Transcranial magnetic stimulation in animal models of neurodegeneration
2022-08-08MohammadUzairTurkiAbualaitMuhammadArshadWooKyoungYooAliMirReemFahdBunyanShahidBashir
Mohammad Uzair,Turki Abualait,Muhammad Arshad,Woo-Kyoung Yoo,Ali Mir,Reem Fahd Bunyan,Shahid Bashir,
Abstract Brain stimulation techniques offer powerful means of modulating the physiology of specific neural structures. In recent years,non-invasive brain stimulation techniques,such as transcranial magnetic stimulation (TMS) and transcranial direct current stimulation,have emerged as therapeutic tools for neurology and neuroscience. However,the possible repercussions of these techniques remain unclear,and there are few reports on the incisive recovery mechanisms through brain stimulation. Although several studies have recommended the use of non-invasive brain stimulation in clinical neuroscience,with a special emphasis on TMS,the suggested mechanisms of action have not been confirmed directly at the neural level. Insights into the neural mechanisms of non-invasive brain stimulation would unveil the strategies necessary to enhance the safety and efficacy of this progressive approach. Therefore,animal studies investigating the mechanisms of TMSinduced recovery at the neural level are crucial for the elaboration of non-invasive brain stimulation. Translational research done using animal models has several advantages and is able to investigate knowledge gaps by directly targeting neuronal levels. In this review,we have discussed the role of TMS in different animal models,the impact of animal studies on various disease states,and the findings regarding brain function of animal models after TMS in pharmacology research.
Key Words: Alzheimer’s disease; depression; glial cells; neurorehabilitation; Parkinson’s disease; repetitive transcranial magnetic stimulation; transcranial direct current stimulation; transcranial magnetic stimulation
Introduction
Over the last two decades,understanding human cognition by non-invasive brain stimulation (NIBS) has received much attention and has advanced considerably. Hitzig (1838-1907),Fritsch (1838-1927),Ferrier (1843-1928),Sherrington (1856-1952),and Penfield (1976-1991) have significantly contributed to research on the electrical stimulation of the cortex to discover the organization and physiology of the human brain. In 1959,Kolin et al. stimulated the nerves of frogs through alternating magnetic fields for the very first time. Later in 1965,Bickford applied magnetic fields to stimulate the human peripheral nerve. This work was not pursued further because the long-lasting activation interval made it impossible to record the action potential of the nerve or muscles (Bickford and Fremming,1965). Magnetic stimulation of frog nerves was also reported by Öberg (1973). However,in the following years,the technique was investigated only sporadically. In 1980,Merton and Morton (1980) electrically stimulated the motor cortex through the scalp using transcranial electrical stimulation (TES). In 1982,Polson et al. reported the first successful magnetic stimulation of the peripheral nerve. They applied 2-ms-duration pulses over median nerve magnetic stimulation and recorded the motor-evoked potentials (MEPs). TES became very useful for many purposes,but it was very painful. Later,in 1985,Barker et al. introduced transcranial magnetic stimulation (TMS) to stimulate the brain with no pain.
In this review,we provide an overview of NIBS while focusing mainly on TMS as an efficient method for studying cortical functions in different animal models. We also discuss the contribution of animal TMS studies validated for the treatment of human diseases and understanding the basics of neurophysiology and neurotransmission.
Database Search Strategy
The Cochrane Library,PubMed,EMBASE and Web of Science were searched to retrieve papers published up to 2020. Literature retrieval was performed using all possible ways including the reference lists and authors’ files from the included studies,in the protocol. A wide range of terms and related truncations including brain stimulation (BS),non-invasive brain stimulation (NIBS),transcranial magnetic stimulation (TMS),repetitive transcranial magnetic stimulation (rTMS),paired-pulse TMS,neurodegenerative diseases,neurodegenerative disorders,Alzheimer’s disease (AD),Parkinson’s disease (PD),Huntington’s disease (HD),depression,major depression,major depressive disorder,animals,animal models,mice,rats,monkeys,cognitive decline,cognitive function,neurocognitive,apoptosis,neurotransmitters,neurotrophic factors,BDNF,NMDA,NMDAR,GABA,glutamate,oligodendrocytes,astrocytes,microglia,and neural stem cells were used. Combinations of Medical Subject Headings (MeSH) and text words were used. The same search terms were also used in other electronic databases.
What Is Transcranial Magnetic Stimulation?
TMS is a non-invasive tool for brain stimulation that electrically stimulates neural tissues,including the cerebral cortex,spinal nerves,cranial and peripheral nerves. TMS can potentially depolarize neurons and effectively evoke measurable effects. It does not require surgery,anesthesia,injections,or intravenous systems,and it causes little or no pain. TMS is performed based on Faraday’s principle of electromagnetic induction,established in 1838. When a high-voltage current passes through a coil,a varying magnetic field is generated. Near the scalp,this time-changing magnetic field induces currents opposite in direction to the original current in the coil. When the magnetic coil is positioned near the skull and a sufficiently strong stimulus is administered,the underlying brain tissue is stimulated (Cuypers and Marsman,2020). TMS can be applied as single pulses of stimulation,or pairs of stimuli separated by variable intervals,or trains of repetitive stimuli at various frequencies to the same or different brain regions. Single-pulse TMS (sTMS) can be used to analyze central motor conduction time and motor cortical outputs,while paired-pulse TMS (ppTMS) can efficiently measure intracortical facilitation,inhibition,and impart significant effects on neuroplasticity (Cui et al.,2020). Trains of stimuli (repetitive TMS,rTMS) can regulate cerebral cortex excitability at the stimulated site and in remote areas,along with functional anatomical landmarks. rTMS has diverse effects on cognitive tasks,behavioral functions,and brain activity that depend on the stimuli frequency.
In recent times,TMS has widely become the wellestablished technique for the examination and modulation of the excitability of human brain areas,yet the basic cellular processes and mechanism behind this method are still unknown. Despite these anomalies,the prospective applications of TMS are enormous,particularly in the determination of the pathophysiology of the brain due to neurological and psychological disorders,and can be further developed and employed into clinically useful diagnostic and prognostic procedures,as well as in the therapeutic measures of various diseases. This tremendous potential of TMS has already been reported by numerous researchers,and requires many clinical trials on model animals to elaborate on the underlying mechanisms and yield options for novel therapeutic approaches.
Paired-pulse TMS
In this method,ppTMS utilizes two successive pulse stimuli via the same coil,with a short inter-stimulus interval (ISI) for a few milliseconds and a long ISI for tens to hundreds of milliseconds (Klomjai et al.,2015). Both of these pulses are delivered to the same regions of the dominant hemisphere to determine the inter-hemispheric inhibition or transcallosal inhibition (Cosentino et al.,2018). ppTMS methods are used to examine the inhibitory or excitatory intracortical networks that are stimulated by the intensity and ISI used (Massé-Alarie et al.,2016; de Goede et al.,2020; Neige et al.,2020). The intensity of the first pulse,or conditioning stimulus,is applied at the sub-motor threshold. The intensity of the second pulse,also called the test stimulus,is applied at the supra-motor threshold (Premoli et al.,2018). The test stimulus induces MEPs,which are thought to be an outcome of a conditioning pulse. Thus,cortical excitability can be evaluated by use of a second stimulus (Kallioniemi et al.,2018).
Repetitive TMS
rTMS offers a safe and non-invasive technique that modulates cortical excitability,function,and inhibition. Moreover,rTMS causes neuronal polarization in the brain and has the potential to treat neurological and psychiatric disorders. The modulation of cortical excitability and disturbance in cortical network function depends on the stimulation parameters (Hanlon et al.,2018; Beynel et al.,2019; Singh et al.,2020). rTMS induces trains of electric currents to the brain at a depth of 2 cm,which are delivered through the pulsating magnetic fields. rTMS produces intermittent magnetic fields in the range of 1.5 to 2 teslas (Klomjai et al.,2015; Singh et al.,2020). The induced currents modulate neuronal excitability with frequencies ≥ 5 Hz (i.e. high-frequency rTMS (HF-rTMS)) or ≤ 1 Hz (i.e. low-frequency rTMS (LF-rTMS)). In contrast to sTMS,rTMS can regulate cortical activity following the stimulation frequency (Lage et al.,2016; Hanlon et al.,2018). High-frequency rTMS appears to increase cortical excitability,MEP size,and intercellular interactions (ICI),whereas lowfrequency rTMS significantly decreases cortical excitability and enhances post-stroke motor recovery (Lage et al.,2016; Du et al.,2019). The recent literature shows that rTMS exerts long-lasting effects on cortical functions,as compared to nonrepetitive TMS (Chervyakov et al.,2015; Caglayan et al.,2019). These studies have revealed that increases in cell proliferation,regional cerebral blood flow,and synaptic plasticity are observed in neuronal circuits after high-frequency rTMS therapy. High-frequency rTMS also activates various cell signaling pathways,including brain-derived neurotrophic factor (BDNF),cyclic adenosine monophosphate-responsive element binding protein (CREB),and phosphatidylinositol 3-kinase/protein kinase B (Akt),to inhibit apoptotic cell death and promote neurogenesis following ischemia (Baek et al.,2018; Caglayan et al.,2019). Additionally,rTMS provides a non-invasive and effective neurorehabilitation approach for stimulating neurons and treating various neurological disorders.
The Molecular Mechanism Behind the Therapeutic Effects of Transcranial Magnetic Stimulation
TMS is an approachable and promising neuromodulation technique due to its ability to promote functional recovery of cortical function following a stroke (Agarwal et al.,2019; Caglayan et al.,2019). TMS promotes the recovery of the axonal terminals,structural integrity,cortical function,neuromodulation,affects gene expression and neurotransmitters,and produces neuronal recovery after a stroke by the action of the alternating magnetic field. TMS affects the stimulated cortical regions and other connected regions (Chervyakov et al.,2015; Baek et al.,2018; Aberra et al.,2020). However,although numerous studies have investigated the beneficial effects of TMS on brain stimulation,the precise mechanism behind the contribution of TMS remains incompletely understood.
TMS inhibits apoptotic cell death,prevents neuronal death,and promotes neuronal survival
In neurodegenerative conditions,including stroke,trauma,severe epileptic seizures,Alzheimer’s disease (AD),and motor system disorders,there is an increase in neuronal cell death. It is believed that apoptosis acts as a major cell death pathway in neurons (Chi et al.,2018; Pemberton et al.,2021),particularly neurons in the ischemic region. TMS prevents neuronal death by inhibiting several members of the B-cell lymphoma 2 (Bcl-2) family,particularly those that stimulate apoptosis,i.e.,Bad,Bax,and Bcl-xS (Kale et al.,2018; Pemberton et al.,2021). Numerous studies using animal models of transient ischemia and prolonged ischemia found that TMS protects neurons and inhibits programmed cell death following an ischemic event. rTMS treatment recovers neuronal functioning after cerebral ischemia/reperfusion injuries,as observed in rat models (Feng et al.,2005; Guo et al.,2014,2017). Gao et al. (2010) studied the neuroprotective effects of HF-rTMS in a transient ischemic mouse model to investigate the mechanism underlying these effects. They found that glucose metabolism was significantly higher,and caspase-3 positive cells were reduced following rTMS treatment. rTMS activates Bcl-2 cells and decreases Bax,indicating that rTMS inhibits apoptotic pathways in the ischemic region (Gao et al.,2010; Yoon et al.,2011). Caspase-3 significantly contributes to the apoptotic pathway and promotes cell death after ischemia. It has been noticed that blocking caspase-3 tends to reduce the damaged brain area,infarct volume,and lesion size by seizing apoptosis (Namura et al.,1998;Figure 1). Ke et al. (2010) studied the impact of LFrTMS on neuronal protein expression in the hippocampus of a mouse model of epilepsy. They concluded that low-frequency (0.5 Hz) rTMS exerted anti-apoptotic and anti-epileptic effects,as it upregulated Bcl-2 expression and Fas protein expression decreased. Protein levels of Bax-induced neuronal cell death,cleaved caspase-1,and cleaved caspase-3/caspase-3 decrease after LF-rTMS treatment (Guo et al.,2017; Luo et al.,2017; Baek et al.,2018; Caglayan et al.,2019). Prolonged exposure to rTMS significantly increases BDNF mRNA levels following ischemia,which may contribute to synaptic plasticity (Müller et al.,2000; Ogiue-Ikeda et al.,2005). Additionally,rTMS significantly activates Bcl-2,decreases Bax,and the number of TUNEL-positive cells in the ischemic hippocampus (Guo et al.,2017). Hence,many studies suggest that TMS exerts its beneficial effects by blocking apoptosis,mainly by regulating the expressions of BDNF,Bcl-2,and Bax (Table 1).
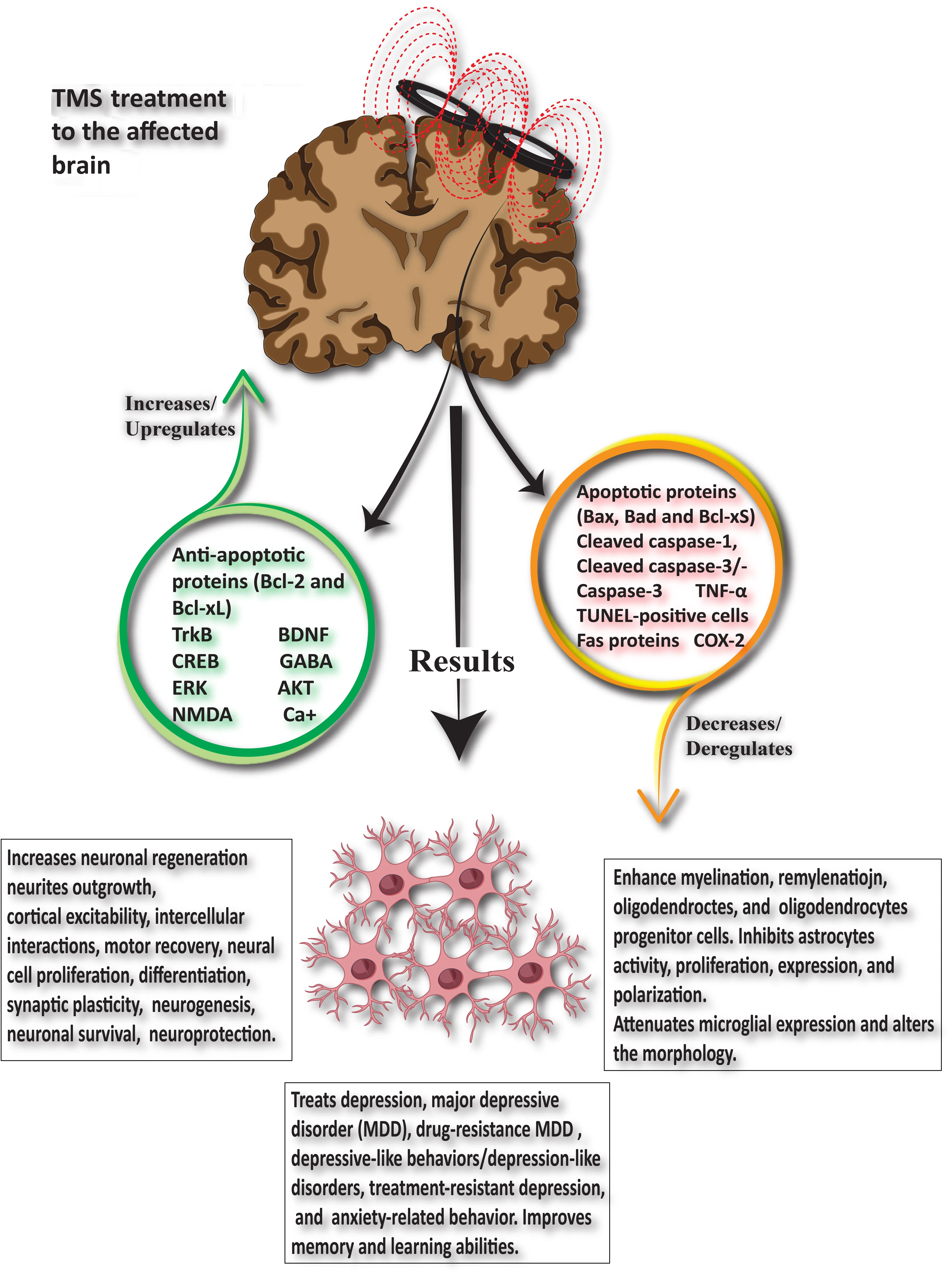
Figure 1|The beneficial effects of TMS on neuronal recovery.
TMS regulates neurotransmitters,neurotrophic factors,and exerts neuroprotective effects
Neurons may be damaged by either stroke,ischemia or neurodegenerative disorders,but TMS has the potential to treat them due to its neuroprotective and neural regenerative effects. Numerous studies have investigated the changes in dopamine synthesis and release following TMS treatment (Table 2). A study on 6-OHDA-induced rat models of Parkinson’s disease (PD) showed that a 0.5 Hz rTMS treatment produced a neuroprotective effect on the dopaminergic neurons (Yang et al.,2010). Thus,more dopaminergic cells survived in the rTMS-treated rats,resulting in significantly increased dopamine (DA) levels (Yang et al.,2010). Another study also concluded that acute rTMS (25 Hz) increases dopamine production in the dorsolateral prefrontal cortex,providing beneficial effects on motor symptoms (Kanno et al.,2004). Low-frequency rTMS has been shown to increase the number of tyrosine hydroxylase-positive dopaminergic neurons,the levels of extracellular dopamine and its metabolites,3,4-dihydroxyphenylacetic acid and homovanillic acid,in Parkinsonian rats (Ba et al.,2017). The functional neuroimaging study by Ko et al. (2008) on healthy volunteers investigated the effects of continuous theta-burst stimulation (cTBS) on dorsolateral prefrontal cortex (DLPFC) stimulation. They found that theta-burst (high-frequency) stimulus delivered to the left DLPFC deteriorated motor function and reduced bilateral striatal dopamine production,particularly in the ipsilateral caudate nucleus and ipsilateral putamen (Ko et al.,2008).
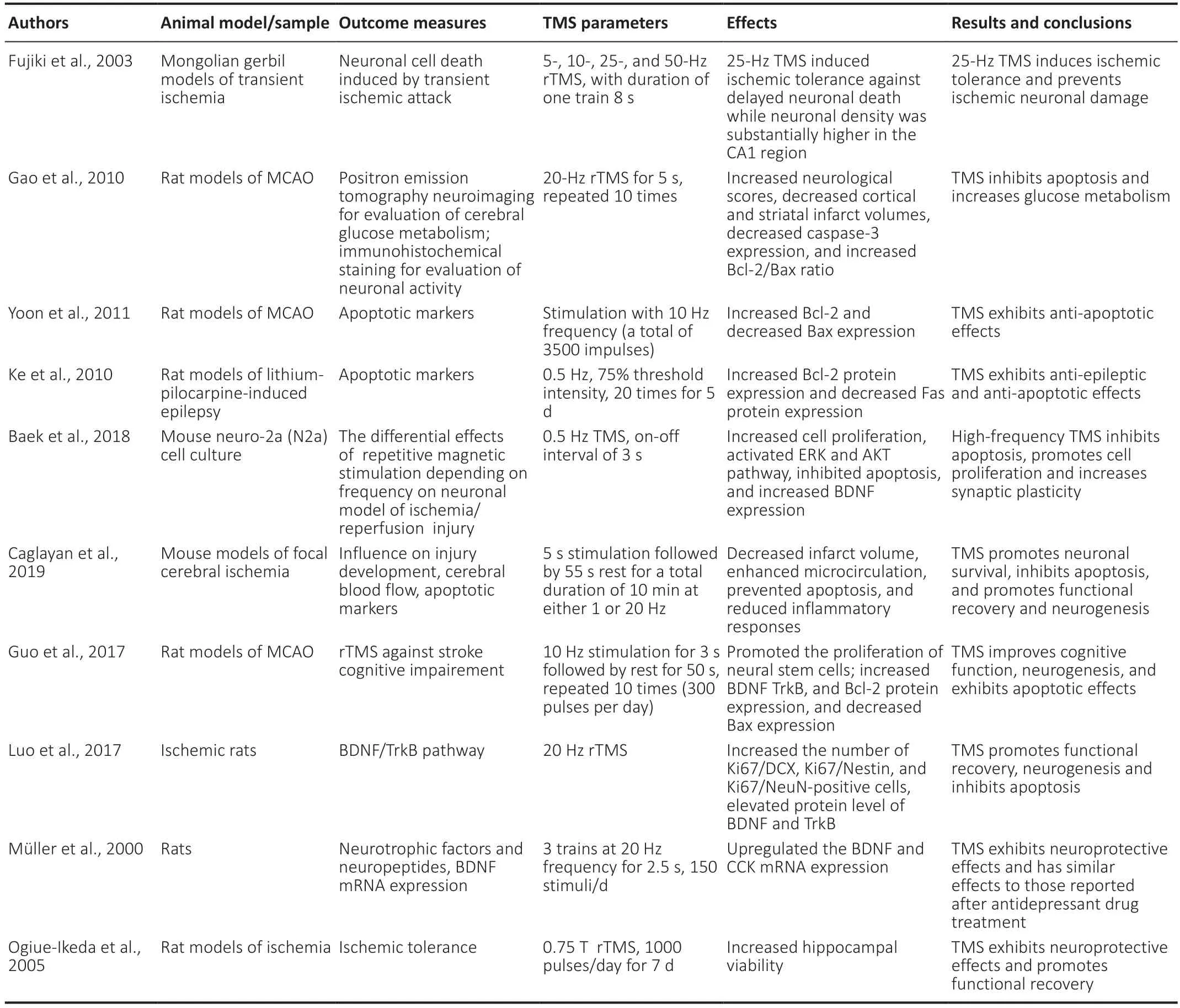
Table 1|Transcranial magnetic stimulation (TMS) parameters and its beneficial therapeutic effects on the brain
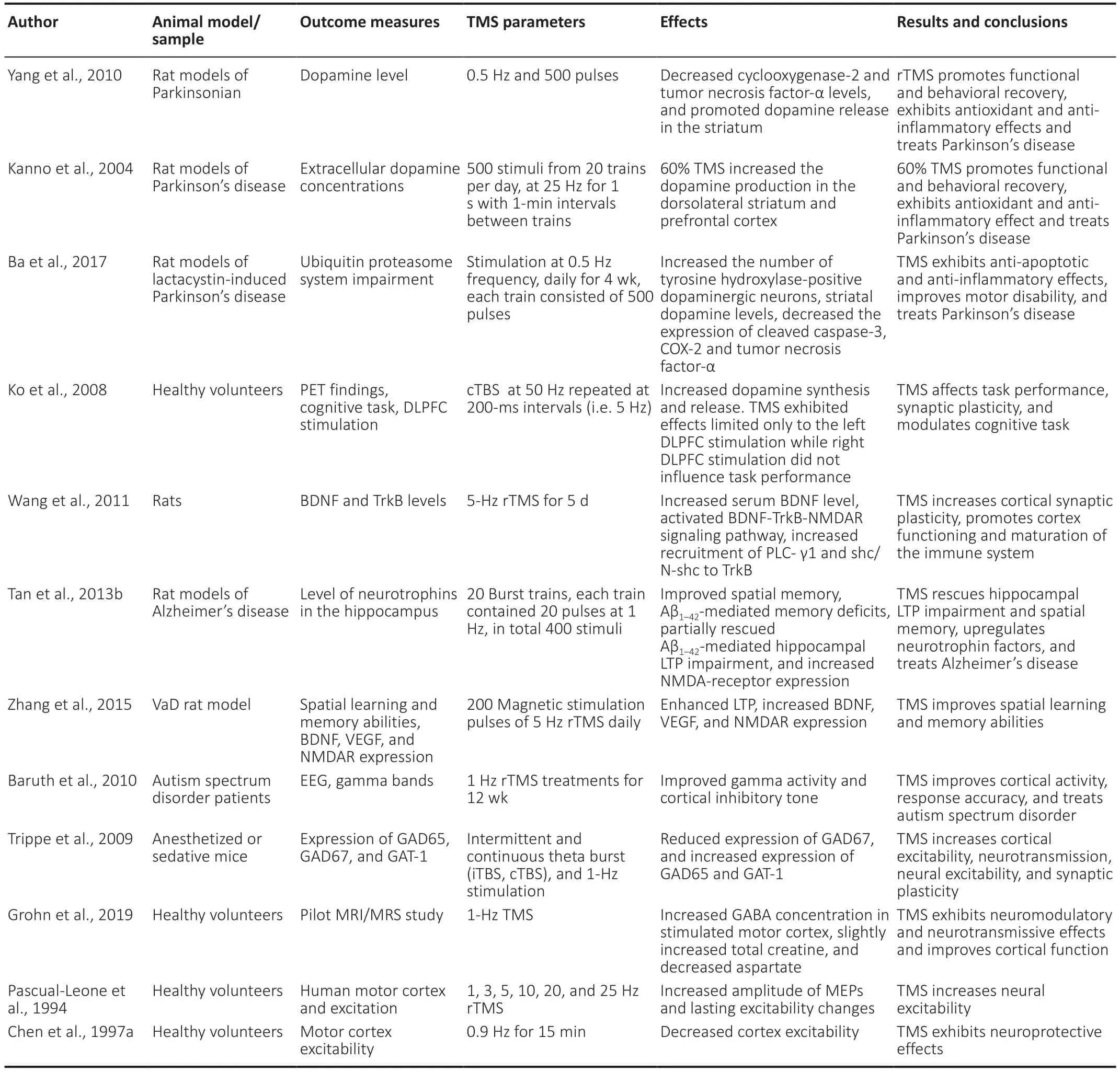
Table 2|TMS regulates the levels of neurotransmitters and neurotrophic factors and exerts neuroprotective effects
TMS also affects the expression levels of several receptors and neurotransmitters. Notably,recent research has shown that rTMS upregulates endogenous neurotrophic content,i.e.,brain-derived neurotrophic factor (BDNF) in the brain (Guo et al.,2017; Levy et al.,2018). Furthermore,BDNF is associated with post-stroke rehabilitation and neuronal protection. Similarly,several studies have shown that the increased expression of BDNF in tissues surrounding the infarction zone during the first week following a stroke is involved in neural protection during acute ischemic damage and neurogenesis (Houlton et al.,2019; Liu et al.,2020; Roslavtceva et al.,2020). The BDNF-Trkb signaling and TrkB-NMDAR interaction,regulated by rTMS,facilitates cortex functioning by increasing cortical excitability (Wang et al.,2011; Zuo et al.,2020). Moreover,low-frequency (1 Hz) rTMS effectively increases hippocampal neurotrophins (NGF and BDNF) and NMDA receptor levels (Tan et al.,2013b). Vascular endothelial growth factors A and B (VEGF-A and VEGF-B) are angiogenesis-related genes with neurotrophic and neuroprotective effects. Studies have found that highfrequency rTMS upregulates VEGF gene-expression,which in turn enhances neurogenesis (Sun et al.,2006; Zhang et al.,2015; Caglayan et al.,2019; Fukuda et al.,2020). Following a stroke,glutaminergic excitotoxicity induces cell death and prevents GABAergic inhibition. Ischemia activates the release of glutamate from neurons,as well as glial cells,inducing acute insult to nerve cells that leads to cell death (Belov Kirdajova et al.,2020). Glutamate is a crucial amino acid in the central nervous system (CNS) that regulates ischemia-induced excitotoxicity (Pinky et al.,2018). Glutamate excitotoxicity may be established after traumatic injury and neurodegenerative disorders,including AD (Belov Kirdajova et al.,2020),PD (Albaugh et al.,2020),and Huntington’s disease (HD) (Crabbé et al.,2019). Glutaminergic excitotoxicity and GABAergic inhibition interaction may affect neuroregenerative processes (Agarwal et al.,2019). Various studies have demonstrated that post-rTMS treatment-induced increases in the level of gamma-aminobutyric acid (GABA) in the medial prefrontal cortex (Trippe et al.,2009; Baruth et al.,2010) and the ipsilateral cortex,accompanied by a decrease in the level of GABA in the contralateral cortex (Gröhn et al.,2019). GABA can stimulate neurite outgrowth,extend the neural network,enhance synaptogenesis,promote neuronal regeneration and differentiation,and increase gene expression (Wu and Sun,2015; Niklison-Chirou et al.,2020). These investigations open a new pathway to understanding the molecular mechanism behind the neurogenesis strategies of TMS by studying neurotrophic factors (such as BDNF and DA) and neurotransmitters (such as GABA and glutamate).
TMS modulates non-neuronal cells (oligodendrocytes,astrocytes,microglia,and adult neural stem cells) and exerts therapeutic effects
TMS therapy is an effective way of diagnosing and treating neurodegenerative diseases,stroke,and ischemia. The insight mechanism of the neuroprotective effect of TMS is still not elucidated clearly. Many pieces of research focused on the beneficial effects of TMS on non-neuronal cells,also known as glia (Yang et al.,2018; Cullen et al.,2019). Glial cells constitute the majority of cells in the adult brain,many folds higher than neurons (Jäkel and Dimou,2017). Neuroglial cells are represented by oligodendrocytes,astrocytes,microglia,and adult neural stem cells (NSCs). These non-neuronal cells do not conduct an electric impulse but play an important role in maintaining tissue homeostasis and providing protection and support to neurons,immune surveillance,and neurotransmission. Central nervous system (CNS) dysfunction or stroke leads to the death of not only neurons but also neuroglia (Jäkel and Dimou,2017). There are few studies on how TMS regulates glial cell activity; however,a small number of studies were done in animal models (Table 3). Neuroglial cells exhibit an important role behind the mechanism of TMS in brain recovery,which has not been greatly explored.
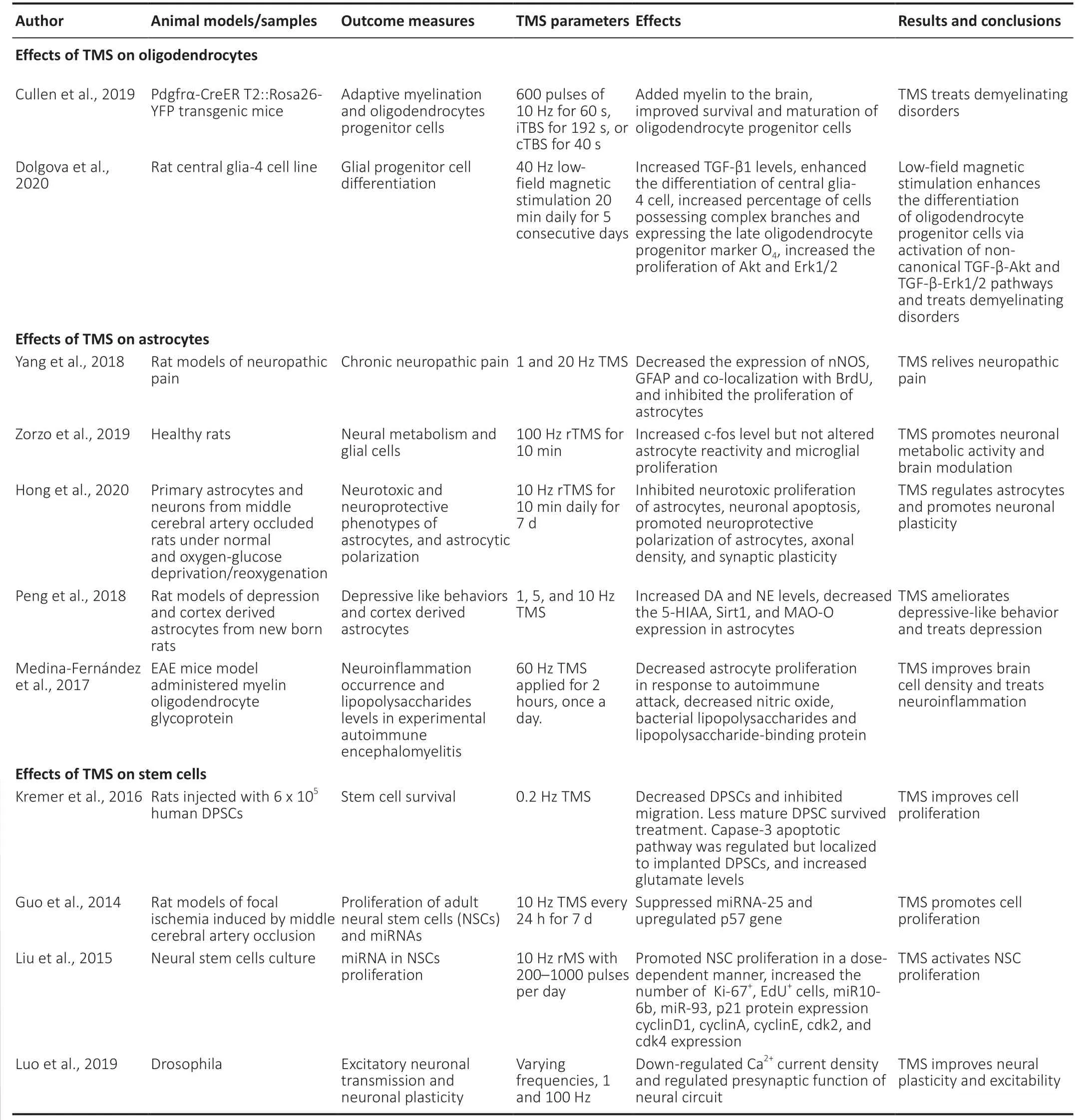
Table 3|TMS modulates non-neuronal cells (oligodendrocytes,astrocytes,microglia,and adult neural stem cell) and exerts therapeutic effects
Effects of TMS on oligodendrocytes
Oligodendrocytes are the highly specialized myelinating neuroglial cells of the central nervous system (CNS). Oligodendrocytes,formed by the proliferation of oligodendrocytes progenitor cells (OPCs),support axons through myelin production,insulation,and optimize action potential conduction (Pepper et al.,2018; Valério-Gomes et al.,2018). They are the susceptible target of injury in a number of neurological disorders and ischemia. As similar to neurons,oligodendrocytes are highly susceptible to damage by oxidative stress,apoptotic pathways,and trophic factors lack (Caprariello et al.,2012; Szebeni et al.,2014). rTMS treatment regulates the neural modulation by modulating GABAergic neurons (Dubin et al.,2016),BDNF (Luo et al.,2017),and VEGF (Zhang et al.,2015). These factors regulate the maturation of oligodendrocytes. Thus,TMS could potentially enhance oligodendrogenesis and myelination (Cullen and Young,2016). Cullen et al. (2019) examined the therapeutic effects of rTMS,delivered as an iTBS pattern,on the oligodendrocytes in a healthy CNS of mice. The results determine that rTMS enhances the number of new premyelinating oligodendrocytes,proliferation,and survival rate of oligodendrocytes. LF-rTMS also enhances myelination,and the increase in myelination cells observed in the cortex region of the adult mice brain (Cullen et al.,2019). Recently,Dolgova et al. (2020) report low-field magnetic stimulation (LFMS) increases the oligodendrocytes progenitor cells differentiation by activating transforming growth factor-beta (TGF-β). In LF-rTMS treated rat central glia-4 (CG4) cell line culture,the elevated level of TGF-β,as well as an increase in the number of cells possessing complex branches and oligodendrocyte progenitor cells observed (Dolgova et al.,2020). The proliferating oligodendrocyte progenitor cells were also found to maintain myelination and remyelination of white matter and ensheathment of axons after SCI. The electrical stimulation of motor cortical neurons results in the increase of oligodendrocyte development and myelin formation in the damaged spinal cord (Li and Li,2017). Thus,the magnetic stimulation can potentially promote oligodendrocyte differentiation and proliferation,which in turn,play an important role in the treatment of CNS disorders and demyelination diseases.
Effects of TMS on astrocytes
Astrocytes are structural,metabolic,and supportive components of neuronal tissues that guide axons,support synapses and regulate the blood brain-barrier. Astrocytes also prevent neurons from oxidative stress,maintain homeostasis at a synapse,and control neuronal signaling (Seifert and Steinhäuser,2013; Allen and Eroglu,2017). Astrocytic glial cells have an important role in neuroinflammation after neurodegenerative conditions,stroke,and ischemia (Cekanaviciute and Buckwalter,2016). Recent researches show that the astrocytes are likely to be an important therapeutic target for neuroprotection,neuro-restoration,and proliferation following stroke and CNS disorders (Barreto et al.,2011; Liu and Chopp,2016). The effects of astrocytes have been well-studied while studying the low- and high-frequency rTMS in mouse models. The rTMS treatment,1 Hz and 20 Hz applied to the rat models of neuropathic pain significantly regulates the expression of astrocytes. The expression levels of neuronal nitric oxide synthase (nNOS),glial acidic fibrillary acidic proteins (GFAP),and the co-localization with 5-bromo-2-deoxyuridine (BrdU) in astrocytes down-regulated after 20 Hz rTMS treatment,as compared with 1 Hz group. These findings suggest that high-frequency (20 Hz) may alleviate neuropathic pain by decreasing the nNOS expression and significantly inhibits the activity and proliferation of astrocytes (Yang et al.,2018). In another study,high-frequency rTMS promotes metabolic and c-Fos activities in the cortex but does not affect astroglial reactivity and microglia reactivity in the hippocampus of healthy rats (Zorzo et al.,2019). rTMS also affects the polarization of astrocytes during cerebral ischemia. Hong et al. (2020) demonstrate that 10 Hz rTMS treatment inhibits neurotoxic astrocyte proliferation and decreases TNF-α,apoptosis,and infarct volume. The anti-inflammatory mediator IL-10 increases after rTMS,thus enhancing neural plasticity and functional recovery following cerebral ischemic stroke in the MCAO rat model subjected to oxygen-glucose deprivation/reoxygenation (Hong et al.,2020). The NIBS techniques have also been reported to exert therapeutic potential against depression,and astrocytes play an important role in this regard. rTMS exerts anti-depressive properties,which depend on different parameters. rTMS treatment has been shown inhibit the over-expression of sirtuin 1 (Sirt1) and MAO-A,in astrocytes,in the prefrontal cortex (PFC) and the cortex-derived astrocytes from the newborn rats (Peng et al.,2018). tDCS activates astrocytes by promoting the intracellular Ca+ signaling that supports synaptic plasticity and amelioration of depression-like behavior (Monai and Hirase,2018). TMS treatment enhances astrocytosis and cell density and can treat neuro-inflammation (Medina-Fernández et al.,2017).
Effects of TMS on microglial cells
Microglial cells are a specialized type of macrophages that remove damaged neurons,protect the injured brain,and mediate immune response in the CNS (Faustino et al.,2011; Zhao et al.,2017). In the absence of microglia,the level of cytokines and chemokines was observed to increase following ischemia,thus increasing infarct volume and profound brain injury (Faustino et al.,2011). Microglia could contribute as a diagnostic marker for neurodegenerative disorders (Hickman et al.,2018; Kwon and Koh,2020). In addition,little is reported on the effects of TMS on microglial cells. However,a study by Zorzo et al. (2019) investigated the effects of 3 days of high-frequency rTMS by examining glial cells in mice. Zorzo et al. (2019) found that high-frequency rTMS did not induce microglial and astrocyte reactivity. In addition,inflammatory response,such as microglial proliferation,was not observed. Moreover,c-Fos expression was increased after TMS treatment (Zorzo et al.,2019). Another study revealed that deep rTMS significantly reduces behavioral anomalies and attenuates myelin breakdown in mouse models. Furthermore,deep rTMS also inhibits microglial expression and activation changes in inflammatory cytokines at the lesioned sites (Yang et al.,2020). Various studies utilize tDCS to elucidate their effects on glial cells. The tDCS treatment effectively induces morphological changes in microglia and shrinks the cell body up to 7.3% in the mouse brain (Mishima et al.,2017). Similarly,Mishima et al. (2019) examined comparable results that tDCS alters the morphology of microglia,but this alteration also depends on adrenergic receptors. tDCS induces the enlargement of microglial soma that is regulated by Ca+signaling and depends upon noradrenaline. Also,the inflammatory response did not appear following tDCS treatment but significantly decreases the microglial surveillance area (Mishima et al.,2019). Moreover,consecutive expression of Iba 1 on microglia in the mouse cortex after the tDCS sessions downregulated in a polarity-dependent manner (Pikhovych et al.,2016). Thus,electrical brain stimulation facilitates stroke rehabilitation via the modulation of microglial regulation and activation.
Effects of TMS on NSCs
NSCs are self-renewing and multipotent cells that have the potential to develop into neurons,astrocytes,oligodendrocytes,or glial cells (Zhao and Moore,2018). Electrical stimulation has been shown to assist NSCs to migrate directionally and differentiate into required cells,which significantly facilitates the treatment of CNS disorders (Huang et al.,2015). Peng et al. (2019) transplanted differentially expressed human NSCs (hNSCs) in an MCAO rat model and treated them with rTMS. The findings show that transplantation of hNSCs followed by rTMS treatment can potentially increase BDNF level,neurogenesis,and functional recovery after ischemic stroke. In another study,Kremer et al. (2016) transplanted human dental pulp stem cells (DPSCs) in the rat cortex followed by rTMS. DPSC survival and migration were found to be decreased or inhibited. In addition,caspase-3 apoptosis pathway and glutamate expression increase in the implanted DPSCs due to rTMS,resulting in reduced survival of DPSCs (Kremer et al. 2016). Caspase-3 expression localized to the implanted DPSC site (Kremer et al.,2016). The rTMS therapy (10 Hz) enhances NSCs proliferation via regulation of micro RNA 25 (miR-25) expression and pathway after focal cerebral ischemia in the subventricular zone (SVZ) of the rat model (Guo et al.,2014). Similarly,it has been found that repetitive magnetic stimulation can potentially increase NSCs proliferation by upregulating miR-106-25 (Liu et al.,2015). Therefore,rTMS combined with stem cell therapy exerts effective therapeutic effects and promotes brain recovery.
TMS affects calcium ions (Ca2+) currents/signaling and promotes neural regeneration and neuroplasticity
The voltage-gated calcium ion (Ca2+) channels play a key role in Ca2+signaling in neuronal cells and are necessary for neurotransmitters release. Ca2+channels regulate neuronal excitability,neurite length,gene expression,and protein levels. The voltage-gated Ca2+channels and Ca2+signaling also have fundamental roles in CNS disorders,such as ischemia,stroke and neuropathic pain (Benarroch,2010). It also has been found that astrocytes and their receptors,including γ-aminobutyric acid (GABA),can mediate Ca2+signaling (Mederos and Perea,2019). After ischemia,Ca2+signaling exerts harmful effects on neurons by enhancing neuronal injury (Ding,2014; Chung et al.,2015). TMS has been reported to regulate Ca2+signaling,thus promoting neuroplasticity and neuroregenerative effects (Table 4). A recent study reported that plasma calcium levels increased significantly bothin vitroandin vivoin humans after rTMS sessions (Stateman et al.,2014). Luo et al. (2019) investigated the effects of rTMS on neuroplasticity,neurotransmission,and recovery after nervous dysfunction by usingDrosophilawhole brain model. Luo et al. (2019) also examined the changes in calcium currents after rTMS at different frequencies (1 Hz and 100 Hz). It has been observed that rTMS regulates calcium current density and activity in a time-dependent manner. The calcium current density was found to be inhibited in the acute group,activated in the 12-hour group,and no changes were observed in the 24-hours group (Luo et al.,2019). The high-frequency rTMS (20 Hz) significantly increases intracellular calcium concentrations in the cortical neurons and may lead to long-term plasticity (Banerjee et al.,2017). These results were also observed by Tan et al. (2013a) that 1 Hz rTMS significantly upregulated Ca2+currents. Moreover,tDCS promotes intracellular Ca2+signaling,supports synaptic plasticity,and ameliorates depression-like behavior (Monai and Hirase,2018). However,after rTMS,Ca2+current density decreased inDrosophilla(Luo et al.,2019). tDCS also promotes the intracellular Ca2+signaling that promotes brain plasticity by forming neurons,astrocytes,and microglial cells. tDCS activates the astrocytic Ca2+signaling and plays an important role in depression treatment (Monai and Hirase,2018). Calcium signaling is essential for pre-synapses,synapses,neural circuit activity,long-term plasticity,and neural plasticity. The electrical stimulation plays an essential role in regulating voltage-gated calcium ion channels and calcium signaling.

Table 4|TMS influences calcium ions current/signaling and promotes neural regeneration and neuroplasticity
Animal Models for Studying TMS
Despite the fact that rTMS has often been implemented in therapeutic clinical trials,its basic mechanisms have not been robustly considered in animal models (Wagner et al.,2007a). Some animal models were used to investigate the mechanism underlying the effects of TMS,but the spatial and temporal dynamics of stimulation patterns remain poorly understood (Boonzaier et al.,2018). As TMS becomes a universally established therapeutic choice,more animal models are needed to examine its long-term safety and adequacy. An ideal animal model would target the behavioral correlates of stimulation in conscious and freely moving subjects.
Animal models for TMS will be helpful for studying the interactions between stimulation-induced changes in neuronal activity,plasticity,and changes in behavioral phenotypes. Most research has focused on the comparative consideration of normal and affected brain functions and the intensification of brain mapping. Animal models could play a potential role in the development of new therapeutic interventions,however,stimulation factors remain yet to be investigated. Although TMS studies involving animal models have distinctive limitations,prospective studies would lead to the provision of more precise data and cannot be discounted.
In 1990,rats were used to study TMS for the first time,after which there was a boom in animal TMS studies. Such studies include animal models of stroke,ischemia (Guo et al.,2017; Boonzaier et al.,2018;Table 1),vascular dementia (Zhang et al.,2018),Alzheimer’s disease (Bagattini et al.,2020),Parkinson’s disease (Ba et al.,2017),and Huntington’s disease (Eddy et al.,2017). Animal models of neurodegenerative disease should fully model the human characteristics of the disease. Although rodents (mice,rats) are considered the most promising animal models for studying the function and behavior of the brain,they are not the only choice for TMS research. One major disadvantage is the ratio of head size to coil size in rodents (Koponen et al.,2020). TMS can stimulate human brains,and the large surface area of the human brain allows currently available coils to differentiate the various areas in the human brain. However,the smallest coils currently available can stimulate the entire rodent brain (Boonzaier et al.,2020). Moreover,the low effectiveness of magnetic stimulation in a rodent brain is another critical drawback,which could make the usefulness of rodent studies debatable (Boonzaier et al.,2020). Although smaller coils have been recently developed to cope with this issue,rodent studies of TMS remain limited in their applicability to the action of TMS in humans. These smaller coils tend to overheat,limiting the number of high-frequency train pulses and reducing field power (Funke,2017). Nonhuman primates also seem to be promising as the large surface area of their brains minimizes focality issues,and they can be trained in many human-like behaviors.
However,animal experimentation also tends to have the following barriers: (1) the cost of maintaining and housing the animals is prohibitive; (2) it can be difficult to train animals to calmly tolerate the loud noises associated with rTMS; and (3) from a technical standpoint,rTMS causes continuous twitching of their jaw muscles (Wagner et al.,2007b; Potashkin et al.,2011). However,cats may represent a good intermediate between rodents and primates for focal stimulation due to their moderate brain size,i.e.,moderate coil ratio,and their amenability to training for experimentation (Wagner et al.,2007a).
Significance of animal models for studying TMS
Various animal studies have been used to investigate the basic molecular mechanism behind TMS effects. As previously mentioned,most of these studies have been performed using rodents,and it is still challenging to transfer findings from these studies to humans. The application of TMS in humans is considerably focal,as the brain surface area in humans is sufficiently large for currently manufactured coils to differentiate the various areas of the human brain. However,TMS cannot be administered focally in rodents due to the small size of their brains as compared to the sizes of currently available coils,and thus the entire brain receives magnetic stimulation (Weissman et al.,1992; Belmaker and Grisaru,1998). According to Weissman et al. (1992),the efficiency of magnetic stimulation is reduced in a small rodent brain,and hence,the sustainability and efficacy of studies using rodents are still uncertain. Stimulation patterns have not been examined for their correspondence to comparative patterns used under clinical conditions (Weissman et al.,1992). When a figure-eight-shaped coil is applied tangentially to the volume conductor,electrical activity significantly increases focally,and a sharp central peak parallel to the long axis is bounded on each side by smaller peaks (Maccabee et al.,1990). The sound of a magnetic stimulator and the effects of TMS on muscles result in a stressful experience for rats. Therefore,there is a need for a sham-controlled group. Extracellular dopamine levels are considerably increased because of stress in the prefrontal cortex (PFC) of the rat brain (Kanno et al.,2004; Cho and Strafella,2009),the environmental exposure,and the handling process (Schwarz et al.,2000). The intensity of stimulus greatly influences extracellular DA levels in the PFC (Feenstra and Botterblom,1996; Volkow et al.,2019).
Transcranial Magnetic Stimulation Applications in Animal Models
Combining animal models of experimental neurological disorders with TMS treatment allows the study and evaluation of the possible mechanism behind the therapeutic properties of TMS,the diagnostics used,and their application in human patients. This information should clarify the feasible utilization of this non-invasive procedure in the treatment of neurodegenerative disorders.
Neurodegenerative disorder models
There are numerous difficulties during generalization of the results from animal models to humans. The factors that must be considered include the animal models used,the type,degree of severity,duration of the disease and the damage induced,the interval of surgical procedures,the size and location of lesions,the ages and sexes of animals,handling procedures such as feeding,maintenance,and the potential effects of pharmacological stress agents. Despite this,animal damage and repair models have been the reason for renowned discoveries in neurological sciences and neurorehabilitation that were not possible through human studies.
Huntington’s disease model
The utilization of electromagnetic forces in neurobiology is usually restricted to investigation of neurodegenerative conditions. However,electromagnetism has been recently applied in neurology as the therapeutic intervention has received more attention,and new techniques and devices have been developed. HD is an autosomal dominant progressive neurodegenerative disorder that occurs because of the dramatic degeneration of the basal ganglia,particularly of GABAergic neurons in the striate nucleus,stimulating particular cognitive,behavioral,and motor degeneration (Naarding et al.,2001). The molecular mechanism of HD is still unknown,and thus no efficient treatment is currently available. However,electrophysiology studies have shown that abnormal MEPs might be associated with altered excitability of the cortico-spinal system as a result of basal ganglia dysfunction (Meyer et al.,1992),while examination using TMS has revealed a decline in cortical excitability before the onset of clinical symptoms. These findings suggest that TMS and other electrophysiological changes might serve as markers of disease progression (Lorenzano et al.,2006; Schippling et al.,2009; Hartmann et al.,2016). Crupi et al. (2008) observed impairment of both cortical and brainstem LTP-like plasticity in symptomatic HD patients,which is similar to the LTP deficits reported in animal models of HD. The studies have shown that continuous TMS (at 60 Hz and 0.7 mT) in a 3-nitropropionic acid (3NP)-induced HD-like rat model prevented the physiopathological changes caused by 3-NP and also reversed these changes following damage (Túnez et al.,2006; Medina and Tunez,2010),while enhancing animal behavior and decreasing apoptosis and oxidative-damage markers. The current study also supported the findings of Brusa et al. (2005) who reported that low-frequency (1 Hz) TMS treatment promoted choreiform movements in HD patients. Likewise,Ihara et al. (2005) examined a decrease in oxidative-damage markers following rTMS in the cerebrospinal fluid of patients with spinocerebellar degeneration. Taken together,these data suggest a possible therapeutic role for TMS. Further research is required in which animal models will be crucial.
Alzheimer’s disease model
AD is a progressive neurodegenerative condition of the central nervous system characterized by irreversible memory loss and cognitive impairment. Even though extensively examined,the mechanism underlying its physiopathology and pathogenesis remains elusive. Notably,basic and clinical studies have demonstrated that altered neuroplasticity,the neurotransmitter system,and synaptic failure are putative common denominators involved in the pathogenesis mechanism. Indeed,synaptic dysfunction is noted in the early stage of AD and has become a therapeutic target for pharmaceutical agents. Mouse models of AD have typically focused on hippocampal long-term potentiation (LTP) impairment as a molecular mechanism associated with synaptic plasticity (Mango et al.,2019). Battaglia et al. (2007) noted that neocortical plasticity impairment was correlated with functional deficits of NMDA glutamate receptors in both AD patients and APP/PSI transgenic mice.
Cappa et al. (2002) reported the activation of the dorsolateral prefrontal cortex following rTMS during verb processing and auditory word recognization,while Cotelli et al. (2006) found that rTMS treatment of the DLPFC accelerated action naming in the control group and enhanced naming performance in AD patients. However,the exact mechanisms involved with improved naming and speech following rTMS are still largely unknown. Cotelli et al. (2008) also demonstrated that rTMS application over the DLPFC improved action naming even in the advanced stages of AD,and proposed that rTMS may enable the brain to recover damaged function during disease development. It has recently been reported that highfrequency rTMS (20 Hz) of the DPLFC improves language performance and object naming in patients with AD (Cotelli et al.,2010). These studies suggest both the beneficial and therapeutic effects of rTMS and its potential usefulness for behavioral and cognitive rehabilitation. However,it should be noted that the induction of long-term changes in cortical excitability depends upon a number of factors,including stimulation intensity and frequency,site of stimulation,and the number of runs. Further research and accurate analysis of these elements are required in order to assess the potential for extrapolating clinical findings in animal models to humans.Future studies on mouse models would be crucial in understanding the mechanisms and effects of TMS on the brain. Although investigators have remained unable to influence specific behaviors,stimulation of the entire brain has facilitated the study of structures that are too deep to stimulate safely in humans. The effects of neuronal excitability and the learning process in memory tasks have been well elaborated using low- and high-frequency rTMS on the hippocampus (Lage et al.,2016). Low-frequency (1 and 8 Hz) rTMS temporarily impaired performance on object recognition tasks,while high-frequency (15 Hz) stimulation improved memory in the test animals (Ahmed and Wieraszko,2006). Moreover,the correlation of these findings with neural mechanisms has been investigated,and it was determined that hippocampal slices from subjects exposed to 15 Hz rTMS showed increased synaptic efficiency and LTP (Ahmed and Wieraszko,2006). The LTP induced by rTMS caused numerous similar neuronal changes,while high-frequency rTMS triggered neural activation,upregulated gene expression (Fujiki and Steward,1997),and varied or altered monoamine regulation (Keck et al.,2002).
Furthermore,mouse research on rTMS and plasticity has also assisted in the understanding of neurodegenerative disorders such as AD. Battaglia et al. (2007) studied the effects of ppTMS on LTP in a mouse model of AD and described the mechanisms of impaired plasticity. In addition,the determined deficits were correlated with and mirrored by TMS studies in patients with AD. Liu et al. (2019) reported that N-methyl-Daspartate receptor (NMDAR) -related dysfunction could alter plasticity. Coherent elaboration of the pathophysiology of AD would facilitate the development of novel potential drugs for modulating NMDAR function to improve plasticity,learning,and memory in AD patients.
Parkinson’s disease
PD is the second most common neurologic disorder after AD and the most complex progressive movement disorder of aging,i.e.,it mainly affects the older adult population (Váradi,2020). Loss of dopamine in the specific area of the brain called the substantia nigra due to abnormal protein aggregation is the underlying mechanism of the pathogenesis of PD (Tufail,2019). The hallmark clinical indicators are resting tremor,rigidity,and bradykinesia.
Pooled evidence suggests that TMS may be useful as a therapeutic intervention for PD. Pascual-Leone and colleagues were the first to report that high-frequency rTMS (5 Hz) facilitated contralateral hand function in normal subjects (Pascual-Leone et al.,1994; Fregni and Pascual-Leone,2007). Multiple studies and meta-analyses (Fregni et al.,2005; Elahi et al.,2009) have also confirmed the modest but significant effects of TMS on PD motor symptoms. Although most studies have focused on the favorable effects of high-frequency stimulation (Siebner et al.,2000; Fregni and Pascual-Leone,2007; Hamada et al.,2008; Elahi et al.,2009; Lefaucheur,2009; Sedláčková et al.,2009),a recent study has also revealed the favorable impact of low-frequency (1 Hz) rTMS in improving motor function (Filipović et al.,2010).
Despite the enormous potential of TMS for PD therapy,the mechanisms underlying its beneficial effects are yet unexplored. Animal models are crucial to achieving an understanding of these mechanisms. It is well established that degeneration of dopaminergic neurons in the striate nucleus plays an important role in the pathogenesis of PD and replacement of these neurons alleviates PD symptoms (Zhai et al.,2019). Several animal studies have already highlighted the effects of prefrontal cortex stimulation on dopamine levels in the striate nucleus (Karreman and Moghaddam,1996). Kanno et al. (2003) revealed that continuous application of high-frequency (25 Hz) rTMS for 3 days in rats resulted in an improvement of motor symptoms by enhancing dopamine release in the dorsolateral striatum (Kanno et al.,2003). Keck et al. (2002) also examined the influence of highfrequency (20 Hz) rTMS on the hippocampal,striatal,and reward-related accumbal dopamine levels and associated metabolites (homovanillic acid and 3,4-dihydroxyphenylacetic acid) in mouse models. The findings provided the firstin vitroevidence of the positive modulatory effects of acute rTMS on both mesostriatal and mesolimbic dopaminergic neurons when employed in the frontal brain region. The increase in dopamine neurotransmission could potentially explain the preliminary findings of the beneficial effects of rTMS on PD. Yang et al. (2010) highlighted the neuroprotective effect of rTMS on the degeneration of dopaminergic neurons in the substantia nigra of a 6-hydroxydopamine-induced rat model of PD. In addition,cyclooxygenase-2 (COX-2) and tumor necrosis factor-alpha (TNF-α) were found to be lower following rTMS in animal models. Yang et al. (2010) (Table 2) also highlighted the positive effects of decreased COX-2 and TNF-α induced by low-frequency rTMS in PD patients including increased levels of dopamine,antioxidants,and anti-inflammatory activity. These multiple animal findings have been endorsed in humans,confirming increased striatal dopamine release following rTMS to the DLPFC (Cho and Strafella,2009). All these related studies linked their etiological findings with mitochondrial changes through oxidative stress,as well as mitochondrial membrane potential impairment due to the pathogenesis of PD (Deas et al.,2011). The therapeutic role of TMS in improving oxidative balance and inflammation as reported by Yang et al. (2010) has also been discursively verified by other researchers (Túnez et al.,2006; Velioglu et al.,2021).
TMS can induce neurogenesis and recovery action of lost neural populations. Following nigrostriatal denervation,4 hours of daily stimulation for 60 days significantly increased bromodeoxyuridine-positive (BrdU+) cells localized in the subventricular zone. These newly generated proliferating neurons stained positive for tyrosine hydroxylase,i.e.,they produced dopamine. In the same subjects,TMS protected against lesion-induced motor impairment (Arias-Carrión et al.,2004).
The overall findings indicate the significant importance of animal models of PD in exploring the therapeutic potential of TMS in PD patients and clarifying the mechanisms of both the remedial pathways mediated by TMS and the pathophysiology of the disease itself.
Depression
NIBS is used to modulate brain networks and induce changes in cortical excitability by means of weak electric currents or magnetic pulses. TMS,a non-invasive brain stimulation technique,uses magnetic fields to stimulate focal cortical brain regions and neural tissues underlying the cerebral cortex (Barker et al.,1985). Due to its non-invasiveness and painless stimulation,TMS has become a valuable tool in neurosciences. One of the most clinically considered applications of TMS is its potential in treating the major depression. In 2008,the US Department of Health and Human Services,Food and Drug Administration (FDA) approved and permited the marketing of TMS as a treatment for major depression. Thus,TMS is a recognized and widely accepted treatment for patients having major depressive disorder,depression-like disorders,or treatment-resistant depression (Dubin et al.,2016). The incredible potential of TMS treatment has been reported by a number of studies using animal models to examine the insight mechanism for TMS novel therapeutic approaches. However,although various researches examine the therapeutic effects of TMS on depression,the pattern of activation,the underlying mechanism behind the contribution of TMS on altering mood,and stimulation parameters (frequency,intensity,duration,and number of pulses) remain incompletely understood. rTMS exerts antidepressant effects that depend upon the frequency and intensity of stimulation applied,that is evident by animal models (Table 5). The low-frequency rTMS (1 Hz) inhibits subsequent cortical responses (Chen et al.,1997a),whereas high-frequency rTMS (5 and 10 Hz) exhibits neural facilitation (Pascual-Leone et al.,1994).
In a forced swim test (FST) model of depression,rTMS significantly decreases immobility time with a range of frequencies from 1 to 25 Hz,suggesting an anti-depressant effect. The 25-Hz frequency rTMS works more efficiently than 5- and 15-Hz frequency rTMS (Sachdev et al.,2002). Similar to studies regarding the effects of TMS on stroke and neurodegenerative disorders,TMS treats depression through affecting neurobiological levels,molecular level,and neurogenesis. It has been observed from rat models that rTMS evokes different patterns of immediate-early gene expression that are effective for treating depression. rTMS activates the neural responses of the paraventricular nucleus of the thalamus (PVT) (Ji et al.,1998). High-frequency (15 Hz) rTMS significantly increases neurogenesis,and cell proliferation in the chronic unpredictable mild stress (CUMS) exposure produces representative depression-like behavior,suppresses the proliferation of hippocampus,and increases immobility time in FST. Moreover,15-Hz rTMS reduces anhedonic-like behaviors and immobility time in FST. Furthermore,these changes were sustained for 2 weeks after the cessation of rTMS treatment (Feng et al.,2012). Wang et al. (2014) and Zhao et al. (2018) observed comparable results. The findings suggest that rTMS administration to CUMS-exposed rat models reverses and reduces depressivelike behaviors and immobility time in FST and upregulates cell proliferation. The CB1 receptor,BDNF protein levels,and Bcl-2/Bax enhance after rTMS (Wang et al.,2014). rTMS also restores the balance of hypothalamic-pituitary-adrenocortical (HPA) axis,reduces adrenocorticotropic hormone (ACTH) and cortisol (CORT) plasma levels in a rat model of chronic unpredictable mild stress-exposed depression (Zhao et al.,2018). 10-Hz rTMS recovers cortical neural excitability,calcium-activated potassium (BK) channels,and thus reduces the depression-like behaviors in the mouse model of depression in the forced swimming test. Sun et al. (2011) reported that Homer1a gene expression was down-regulated after forced swimming but this was recovered by 10-Hz rTMS. Zuo et al. (2020) investigated the effects of 15-Hz versus 25-Hz high-frequency rTMS in a mouse model of CUMS-induced depression. HF-rTMS ameliorates depression-like and anxietyrelated behaviors,as well as inhibits CUMS-induced neuronal loss and apoptosis. At both frequencies (15 Hz and 25 Hz),rTMS exerts antidepressant effects,and increase BDNF,Homer1a,p11,and p-trkB proteins level. In contrast,15-Hz rTMS inhibits CUMS-induced neuronal loss and apoptosis,whereas 25-Hz HF-rTMS promotes synaptic plasticity (Zuo et al.,2020). On the other hand,low-frequency pulsed magnetic fields (1 Hz) reduce depressive-like behavior and cognitive impairment in a mouse model of chronic unpredicted stressinduced depression. LF-rTMS influences the reversal memory of rats,prevents spatial learning and memory deficits (Yang et al.,2019). In another study (Moshe et al.,2016),subconvulsive electrical stimulation,a different type of brain stimulation,was used to examine its effects on depressivelike behaviors and BDNF levels. The authors treated the prelimbic cortex of depressed rat line model with repeated sub-convulsive electrical stimulations for treatment of major depressive disorder and drug-resistant major depressive disorder. The repeated sub-convulsive electrical stimulations were found to ameliorate depressive-like behaviors and increase BDNF protein expression in the hippocampus and prefrontal cortex of a hereditary rat model of drug-resistant major depressive disorder (Moshe et al.,2016). The increase in BDNF protein expression level was also confirmed by Dubin et al. (2016). The NIBS technique,in particular TMS,has potential anti-depressive properties,mainly by regulating apoptosis and BDNF and other protein levels. TMS was used to treat depression and similar disorders through regulating neurogenesis,synaptic plasticity,cell proliferation,and neural facilitation (Table 5). TMS remains the best choice for preclinical and clinical settings of depression treatment by electrical stimulation.
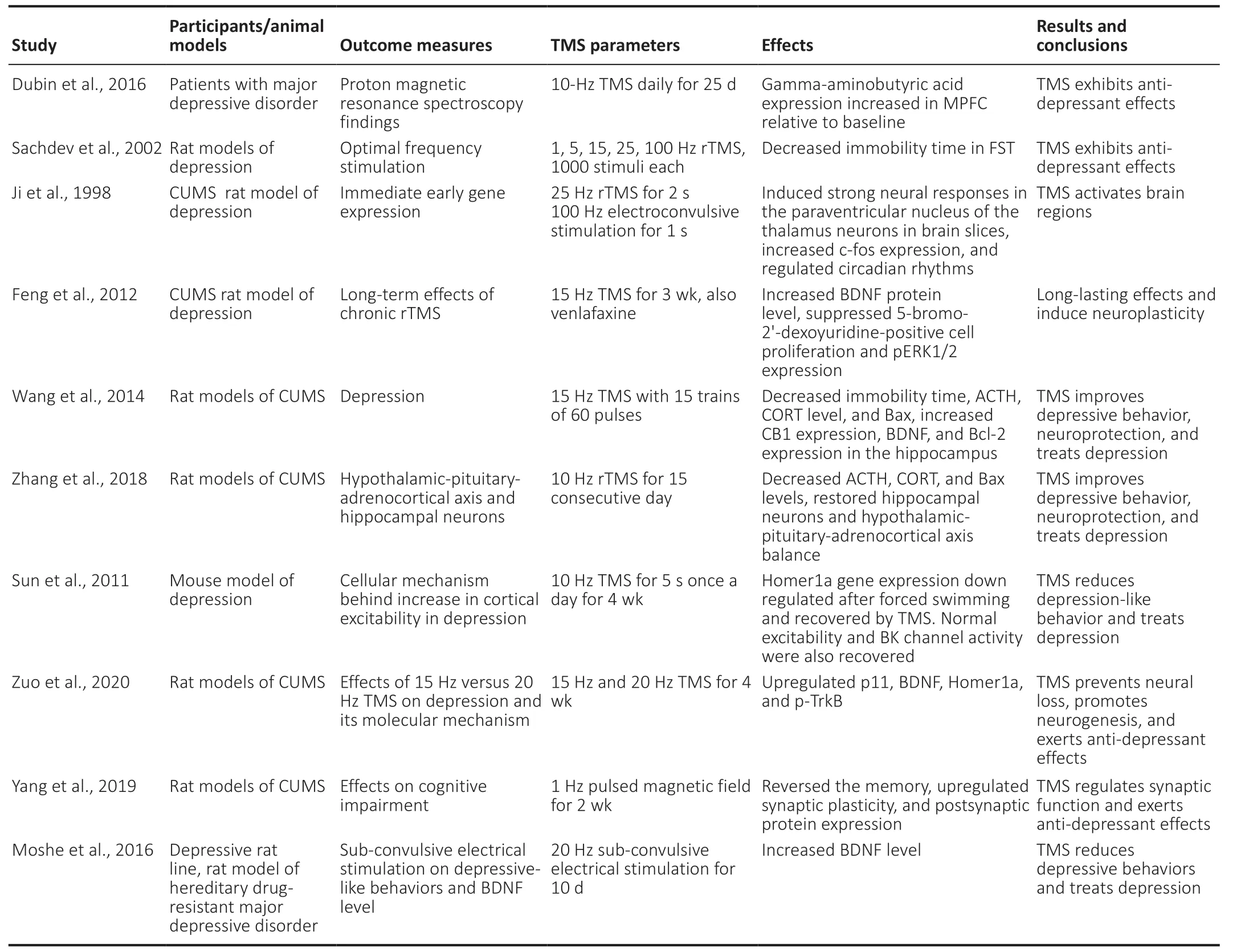
Table 5|TMS exerts therapeutic effects on depression
Potential Side Effects of Transcranial Magnetic Stimulation in Humans
Although TMS,first successfully demonstrated in 1985,is a non-invasive neuromodulation technique that stimulates the brain,its potential direct,long-term,or short-term side effects of magnetic stimulation must be considered. TMS is non-systemic; that is,any side effects are restricted to the treatment area and are not spread to the other parts of the body. The current safety precautions and practice recommendations have been outlined by Wassermann (1998). The safety guidelines for TMS describe the limits for frequency combinations,current intensity,and trains of stimuli length that should be considered to prevent most problems and side effects (Wassermann,1998). Caution is necessary since when the high voltage current passes through the coil,subjects may experience tingling in the scalp and tension headaches (Bolognini and Ro,2010). rTMS may induce epileptic seizures if it is applied at higher frequencies and intensities. These severe and critical side effects depend upon the stimulation parameters and the stimulation frequency (Sack and Linden,2003). Seizures are rare after sTMS,ppTMS,and lowfrequency rTMS. However,they may appear after highfrequency rTMS. This may be avoided by utilizing specified stimulation frequencies and a combination of parameters,as reported by Chen et al. (1997b) and Wassermann (1998). TMS can cause a disturbance in the cognitive domain that remains beyond the stimulation period itself. TMS may also generate required,unrequired,and potentially long-lasting changes to the subject area. In the context of safety,parameter combinations with short trains and long inter-train intervals carry a low risk (Rossi et al.,2009),and the intensity of TMS is a more important parameter than the frequency. Individual cortical excitability should be considered for stimulation intensity (Sack and Linden,2003).
Discussion
TMS is a non-invasive neuromodulation procedure with relatively few adverse effects. It is most frequently used to study brain stimulation. TMS has been used extensively due to its non-invasiveness,painlessness,effectiveness,and safety. TMS significantly stimulates the brain and extensively treats stroke,ischemia,and neurodegenerative disorders. The consecutive pulses of stimulation applied with variable intervals to various brain areas or trains of repetitive stimuli at various frequencies can modulate brain activity by regulating several neurotransmitters,neurotrophic factors,cellular signaling,and cellular pathways (Tasset et al.,2012; Guo et al.,2017). TMS likely ameliorates CNS disorders by increasing BDNF,NMDA,GABA and extracellular dopamine levels,and Bcl-xL protein expression,while reducing levels of Bax and other proteins in the caspase family to inhibit apoptotic pathways. Studies have confirmed that rTMS potentially regulates neural activity,such as neurogenesis,neuronal regeneration,and neuronal excitability (Gao et al.,2010,2017; Jin et al.,2018). However,the cellular and molecular basis of the effects exerted by TMS is still unclear.
TMS is still of great interest for research in neurological sciences because of its therapeutic effects. Animal models are very advantageous for studying the therapeutic effects of TMS and the mechanism behind its brain-stimulating activity. Thus,suitable animal models are required to assess the effects of TMS as well as its efficiency and safety. Animal models,most commonly using rats and mice,significantly contribute to studying new therapeutic interventions and TMS stimulation parameters that are yet to be investigated. Hence,animal models play a pivotal role in the study of the influence of TMS on brain activity and provide an easy way to investigate its effects at the molecular level. Various studies of neurodegenerative disorders and stroke have been performed using animal models to investigate the therapeutic effects and changes at the molecular level induced by TMS (Battaglia et al.,2007; Ba et al.,2017; Boonzaier et al.,2018).
Animal models are extensively used to investigate the therapeutic effect of TMS on neurodegenerative conditions,including AD,HD,PD,and depression. TMS could be a potential therapeutic tool to modulate neuronal function at the molecular and cellular levels,improve neuronal activity,excitability,plasticity,and functional recovery. In many neurodegenerative conditions,such as stroke,epilepsy,depression,AD,HD,and PD,a significant increase in neuronal cell death was observed due to apoptosis. Studies invovling animal models have found that TMS inhibits the apoptotic pathway by decreasing apoptosis-promoting proteins (Bax,Bad,and Bcl-xS) while increasing anti-apoptotic proteins such as Bcl-2 and Bcl-xL (Fujiki et al.,2003; Yoon et al.,2011; Kale et al.,2018; Pemberton et al.,2021). TMS also reduces caspase-3 protein expression that triggers apoptosis. In addition,blocking caspase-3 expression reduces ischemic area,infarct volume,and lesion size by seizing apoptosis (Namura et al.,1998; Ferrer et al.,2003). TMS arrests the apoptosis pathway and promotes neurogenesis and neuronal regeneration,which may be the basic mechanism behind the neuroprotective effects of TMS on the brain.
TMS potentially treats different neurodegenerative disorders by stimulating various neurotransmitters and neurotrophic levels in the brain,thereby improving neuronal activity. Studies have demonstrated that TMS treatment increases BDNF expression that regulates synaptic plasticity,cortical excitability,and neurogenesis (Roslavtceva et al.,2020). BDNF promotes nerve regeneration,plasticity,and synaptic formation via tyrosine kinase receptor B (Trk-B). Following rTMS treatment,BDNF-TrkB signaling increases,which in turn stimulates BDNF-TrkB-NMDA interaction and improves cortex functioning by increasing cortical excitability (Wang et al.,2011). The rTMS treatment also ameliorates gammaaminobutyric acid expression in neurons (Baruth et al.,2010). Additionally,TMS regulates dopamine synthesis and release. TMS promotes the survival of dopaminergic cells,thus increasing extracellular dopamine levels and significantly improving motor function (Yang et al.,2010; Ba et al.,2017). Moreover,TMS also increases intracellular Ca2+concentration and activates Ca2+channel (Tan et al.,2013a; Banerjee et al.,2017). Hence,TMS may exert neuroprotective effects by considerably regulating neurotransmitters and neurotrophic factors and promoting neurogenesis and neuromodulation.
Animal models of TMS and NIBS are the sine qua non to fully understand the impact of these techniques,optimize safety parameters,and improve clinical effectiveness. Future research should be performed to pursue results in awake animals (such as cats or nonhuman primates) and combine neuroimaging techniques (fMRI) with electrode recording to better assess these techniques for validated implementations in clinical therapy. Furthermore,a combination of TMS and neuroimaging techniques is required for the effective study of the brain and neuronal activities and to establish specific TMS treatment protocols. Specifying parameters for TMS will be necessary to optimize TMS interventions in the near future for the efficient treatment of neurological disorders.
This review has shown that:
i) Various animal models exist for the testing and development of TMS protocols.
ii) Animal models used for TMS provide insights into human diseases that TMS proposes to alleviate.
iii) Knowledge of the effects of TMS on neurotransmission and how TMS interacts with disease therapy comes from animal studies.
Conclusions
The main goal of this review was to summarize the basic understanding of TMS in brain stimulation and neurodegenerative disorders gained from animal models. In addition,we emphasized the effects of TMS and rTMS on biological parameters studied using animal models. Although there are few studies on TMS brain stimulation in animal studies,the therapeutic and neurorehabilitative effects of TMS are promising and open a pathway for researchers to further explore its molecular mechanism. rTMS has positive effects on functional recovery through neural regeneration,neurogenesis,and neuroprotection. Moreover,negative or null results should also be reported to fully understand the effectiveness of TMS. Furthermore,methodological limitations were discussed throughout this review to highlight the need for improvements in the reporting of mechanisms and practices associated with TMS.
Author contributions:All authors participated in design,writing and review of the manuscript and approved the final version of the manuscript.
Conflicts of interest:The authors declare that they have no conflicts of interest.
Financial support:None.
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal,and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License,which allows others to remix,tweak,and build upon the work non-commercially,as long as appropriate credit is given and the new creations are licensed under the identical terms.
Corrigendum
Corrigendum: Offspring of rats with cerebral hypoxia-ischemia manifest cognitive dysfunction in learning and memory abilities
https://doi.org/10.4103/1673-5374.314115
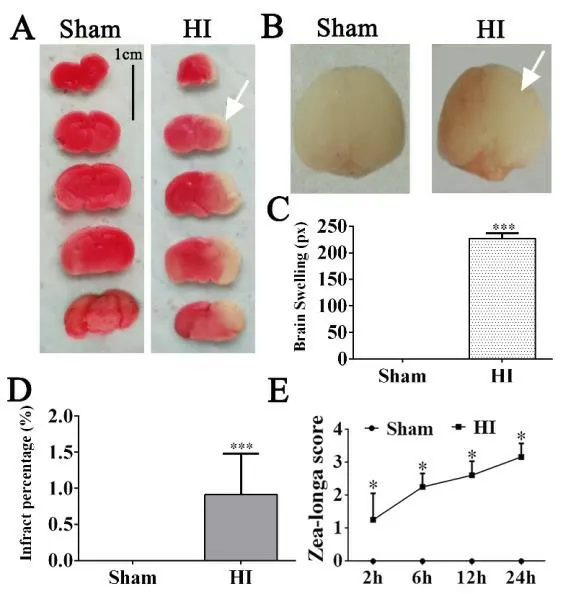
In “Offspring of rats with cerebral hypoxiaischemia manifest cognitive dysfunction in learning and memory abilities”,which was published on pages 1662-1670,Issue 9,Volume 15 ofNeural Regeneration Research(Xue et al.,2020),Figure 1A appears incorrectly because of the author’s error made in image selection.
The correct Figure 1 is shown as below:
In addition,a grant was lost in the financial support section; the correct financial support section is shown as below:
This study was supported by Research Fund for the Doctoral Program,No. 201903 (to LLX); the National Natural Science Foundation of China,No. 81560215 (to FW); and the Innovative Research Team Program of Science and Technology in Yunnan Province of China,No. 2017HC007. The funding sources had no role in study conception and design,data analysis or interpretation,paper writing or deciding to submit this paper for publication.
The online version of the original article can be found under doi: 10.4103/1673-5374.276359.
The authors apologize for any inconvenience this corrigendum may cause for readers and editors ofNeural Regeneration Research.
Reference
Xue LL,Wang F,Niu RZ,Tan YX,Liu J,Jin Y,Ma Z,Zhang ZB,Jiang Y,Chen L,Xia QJ,Chen JJ,Wang TH,Xiong LL (2020) Offspring of rats with cerebral hypoxia-ischemia manifest cognitive dysfunction in learning and memory abilities. Neural Regen Res 15:1662-1670.
杂志排行

中国神经再生研究(英文版)的其它文章
- Deciphering the role of PGC-1α in neurological disorders: from mitochondrial dysfunction to synaptic failure
- Dying by fire: noncanonical functions of autophagy proteins in neuroinflammation and neurodegeneration
- SYNGR4 and PLEKHB1 deregulation in motor neurons of amyotrophic lateral sclerosis models: potential contributions to pathobiology
- Cholesterol synthesis inhibition or depletion in axon regeneration
- Challenges in developing therapeutic strategies for mild neonatal encephalopathy
- Delving into the recent advancements of spinal cord injury treatment: a review of recent progress