基于高斯曲率最大化的蓝莓温室光照与CO2综合调控策略
2022-08-08徐立鸿刘辉辉蔚瑞华
徐立鸿 刘辉辉 蔚瑞华
(同济大学电子与信息工程学院, 上海 201804)
0 引言
蓝莓光合作用直接影响产量和品质。影响光合作用最主要的因素有:温度、光照强度和CO2浓度[1],研究表明随着温度上升蓝莓净光合速率先增大后减小[2],高温胁迫会降低酶的活性[3],进而降低蓝莓光合作用。补光和增施CO2则能够在一定程度上降低叶片气孔限制,提高作物净光合速率[4]。
目前CO2调控[5-9]和补光控制[10-13]方面的研究多以单因子进行,较少考虑对光照、CO2进行联合调控。针对光照、CO2联合调控,白京华[14]利用支持向量回归建立光合速率模型,并采用离散曲率获取不同温度下光照、CO2调控目标区域,为光照、CO2的联合调控提供了依据。辛萍萍[15]在适宜温度内研究了基于效益优先的光照、CO2协同调控目标值的获取方法,降低调控成本的情况下有效提升了作物的光合速率。单慧勇等[16]提出了在CO2资源受限下的光照、CO2耦合调控。光照、CO2调控方面的研究大多以黑盒模型的方法建立作物净光合速率模型,模型缺乏机理性和普适性。在研究光照、CO2单因子调控中,采用离散曲率方法计算光响应曲线和CO2响应曲线曲率最大点的方法,极有可能由于数据的波动而改变光响应曲线和CO2响应曲线的凹凸性,影响曲率正负,甚至可能发生曲率爆炸,无法得出正常的曲率最大值。采用机理模型进行建模一方面可以增强模型机理性和普适性,另一方面也可以避免数据的波动影响曲率的计算。
针对以上问题,本文通过采集不同温度、光照强度、CO2浓度嵌套下的蓝莓净光合速率数据,在不同温度下以直角双曲修正模型为光响应模型,建立带CO2修正的蓝莓净光合速率机理模型。根据不同温度下蓝莓净光合速率模型的高斯曲率函数构造适应度函数,采用粒子群算法对蓝莓净光合速率模型的高斯曲率函数进行最大值寻优,并计算对应的光照强度和CO2浓度。以此构建基于高斯曲率最大化的蓝莓温室光照、CO2综合调控策略,为蓝莓温室光照、CO2的高效节能调控提供依据。并采用本团队提出的温室环境多因子协调控制算法[17]对温湿度进行调控,使其满足蓝莓生长的温湿度要求,以在此基础上研究光照和CO2的综合调控策略。
1 材料与方法
1.1 试验材料
试验于2021年3—5月在昆山市永宏温室有限公司Venlo型温室内进行,试验温室坐标为(31°33′N,121°11′E)。选择3年生的南高丛蓝莓“绿宝石”作为试验品种。供试蓝莓种植于塑料盆内,选择泥炭土为主要原料的营养土作为栽培基质,土壤pH值约为5.2。试验期间按照常规的管理对蓝莓进行浇水、施肥、防虫等操作,不使用任何农药和激素。
1.2 试验方法
试验随机选择长势良好的蓝莓植株作为样本,采用美国LI-COR公司生产的Li-6400XT型便携式光合速率仪测定蓝莓叶片净光合速率。测量时间段为08:00—18:00,测量期间温室内相对湿度范围为45%~75%。使用便携式光合速率仪的不同子模块按照试验需要人工营造叶室环境。其中,利用控温模块构造16、20、24、28、32、36、40、44℃等8个温度梯度;利用LED光源模块(6400-02B)构造2 400、2 100、1 800、1 500、1 200、900、700、500、300、100、50、0 μmol/(m2·s)等12个光量子通量密度梯度;利用CO2注入模块(6400-01)设定14个CO2测试点:400、300、100、50、0、400、400、500、700、1 000、1 300、1 600、1 900、2 200 μmol/mol。由于CO2测试点从0 μmol/mol变为500 μmol/mol时植物所需的诱导时间较长,故增设2个400 μmol/mol的CO2测试点,以上2个CO2测试点不作实际建模数据使用,因此共1 152组试验。其中对同一株蓝莓叶片重复测量3次,剔除异常数据后求均值,共获得1 152个有效数据。
2 光合速率机理模型
基于实际采集数据建立不同温度下光照、CO2耦合的蓝莓净光合速率机理模型,为构建基于高斯曲率最大化的蓝莓温室光照、CO2综合调控策略提供模型基础。
2.1 常见的光合速率机理模型
直角双曲线模型、非直角双曲线模型[18]和负指数模型均可以用来描述光响应模型和CO2响应模型,但上述模型均不能有效反映光响应曲线和CO2响应曲线的饱和点[19],叶子飘等[20-21]在此基础上提出的直角双曲线修正模型则能较好地反映光响应曲线和CO2响应曲线的饱和点。常见的光响应模型和CO2响应模型如表1、2所示。
表1、2中αI为弱光条件下光响应曲线斜率或初始量子效率;PImax为强光作用下的最大净光合速率;I为光量子通量密度;θI(0<θI<1)为光响应曲线的凸度;βI为修正系数;γI为光响应曲线弯曲程度;αC为CO2响应曲线初始斜率;PCmax为高CO2浓度下最大净光合速率;C为CO2浓度;θC(0<θC<1)为CO2响应曲线的凸度;βC为修正系数;γC为CO2响应曲线弯曲程度;P为光合作用速率。

表1 常见的光响应模型Tab.1 Light response models

表2 常见的CO2响应模型Tab.2 CO2 response models
2.2 不同温度下光合速率机理模型
直角双曲线修正模型存在极值点,可以较好地估算饱和点光照强度、CO2浓度以及最大净光合速率,因此本文选择直角双曲线修正模型作为光响应模型对蓝莓净光合速率进行建模,并在该模型基础上加入CO2影响因子。基于直角双曲线修正模型的蓝莓净光合速率光响应模型表达式为
(1)
式中Pn——蓝莓净光合速率
rd——蓝莓呼吸速率
Blackman限制因子律[23]表明:其他环境因子对作物光合作用的影响可以在光响应模型的基础上乘以环境因子的修正函数。本文仅考虑CO2浓度和光量子通量密度对蓝莓净光合速率的影响,结合Blackman限制因子律,采取CO2响应函数模型进行修正。本文CO2响应模型选择直角双曲线修正模型。综上所述,建立包含CO2浓度和光量子通量密度的蓝莓净光合速率模型在不同温度T(16、20、24、28、32、36、40、44℃)下对蓝莓净光合速率模型Pn进行拟合。每组温度T下有144组样本,随机选取115组样本数组数据作为训练集(约占总样本数80%),剩下29组样本作为测试集(约占总样本数20%),采用拟牛顿法(BFGS)+通用全局优化法进行参数拟合,BFGS具有良好的自校正能力和超线性收敛性等特点,能有效避免通用全局优化算法计算复杂且难以收敛问题,拟合过程在1stOpt软件中进行。拟合得到的结果如表3所示。

表3 不同温度下蓝莓净光合速率模型参数Tab.3 Parameters of blueberry photosynthetic rate model at different temperatures
(2)
本文所建的不同温度下蓝莓净光合速率模型的评价指标如表4所示。

表4 不同温度下蓝莓净光合速率模型评价指标Tab.4 Evaluation index of blueberry photosynthetic rate model at different temperatures
由表4可知,平均绝对误差(MAE)在0.283 2~0.551 9 μmol/(m2·s)之间,平均相对误差(MRE)在0.085 9%~0.577 9%之间,均方根误差(RMSE)在0.379 1~0.716 6 μmol/(m2·s)之间,R2最低0.980 8。
不同温度下拟合得到的蓝莓净光合速率模型如图1所示。从图1可知,不同温度下的蓝莓净光合速率模型,在CO2浓度一定时,蓝莓净光合速率随着光子通量密度的增大而增大,当超过光饱和点时随着光子通量密度的增强蓝莓净光合速率呈现抑制现象;在光子通量密度一定时,蓝莓净光合速率随着CO2浓度的增大而增大,当超过CO2浓度饱和点时随着CO2浓度的继续增强,蓝莓净光合速率呈现抑制现象。

图1 不同温度下的蓝莓净光合速率模型Fig.1 Blueberry photosynthetic rate model at different temperatures
3 基于高斯曲率最大化的光照与CO2综合调控策略
3.1 蓝莓净光合速率模型的高斯曲率函数
在二维平面内,曲率为单位弧段切线转过角度的极限,表征了曲线上某点的弯曲程度。在CO2响应曲线中,常以曲线曲率最大点对应的CO2浓度来作为CO2浓度对作物净光合速率影响程度由强到弱的特征点[24]。以该特征点作为CO2调控目标点可以在满足作物光合作用需求的情况下最大程度地提升CO2调控效率。
将曲率的概念推广到三维,对于空间上一点,在任意方向上可以得到一个曲率。空间曲面上任意一点的曲率是描述三维模型形状的重要属性,它反映了点所在曲面的凹凸程度,具有旋转不变性和平移不变性[25]。过这一点,拥有无穷多个相互垂直的正交曲率,存在一组正交曲率使得这组正交曲率分别取极大极小值,则称这组正交曲率为主曲率。高斯曲率为主曲率的乘积。
蓝莓净光合速率与CO2浓度、光照强度不在同一量级内,需要将其进行归一化后再计算各响应曲面的高斯曲率。这里采用归一化函数进行归一化。归一化后的区间为[0,1],归一化函数为
y=(x-xmin)/(xmax-xmin)
(3)
式中y——归一化后数据
x——待归一化数据
xmax、xmin——待归一化数据的最大、最小值
不同温度下蓝莓净光合速率模型的高斯曲率函数kG计算式为
(4)
其中
(5)
(6)
(7)
(8)
(9)
(10)
3.2 基于粒子群算法的高斯曲率最大值寻优
粒子群算法是一种模拟鸟类觅食的随机搜索算法,其算法原理是对单个粒子在搜索空间内搜索最优解,并将其标记为当前个体的极值;将所有个体极值进行共享,找到最优的个体极值作为群体最优解[26];所有粒子根据当前的个体极值和群体极值进行更新。所有粒子具有2个属性:速度和位置,其中速度表征快慢,位置表征方向。粒子速度和位置更新公式为
vid(t+1)=ωvid(t)+c1r1(pid(t)-xid(t))+
c2r2(pgd(t)-xgd(t))
(11)
xid(t+1)=xid(t)+vid(t+1)
(i=1,2,…,N;d=1,2,…,D)
(12)
式中N——粒子群个数
D——空间维度
ω——惯性权重因子
c1——个体学习因子
c2——群体学习因子
r1、r2——介于(0,1)之间的随机数
pid——第i个粒子在d维的最优位置
pgd——整个粒子在d维的最优位置
vid——第i个粒子在d维的速度
xid——第i个粒子在d维的位置
惯性权重因子ω影响粒子搜索范围和精度:ω越大全局搜索能力越强,局部搜索能力越弱,容易导致收敛精度较低,ω越小局部搜索能力越强,全局搜索能力越弱,容易陷入局部最优。传统的ω优化采用线性递减的方式虽然可以加快收敛速度,但是收敛前期容易忽略最优值而导致全局搜索能力的降低[27]。本文采用先增后减的方式进行优化,前期ω增大过程可以有效避免陷入局部最优,后期ω减小过程可以加快收敛速度,具体方式为
(0.5≤ωmin<ωmax≤1)
(13)
式中ω0——惯性权重初值,取0.75
ki——当前迭代的次数
kmin——最小迭代次数
kmax——最大迭代次数
ωmin——惯性权重最小值
ωmax——惯性权重最大值
采用变化的学习因子避免粒子陷入局部最优,个体学习因子和群体学习因子的变化量为
(14)
(15)
式中cr1、cr2——个体学习因子和群体因子初始值
cmax1、cmax2——个体学习因子和群体因子的最大值
根据不同温度下蓝莓净光合速率模型的高斯曲率函数kG构造适应度函数
f(C,I)=-kG
(16)
粒子群算法流程如图2所示。
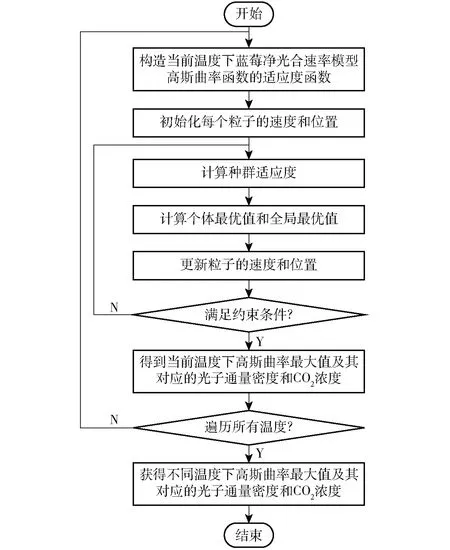
图2 粒子群算法流程图Fig.2 Flow diagram of PSO
计算得到不同温度下蓝莓净光合速率模型高斯曲率最大值及其对应的光照强度和CO2浓度点,计算结果如表5所示。
3.3 不同温度下光照、CO2综合调控策略建立
根据表5中的数据,采用多项式拟合建立不同温度下光照、CO2综合调控策略,拟合结果为
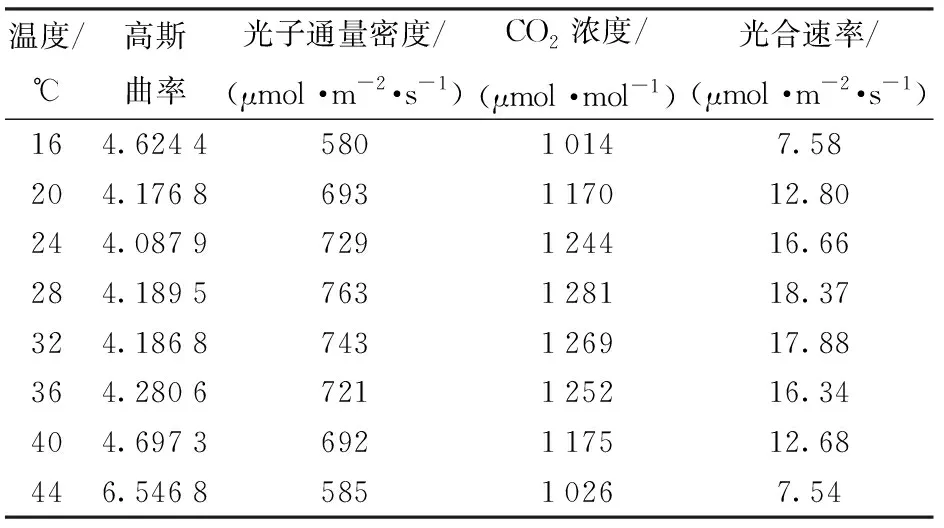
表5 不同温度下高斯曲率最大点及其对应的光照强度和CO2浓度Tab.5 Maximum point of Gaussian curvature and its corresponding light intensity and CO2 concentration at different temperatures
I=-2.317 2×10-3T4+0.284 0T3-13.447 6T2+
289.256 5T-1 616.884 8
(16℃≤T≤44℃)
(17)
C=-2.751 5×10-3T4+0.332 0T3-15.805 4T2+
349.023 6T-1 703.807 4
(16℃≤T≤44℃)
(18)
光照强度与温度的多项式拟合中R2为0.988 8,CO2浓度与温度的多项式拟合中R2为0.998 7,拟合效果如图3所示。
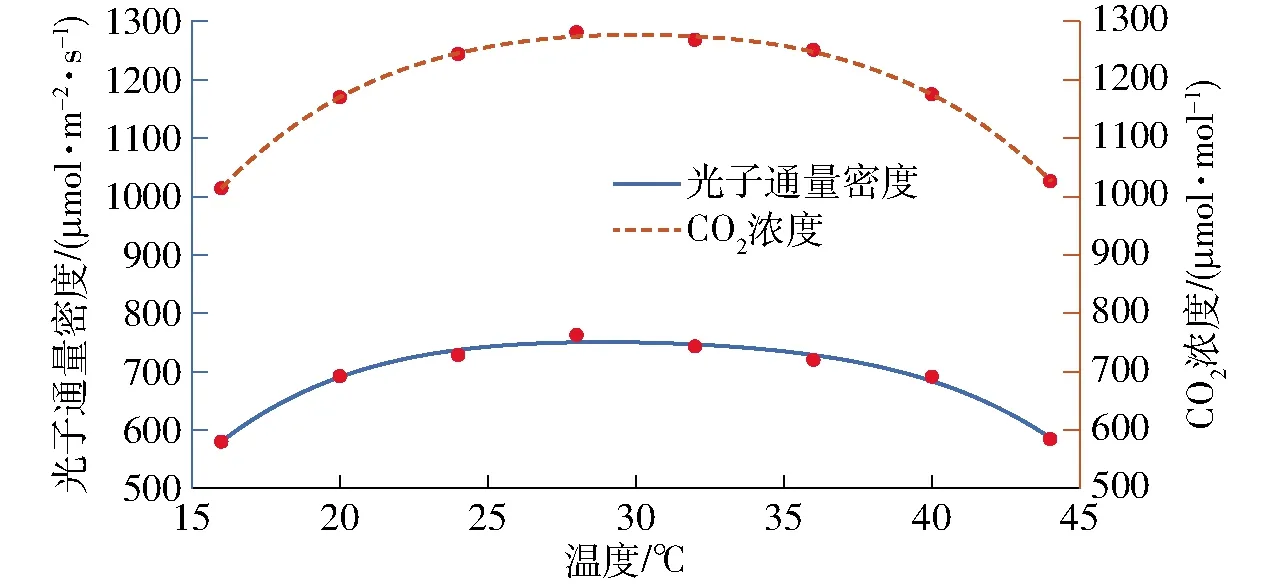
图3 光照强度、CO2浓度综合调控策略拟合曲线Fig.3 Integrated control strategy of light and CO2
4 结果与讨论
4.1 理论调控效果对比
为验证不同温度下光照与CO2综合调控策略的效果,本文与不同温度下蓝莓光合速率饱和点进行对比,对比结果如表6所示。基于高斯曲率最大化的调控相比于光合速率饱和点调控,平均光照强度下降60.73%,CO2浓度下降25.00%,而平均净光合速率仅下降14.29%。充分说明,本文采用的方法在牺牲少量光合速率的情况下,大幅减少了光照和CO2的调控阈值,有效地节约了光照和CO2的施放成本。该方法对于温室作物光照、CO2的调控具有高效节能的特征。
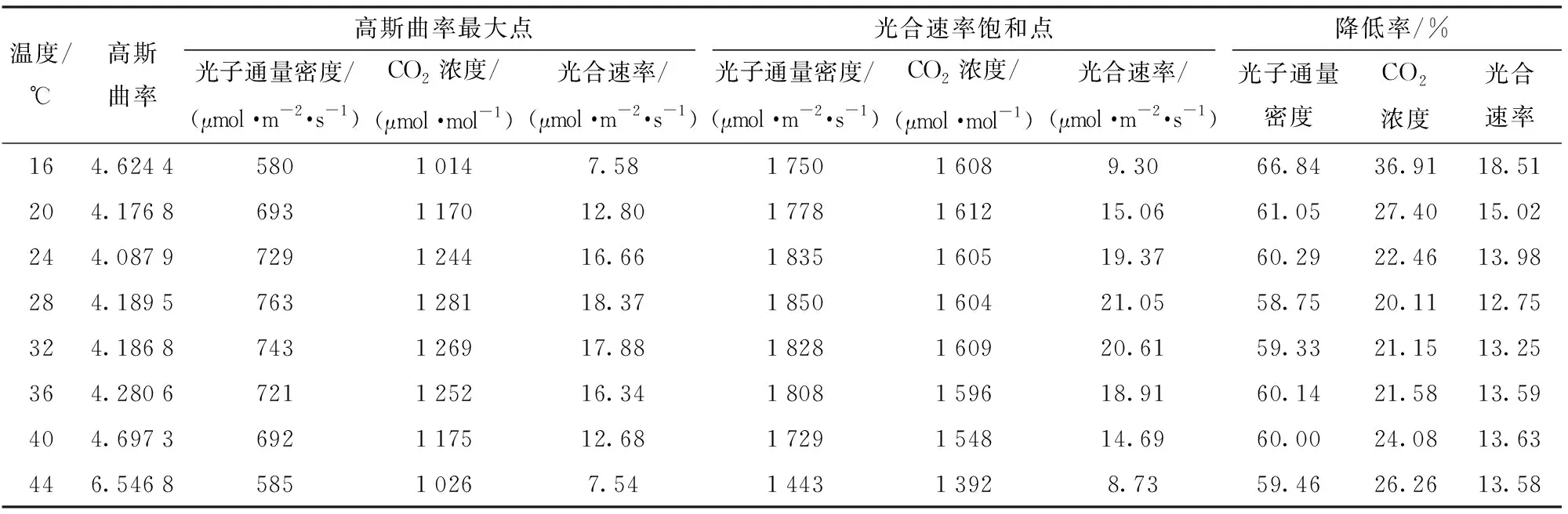
表6 高斯曲率最大化调控与光合速率饱和点对比Tab.6 Comparison of maximum control of Gaussian curvature and saturation point of photosynthetic rate
4.2 实际调控效果分析
为了验证本文提出的光照、CO2综合调控策略的实际效果,选择2021年5月28日(室外最低气温20℃,最高气温30℃,阴天)、5月29日(室外最低气温21℃,最高气温30℃,多云)2天进行手动调控。调控时间为08:00—18:00,调控时间段内室内相对湿度为45%~70%。试验以小时为单位进行调控,整点时刻,取前15 min室内温度、光照强度、CO2浓度均值作为当前时刻环境初始值。利用光照、CO2综合调控策略获取整点时刻不同温度下的光照、CO2调控目标值。采用Li-6400XT型便携式光合仪的控温模块、光源模块和CO2注入模块设定对应的温度、光照强度和CO2浓度测定蓝莓叶片的光合速率:当室内光照强度、CO2浓度低于调控目标值时,以调控目标值作为设定值;否则以当前环境光照强度、CO2浓度作为设定值。与光合速率饱和点进行对比,调控效果对比图如图4所示。从图4g、4h可以看出,采用本文建立的光照、CO2综合策略可以有效增大蓝莓净光合速率。相比于饱和点调控,平均光照强度分别降低59.44%(5月28日)、59.61%(5月29日),平均CO2浓度分别降低20.14%(5月28日)、20.36%(5月29日),而平均光合速率仅降低12.76%(5月28日)、11.85%(5月29日)。相比于自然环境下的光合速率,本文提出的调控策略平均光合速率提高了2.43倍(5月28日)、1.87倍(5月29日)。

图4 调控效果对比Fig.4 Control effect comparisons
5 结论
(1)以直角双曲线修正模型为基础,建立了不同温度下包含光照、CO2的蓝莓净光合速率模型,不同温度下模型评价指标中平均绝对误差在0.283 2~0.551 9 μmol/(m2·s)之间,平均相对误差在0.085 9%~0.577 9%之间,均方根误差(RMSE)在0.379 1~0.716 6 μmol/(m2·s)之间,R2达到0.980 8以上,模型精度较高,能较好地反映蓝莓净光合速率与光照强度、CO2浓度的变化关系。
(2)提出以高斯曲率最大时的光照强度、CO2浓度为调控目标值,采用粒子群算法进行高斯曲率最大值寻优,获得不同温度下的光照、CO2调控值。利用多项式拟合建立不同温度下蓝莓光照、CO2综合调控策略,并且光照、CO2综合调控策略精度较高,拟合结果中光照调控策略的R2为0.988 8,CO2调控策略的R2为0.998 7。
(3)在不同温度下将基于高斯曲率最大化的光照、CO2综合调控策略与蓝莓光合速率饱和点调控策略进行对比发现,采用本文提出的调控策略相比于光合速率饱和点调控策略,平均光照强度下降60.73%,CO2浓度下降25.00%,而平均净光合速率仅下降14.29%。说明本文提出的方法在牺牲少量光合速率的情况下,可以大幅减少光照、CO2的调控阈值,从而可以有效地节约光照、CO2的施放成本。通过与自然环境下的光合速率对比发现,采用本文提出的光照、CO2综合调控策略能大幅提升作物光合速率。充分说明,采用基于高斯曲率最大化的光照、CO2综合调控策略兼具高效节能的特征。
