯萎病的抗性。
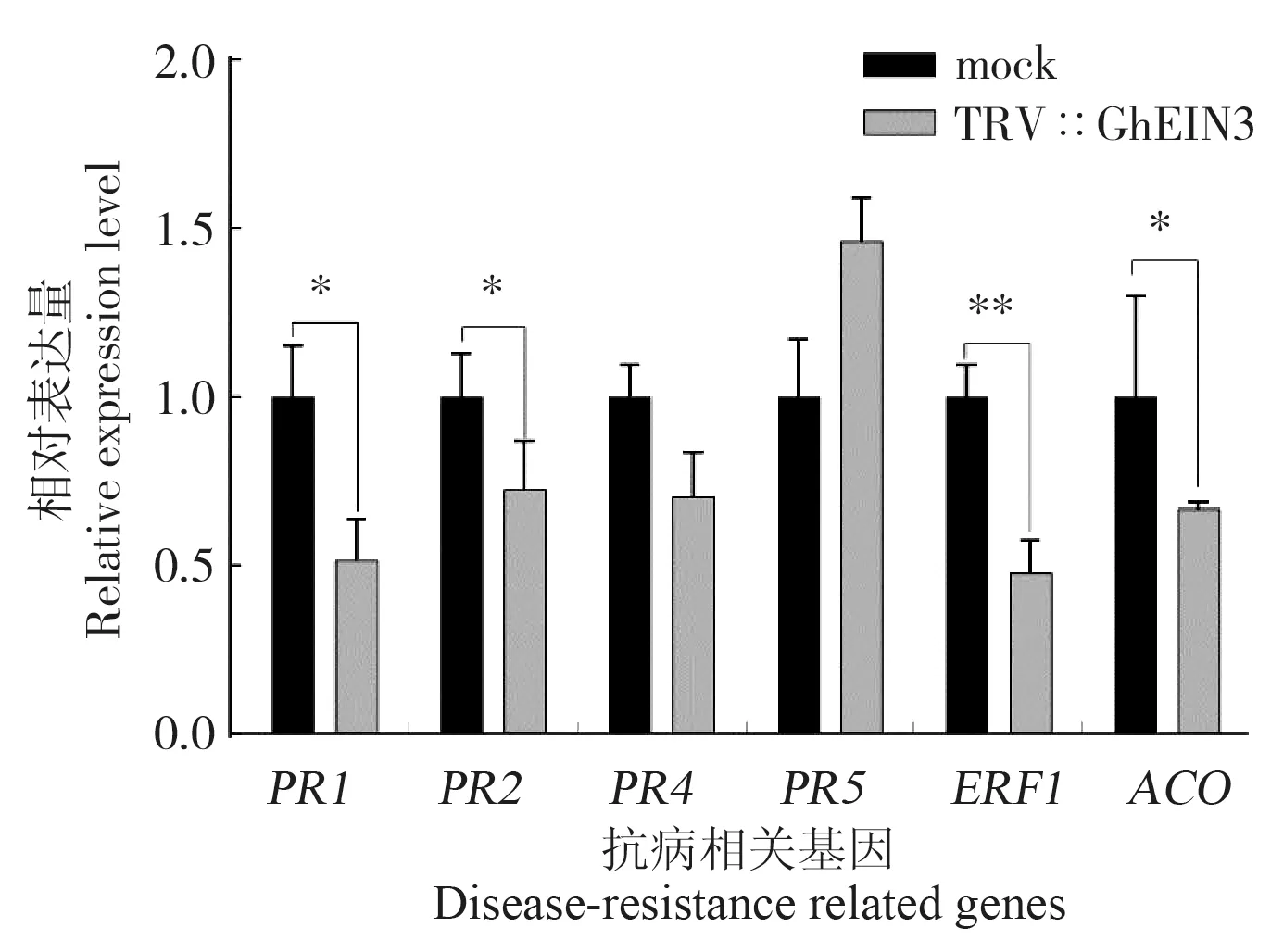
图7 GhEIN3 基因沉默植株接种F430 后抗病相关基因的表达分析Fig. 7 Expression analysis of disease resistance related genes in GhEIN3-silenced plants infected with F430
3 讨论
乙烯在植物生长发育及逆境应答中发挥重要的作用, 而EIN3 和EIL 是乙烯信号通路的关键转录因子,EIN3 能够特异性结合乙烯响应相关基因启动子的特定DNA 序列, 进而激活这些基因的转录,参与植物抗逆响应[41]。
本研究根据枯萎病菌诱导棉花基因表达谱在棉花基因组中鉴定出响应枯萎病菌的转录因子GhEIN3基因,ORF 全长为1 842 bp,编码613个氨基酸, 所编码氨基酸中含有典型EIN3 结构域,枯萎病菌侵染后,基因表达量呈上调趋势,推测其参与了棉花抗枯萎病响应。 前人研究表明,过表达EIN3-1、EIN3-2的大豆植株对大豆疫霉根腐病的抗性明显增强[23];油菜(Brassica napus)中BnEIN3基因能被核盘菌诱导上调表达, 且其在高抗病品种中的表达水平显著高于中抗病品种和感病品种[42]。 以上研究结果与本研究结果一致。
研究表明,SA、JA 和ET 是植物抗逆信号转导中的重要信号分子[43-44]。 研究使用外源激素SA、JA 和ET 分别处理棉花幼苗,利用qRT-PCR技术分析GhEIN3基因表达特征, 结果表明,施加ET 后,GhEIN3基因在各处理时间点的表达量均高于同时期Mock,说明其为ET 通路中的正调控因子, 也与前人对该基因家族研究结果一致;但施加SA、JA 后, 该基因表达量整体低于Mock,表明SA、JA 抑制GhEIN3基因表达。 研究显示,SA 与ET 之间存在拮抗关系[45],而EIN3 在病原微生物胁迫下, 同时能负调控SA 生物合成途径关键基因SID2的表达[22],也能够抑制JA 途径诱导的烟碱合成酶基因表达[46],推测该基因可能是SA 与ET 拮抗作用的节点。 大部分研究认为,JA 与ET 两个信号通路协同作用, 共同调节下游基因的表达; 但也有研究表明,ET 与JA 之间也存在拮抗作用,ET 能够抑制JA 途径中THI2.1、VSP等关键基因的表达[47],暗示各个激素信号转导途径所构成的信号传递网络中存在复杂的调节作用,而并不是单一的拮抗或者协同作用。GhEIN3基因在SA、JA 和ET 信号通路交互过程中所起到的作用也有待进一步研究。
研究发现,SA 通路为SA→NPR1→TGA→PR1→抗病,NPR1基因是多种信号途径的交叉点,通过与TGA 转录因子的相互作用调控PR基因 表 达[48];EIN3和EIN3-Like1 基 因 通 过 抑 制SID2编码的ICS1 表达而负调控病原菌诱导的SA 合成[49]。白桦中,抑制BpEIN3表达后,转基因植株中参与脱落酸信号转导的相关基因、SA 信号转导相关的NPR1基因和JA 信号通路的负调控基因JAZ10均上调表达[50],而NPR1 最早被发现在拟南芥中正向调控SA 信号通路中PR基因的表达[51],由此推测,EIN3/EIL1 通过负调控SID2基因的表达调节植物体SA 合成,进而调控PR基因表达, 影响植物抗病性。 本研究利用VIGS 技术在高抗枯萎病棉花品种中棉所12 中沉默GhEIN3基因,其沉默效率较高,且接菌试验表明沉默株系明显感病;对病程相关基因的表达量分析结果显示, 沉默株系中PR5表达量高于对照, 而PR1、PR2、PR4表达量均低于对照, 暗示GhEIN3基因能够调控PR基因表达。 但是,本研究采用的VIGS 技术只能降低GhEIN3基因的表达量(基因表达量约为未沉默株系的14%),无法完全抑制其表达,所以根据上述结果仅能初步推断GhEIN3基因参与并响应枯萎病侵染棉花的过程,而对于该基因是否如前人研究一样通过调控SA 合成关键基因SID2的表达来调节棉花对枯萎病的抗性有待后续研究。
ERF 是植物特异性转录因子家族乙烯响应元件结合蛋白 (ethylene responsive element binding protein, EREBP)的成员之一,离体和活体试验均证实EIN3 与特定ERF1启动子的原初响应元件结合形成二聚体,调节ERF1转录[26]。 本研究沉默GhEIN3基因的植株中,ERF1基因的表达低于对照, 由此推断GhEIN3基因对ERF1基因具有调控作用。ACO基因是编码ACC 氧化酶的关键基因,ACC 氧化酶是ET 合成过程中重要的氧化酶[52],而ACO 酶活性下降,可以降低内源乙烯合成速度[53]。 本研究结果显示,GhEIN3基因为ET 信号通路中的正调控因子, 且GhEIN3基因沉默株系中,ACO基因的表达量下降,由此推测GhEIN3基因可能通过调控ACO基因表达调节ACC 的合成影响棉花内源乙烯合成, 进而影响棉花对枯萎病的抗性。
本研究就GhEIN3在不同胁迫处理下的表达特征、GhEIN3沉默后植株的抗病性与GhEIN3沉默后抗病相关基因的表达做了初步研究,为解析棉花中GhEIN3参与调控棉花对枯萎病菌抗性的机理提供了新的基础信息; 植物中EIN3基因是2 个相关乙烯途径的重要基因, 而GhEIN3如何具体参与调控棉花响应枯萎病菌胁迫仍有待深入研究。
4 结论
本研究从陆地棉中鉴定出1 个新的能够响应枯萎病菌和激素胁迫的EIN3/EIL基因家族成员GhEIN3,全长1 842 bp,含有典型的EIN3 蛋白结构域, 能够响应枯萎病菌、ET、JA 和SA 胁迫, 经ET 与JA 诱导后,GhEIN3基因的整体表达水平分别显著上调和下调。 通过VIGS 技术沉默GhEIN3的棉株中PR1、PR2、ERF1、ACO基因表达水平显著降低,沉默棉株对枯萎病的抗性减弱,表明GhEIN3基因在棉花抗枯萎病的过程中起积极作用。
