低温胁迫对优良三色堇自交系幼苗耐寒性的影响
2022-08-08常珍惜杜晓华贾文庆朱小佩穆金艳刘会超
常珍惜,杜晓华,贾文庆,朱小佩,穆金艳,刘会超
(河南科技学院园艺园林学院,河南 新乡 453003)
三色堇(Viola ×wittrockiana)是堇菜科堇菜属的一、二年生草本花卉,其品种繁多、色彩鲜艳、花期较长,素有“花坛皇后”的美誉[1],自20 世纪八十年代以来,逐渐成为我国城乡重要的花坛和盆栽花卉之一.在我国长江流域,三色堇主要应用于秋、冬、春三季,而在我国北方,由于冬季气温较低,三色堇一般需要设施保护,春季才能应用于室外[2].研究三色堇资源的耐寒性,发掘耐寒三色堇种质资源,培育耐寒三色堇品种,对于解决三色堇在我国北方冬季越冬,丰富我国寒冷季节可应用的室外花卉具有重要意义.
前人研究表明,低温会给植物造成一定的胁迫和伤害,这些伤害体现在多个方面,其中细胞膜的伤害可用丙二醛含量变化、细胞相对电导率等来表征,其也可作为植物抗寒性鉴定的依据[3].植物在受到低温胁迫时,机体内可启动一系列生理生化反应来降低低温造成的伤害[4],包括渗透调节物质含量的变化以及抗氧化系统的变化.植物体内渗透调节物质,主要是有机小分子,如可溶性糖、可溶性蛋白、脯氨酸等[5].杨克彬等[6]研究发现,7 个切花红掌在6 ℃低温下,随着低温胁迫时间的延长,各品种的相对电导率、丙二醛及渗透调节物质含量逐渐上升,抗氧化酶呈现先升后降的结果.刘冰等[7]比较了3 种杜鹃花材料在杭州自然低温后,测定部分生理指标,结果表明:杜鹃花的抗寒性与蔗糖、可溶性蛋白、脯氨酸及丙二醛含量密切相关;许瑛等[8]在秋冬季自然环境下,通过测定8 个菊花品种的抗寒性,试验发现8 种菊花的半致死温度随驯化温度降低而降低,并验证生长恢复试验同半致死温度结果一致,结果表明半致死温度可以作为评价菊花抗寒性的重要指标之一.
有关三色堇耐寒性研究,罗玉兰研究了低温下1 个本地三色堇品种与1 个荷兰三色堇品种的相对电导率及膜损伤率等指标的变化,发现本地三色堇抗寒性低于荷兰三色堇品种[9];齐阳阳等[10]通过对三色堇进行-5 ℃低温胁迫发现,其可溶性糖、脯氨酸含量呈现先升后降的趋势.但上述研究仅从生理层面进行研究,缺乏植株直观表现.本研究以9 种三色堇优良自交系为材料,研究低温胁迫下三色堇渗透调节物质及膜系统指标的变化,结合田间耐寒性调查,探讨三色堇在低温下的生理响应以及与耐寒性的关系,为三色堇抗寒材料的筛选及耐寒新品种的选育提供理论参考.
1 材料与方法
1.1 试验材料
试验材料为河南科技学院三色堇课题组多年选育的9 份三色堇自交系,材料基本情况详见表1.
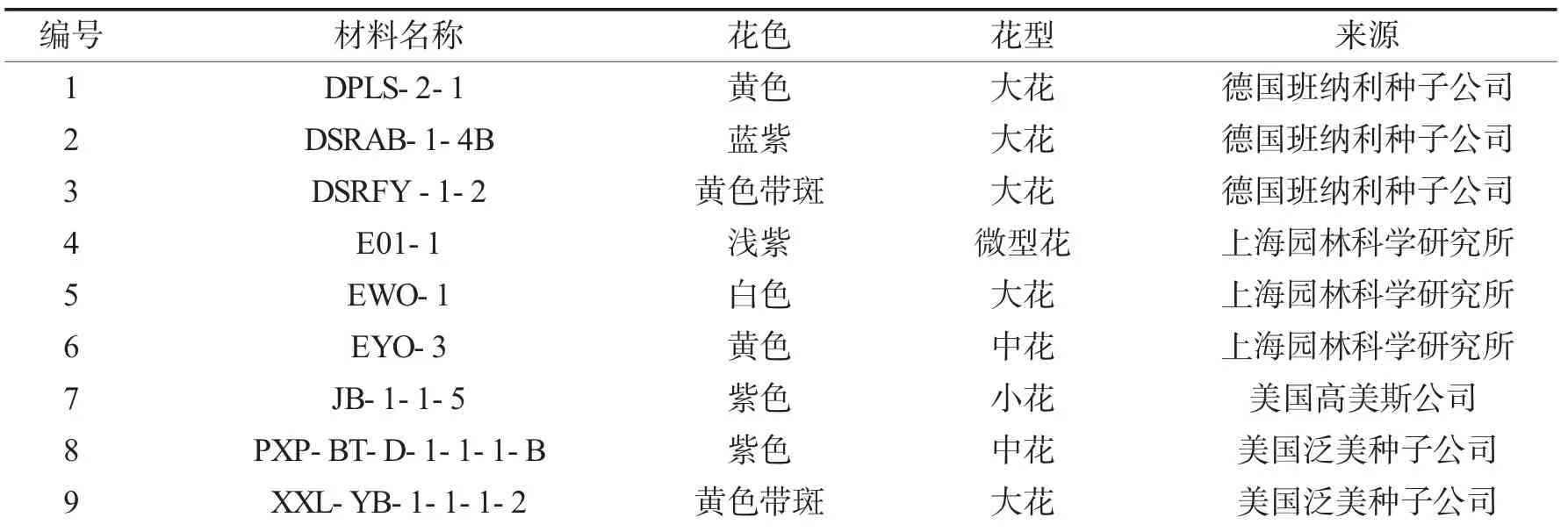
表1 供试的9 份三色堇自交系材料Tab.1 The nine inbred lines of pansies in this study
试验用苗为2020 年10 月上旬播种,幼苗长到7~8 片真叶时,将其分为两部分.一部分幼苗移栽至大田进行田间抗寒性试验,每品种栽植50 株,株行距30 cm×30 cm,田间温度测定采用温湿度记录仪;另一部分幼苗(每品种10 株)在人工气候箱内20 ℃下恒温培养24 h 后,进行人工模拟低温处理,温度每1 h降低1 ℃,待温度降至0 ℃时,培养24 h,光照强度10 000 lx,昼夜光照时间12 h/12 h,每株取1~2 片叶测定生理生化指标.
1.2 试验方法
1.2.1 三色堇田间抗寒性调查与冷害指数测定 依据新乡市气象局预报结合田间电子温度计测定,2020年12 月25 日至26 日,为试验区冬季最低气温(-2 ℃~ -8 ℃).低温胁迫24 h 后,对三色堇进行冷害调查.
三色堇田间冷害症状调查分级标准结合刘旭梅等[11]、李刚等[12]以及齐阳阳等[13]在杜鹃、常绿树种幼苗、大花三色堇等低温胁迫后,研究文献制定冷害分级.
冷害分级标准具体如下:
一级:植株正常无冻害;
二级:叶片呈现皱缩,有失绿;
三级:叶片皱缩小于50%,绿色较多;
四级:叶片皱缩大于50%,绿色较少;
五级:叶片皱缩,有坏死斑块,整体萎蔫;
六级:植株干枯.
冷害指数计算公式(1)
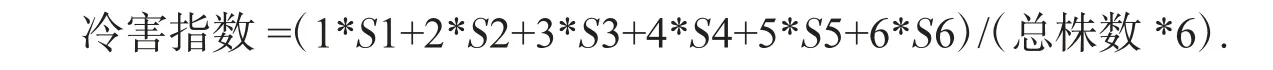
式(1)中:S 为每一冷害级的苗数,每材料调查30 株.
1.2.2 生理生化指标测定 采用Yayin1661 叶绿素荧光仪测定各材料叶绿素荧光参数[14],采用电导法测定相对电导率[15],采用茚三酮比色法测定游离脯氨酸含量[14-15],采用考马斯亮蓝法测定可溶性蛋白含量[15],采用苯酚比色法测定可溶性糖含量[15],采用硫代巴比妥酸法测定丙二醛含量[15],所有指标的测定均重复3 次.
1.2.3 抗寒性综合评价 参照龙海涛等[16],付晴晴等[17]的方法,用隶属函数标准差系数权重法进行9 个三色堇自交系抗寒性的综合评价.具体计算公式如下
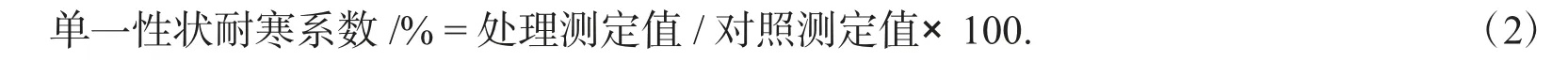
根据单一性状耐寒系数,使用SPSS 软件分析得出各综合指标主成分测定值CI(Xj);
隶属函数分析
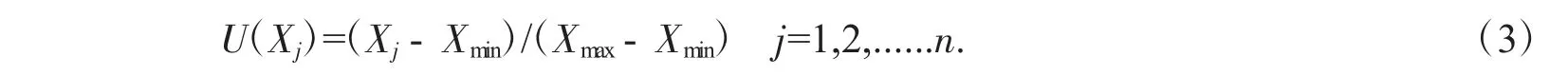
式(3)中:U(Xj)表示j 指标抗寒隶属函数值,Xj表示j 指标的测定值,Xmin表示9 种三色堇中j 指标最小值,Xmax表示9 种三色堇中j 指标最大值.
权重
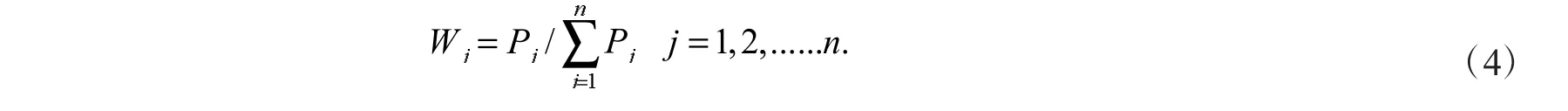
式(4)中:Wj值表示第j 个综合指标在所有综合指标中的重要程度,Pj为各综合指标的贡献率.
综合评价
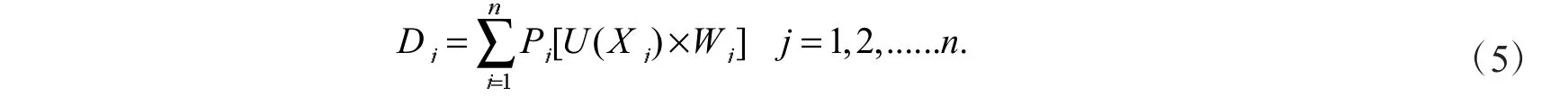
式(5)中:D 值为9 份三色堇材料在低温胁迫条件下,各生理指标所得的抗寒性综合评价值.
1.2.4 数据分析方法 采用Excel 2003 软件整理数据,并计算三色堇各指标隶属函数值和综合评价值等,利用DPS 7.5 和SPSS 25.0 软件数据处理系统,对数据进行统计和方差分析.
2 结果与分析
2.1 自然低温下9 份三色堇材料的冷害指数
供试的9 份三色堇材料冷害调查表见表2.
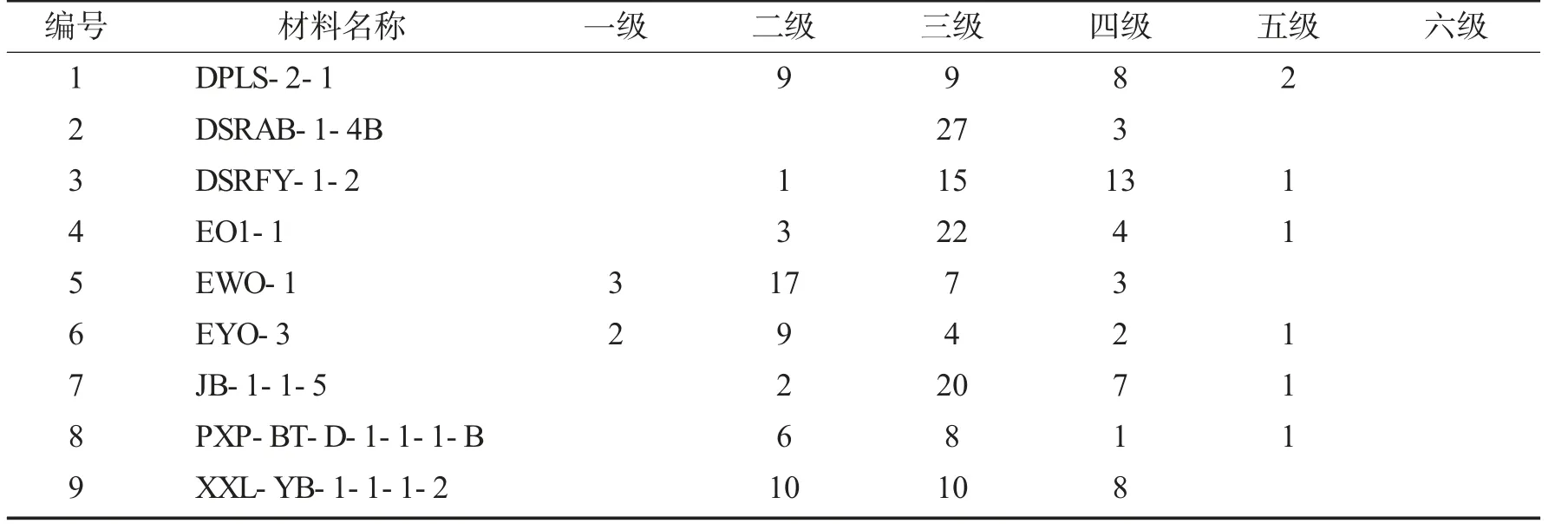
表2 供试的9 份三色堇材料冷害调查表Tab.2 Cold damage of 9 inbred lines of pansies 株
由表2 可以看出,9 份三色堇材料中EWO-1 一级和二级植株达20 株,所占比例最高,显示该材料耐寒性最强;其次是EYO-3 一级和二级植株为11 株;达到四级和五级标准最多的是DSRFY-1-2、DPLS-2-1、XXL-YB-1-1-1-2,分别为14 株、10 株及8 株.
供试的9 份三色堇材料进行大田耐寒性分析如图1 所示.
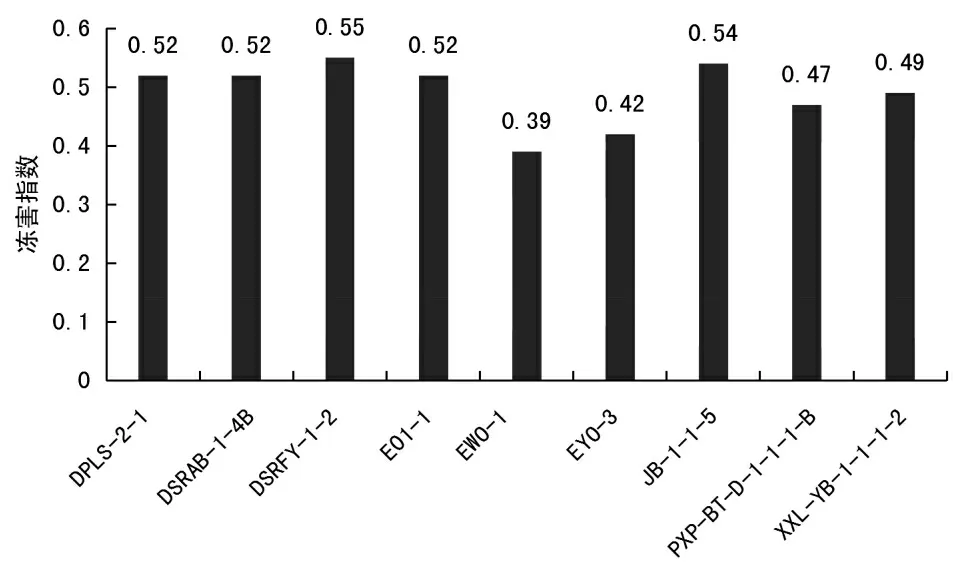
图1 三色堇材料冷害指数测定Fig.1 Chilling injury index of each material after lowtemperature stress
如图1 可知,低温对9 种三色堇材料造成不同程度的伤害,其冷害指数在0.39~0.55 之间.9 份材料中,EWO-1 的冷害指数值最低,为0.39,其次为EYO-3,为0.42,冷害指数值最高的为DSRFY-1-2 材料,达到0.55.反映出EWO-1 的耐寒性最强,其次是EYO-3,最不耐寒材料为DSRFY-1-2.
2.2 低温胁迫下9 份三色堇自交系生理生化指标的变化
低温胁迫下对三色堇生理生化指标的影响如图2 所示.
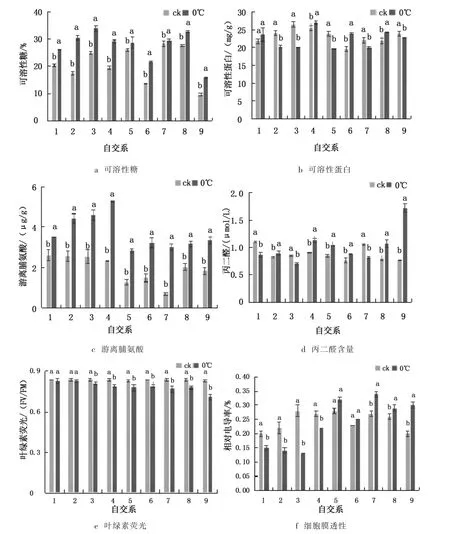
图2 低温胁迫三色堇生理生化指标的影响Fig.2 Effects of lowtemperature on some physiological and biochemical indexes of pansies
由图2-a 可知, 同对照相比,0 ℃处理后各材料可溶性糖含量增加, 除EWO-1 和JB-1-1-5 两个材料,另外7 种材料与对照相比,存在显著性差异(P<0.05,下同),其中增加最多材料为DSRAB-1-4B,增长了74.76%.由图2-b 可知, 低温胁迫导致不同三色堇材料可溶性蛋白含量出现一定程度上升或下降趋势.除DPLS-2-1 外,低温处理使其他8 份三色堇材料可溶性蛋白含量与对照相比呈现出显著性差异.其中E01-1、EYO-3、PXP-BT-D-1-1-1-B 的可溶性蛋白含量上升,上升最大为EYO-3 材料,比对照上升了4.28 mg/g;另外5 个三色堇材料中可溶性蛋白下降,下降最大为DSRFY-1-2,比对照下降了6.43 mg/g.由图2-c 可知,低温胁迫后,9 份三色堇材料游离脯氨酸含量与对照相比均有显著性上升,其中上升最高材料为JB-1-1-5,比对照增长了346.27%,E01-1、EWO-1、EYO-3,分别增长129.57%、124.6%、116.89%,增长最少为DPLS-2-1 材料,增长了35%.由图2-d 可知,低温胁迫后,9 份三色堇材料中,6 份材料丙二醛含量上升,3 种下降,差异显著.上升最高为XXL-YB-1-1-1-2 材料,上升了0.96 μmol/g,比对照增加了126.32%,其次为PXP-BT-D-1-1-1-B 和E01-1 材料,分别提高了38.96%、24.18%.下降幅度最大材料为JB-1-1-5 和DPLS-2-1,分别下降了22.86%、20.91%.由图2-e 可以看出,低温胁迫后,除DPLS-2-1、DSRAB-1-2 外,其他7 份材料的叶绿素荧光(Fv/Fm)均表现出显著性下降,下降幅度最大为XXL-YB-1-1-1-2 材料,下降了14.46%,下降最少为DPLS-2-1 和DSRAB-1-2 材料,均下降了1.19%.由图2-f 可知,在低温胁迫后,除EWO-1 和EYO-3 材料外,其他7 份三色堇材料同对照相比相对电导率均有显著性上升或下降.其中DSRFY-1-2、DSRAB-1-4B、DPLS-2-1、E01-1 材料下降,分别下降了53.57%,36.36%、25%、18.52%;EWO-1、EYO-3、JB-1-1-5、PXP-BT-D-1-1-1-B、XXL-YB-1-1-1-25 上升,XXL-YB-1-1-1-2 上升最高,达到50%.
2.3 9 份三色堇自交系材料的耐寒性综合评价
2.3.1 主成分分析6 个单项指标的主成分分析结果见表3.
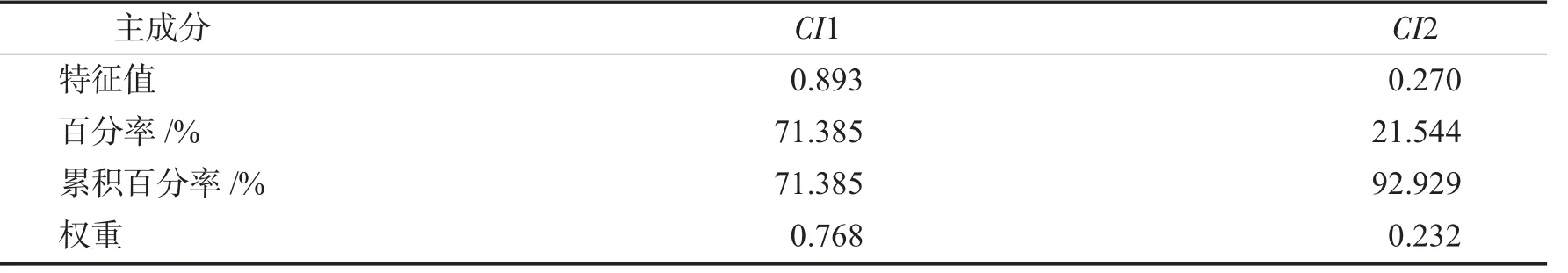
表3 低温胁迫下2 个综合指标特征量及贡献率Tab.3 Characteristics and contribution rate of three comprehensive indexes of pansies under lowtemperature stress
由表3 可知,6 个单项指标贡献率分别为:71.385%(Pj1)、21.544%(Pj2)、4.530%(Pj3)、2.150%(Pj4)、0.382%(Pj5)、0.009%(Pj6).其中2 个主要综合指标(CI1、CI2)的贡献率分别为71.385%、21.544%,累积贡献率为92.929%,可以代表原始6 个单一评价指标,可依据这2 个综合指标进行9 种三色堇材料的耐寒性评价.
2.3.2 抗寒性综合评价 9 种三色堇材料主成分综合指标、U(xj)、D 值及耐寒性排序见表4.

表4 9 种三色堇材料主成分综合指标、U(xj)、D 值及耐寒性排序Tab.4 Comprehensive index,U(xj),D value and cold resistance of 9 inbred lines of pansies
由表4 可知,利用两个主成分综合指标值得出隶属函数综合评价值(D 值),D 值愈大其耐寒能力愈强,故9 种三色堇材料在0 ℃胁迫下的耐寒性依次为:XXL-YB-1-1-1-2>PXP-BT-D-1-1-1-B>EWO-1>EYO-3>JB-1-1-5>E01-1>DSRAB-1-4B>DPLS-2-1>DSRFY-1-2.
3 结论与讨论
本试验研究表明,低温胁迫下,9 份三色堇材料的部分生理生化指标产生上升/下降的结果.抗寒性综合评价表明,9 份材料抗寒能力由强到弱依次为:XXL-YB-1-1-1-2>PXP-BT-D-1-1-1-B>EWO-1>EYO-3>JB-1-1-5>E01-1>DSRAB-1-4B>DPLS-2-1>DSRFY-1-2;大田冷害调查结果9 份材料抗寒能力由强到弱依次为:EWO-1>EYO-3>PXP-BT-D-1-1-1-B>XXL-YB-1-1-1-2>DPLS-2-1=DSRAB-1-4B=E01-1>JB-1-1-5>DSRFY-1-2.综合上述两种评价方法,9 份材料中,较为耐寒材料:PXP-BT-D-1-1-1-B、EWO-1、EYO-3,不耐寒材料:JB-1-1-5 和DSRFY-1-2.
低温处理条件下因植物种类不同,在低温胁迫后,渗透调节物质变化规律有所差异.齐阳阳等[10]以三色堇离体叶片为试材,随着低温胁迫时间的增加,三色堇叶片的脯氨酸、可溶性糖含量呈现先升后降趋势.张雨等[18]通过对低温胁迫下板凳果生理指标的研究,发现板凳果体内MDA、可溶性蛋白和相对电导率呈上升结果.何伟等[19]探索葡萄半致死低温时发现,葡萄随着低温时间的增加,可溶性糖、可溶性蛋白均先升后降,而游离脯氨酸含量呈现缓升缓降、MDA 呈现平缓上升结果.本试验中,低温胁迫下9 份三色堇材料的可溶性糖含量整体提高,且DSRAB-1-4B、XXL-YB-1-1-1-2、EYO-3、E01-1、DSRFY-1-2 等5种材料显著提高;可溶性蛋白含量4 份材料下降,5 份材料上升;游离脯氨酸含量3 份材料下降,6 份材料上升,上升的6 份材料其耐寒性也较高,与前人研究报道结果相近,这进一步说明低温胁迫下,三色堇抗寒生理表现过程中,可溶性糖起到了更加关键作用,脯氨酸和可溶性蛋白也起到了一定作用.
植物体内细胞膜起着维持其正常代谢的重要作用,当植物受到胁迫后,细胞膜受损,透性增大,使得电解质外渗,引起植物细胞浸提液电导率增大[15].本试验中,与对照相比,0℃处理后,9 份材料相对电导率值出现增大或减小的变化,电导率明显上升的自交系XXL-YB-1-1-1-2、JB-1-1-5、EWO-1、EYO-3、PXP-BT-D-1-1-1-B 等材料,其抗寒性综合值也较高,反之,电导率明显降低的材料,其抗寒性综合值也较低.MDA 是反映细胞膜破坏程度的另一指标,用来表现植物的损伤程度[20],当植物体细胞膜受到的损伤越大,其体内MDA 的含量就会越多[21],本试验结果表明,低温胁迫后,DPLS-2-1、DSRFY-1-2、JB-1-1-5 3 种材料的MDA 值上升,表明这3 份三色堇材料细胞膜损伤较大,抗寒性低于另外6 份三色堇材料.
光合作用是维持植物正常生长发育的重要因素之一,且对温度的变化较为敏感,温度的上升或下降会影响光合效果[22],叶绿素荧光参数是测定叶片在光合作用过程中自身对光能吸收、传递、耗散等光合能力的一种方法[23-25].本试验在低温胁迫后得出9 种三色堇材料光合能力在0℃条件下均有下降,表示低温一定程度抑制了光合作用,但分析表明,叶绿素荧光系数与抗寒性贡献率较小,可以不作为抗寒性指标.
刘国宇等[26]通过对3 种热带观赏植物抗寒性研究发现,可溶性糖含量、游离脯氨酸含量与植物抗寒性呈正相关,结合本次试验结果分析发现,印证了上述刘国宇等人试验结果,可溶性糖和游离脯氨酸含量低温后呈现升高,是植物应对低温的防御反应,可能与抗寒性是正相关的,丙二醛含量以及叶绿素荧光指数多数材料下降,可能与抗寒性成负相关.
