中华鳖饲料复合植物蛋白替代鱼粉的效果研究
2022-08-06朱站英王刚黄仲园华雪铭
朱站英,王刚,黄仲园,华雪铭,3,4
(1.湖州海皇生物科技有限公司,浙江湖州 313105;2.上海海洋大学,农业农村部鱼类营养与环境生态研究中心,上海 201306;3.上海海洋大学,农业农村部淡水水产种质资源重点实验室,上海 201306;4.上海海洋大学,水产科学国家级实验教学示范中心,上海 201306)
为降低中华鳖饲料生产成本,众多学者对鱼粉替代都做了大量研究。有研究表明,在饲料中使用国产鱼粉不会降低石斑鱼和黄颡鱼的生长性能,但国产鱼粉因含有较高的组胺而影响其新鲜度,并因此降低虹鳟的摄食率(Cowey,1992)。鉴于此,鱼粉和优质植物蛋白源的合理搭配成为实现饲料配方优化的又一有效途径。在优质蛋白源的研究中,将某些植物蛋白进行发酵,降解为易于吸收的小分子蛋白和小肽,并降低其抗营养因子,提高蛋白的吸收利用率,已成为另外一个研究热点。发酵豆粕是优于去皮豆粕的鱼粉替代物,已有研究表明,当饲料中发酵豆粕的用量不超过10%时,并不会显著性影响石斑鱼、鳗鱼、中华鳖和加州鲈的特定生长率、饲料效率和蛋白质效率(Zou等,2012)。
大米蛋白粉因其氨基酸平衡性与秘鲁鱼粉相似,已成为新型蛋白质替代源。Palmegiano 等(2007)对黑鲷的研究发现,大米蛋白粉替代20%鱼粉时取得了很好的效果,但当替代量超过20%时,会显著降低其生长性能。复合蛋白质替代源因其氨基酸互补效应和经济效益显著,往往是配方调整策略的首选。已有研究表明,含有大米蛋白粉的复合蛋白质源最高可以替代80%的鱼粉,而不显著影响凡纳滨对虾的生长、成活和饲料效率(刘尧等,2014)。本试验旨在丰富中华鳖饲料原料来源,调整配方结构,通过改变大米蛋白粉、发酵豆粕和去皮豆粕的用量,探讨复合植物蛋白源替代鱼粉在中华鳖饲料上的可行性,为降低中华鳖饲料生产成本、找到一种性价比最优的配方提供依据。
1 材料与方法
1.1 养殖试验
1.1.1 试验设计 根据饲料配方中复合植物蛋白源与鱼粉使用量比值(28%、28%、33%与41%)共设4组,分为对照组(生产配方P/F28C),试验组P/F28E、P/F33E 和P/F41E,在P/F28C 组的基础上不使用进口鱼粉1,提高进口鱼粉2 和发酵豆粕的使用量,并辅以降低国产鱼粉2 使用量得到试验组P/F28E 配方;同步提高进口鱼粉2、大米蛋白粉和发酵豆粕的使用量,并辅以进一步降低国产鱼粉2 的使用量来降低进口鱼粉1 的使用量,得试验组P/F33E 配方;在试验组P/F33E 的基础上进一步升高大米蛋白粉和进口鱼粉2 使用量来降低进口鱼粉1 的使用量,得试验组P/F41E 配方。配方设计的总体思路是在降低高蛋白含量进口鱼粉的同时,适当增加低蛋白含量进口鱼粉和优质植物蛋白源(大米蛋白和发酵豆粕)的用量,同时降低低蛋白含量国产鱼粉和去皮豆粕的用量。每组4 个重复,每重复1000 只中华鳖(台湾品系)。试验用中华鳖购自浙江省杭州市萧山区中华鳖养殖场,鳖体重(6.93±0.28)g。饲料组成及营养水平见表1。
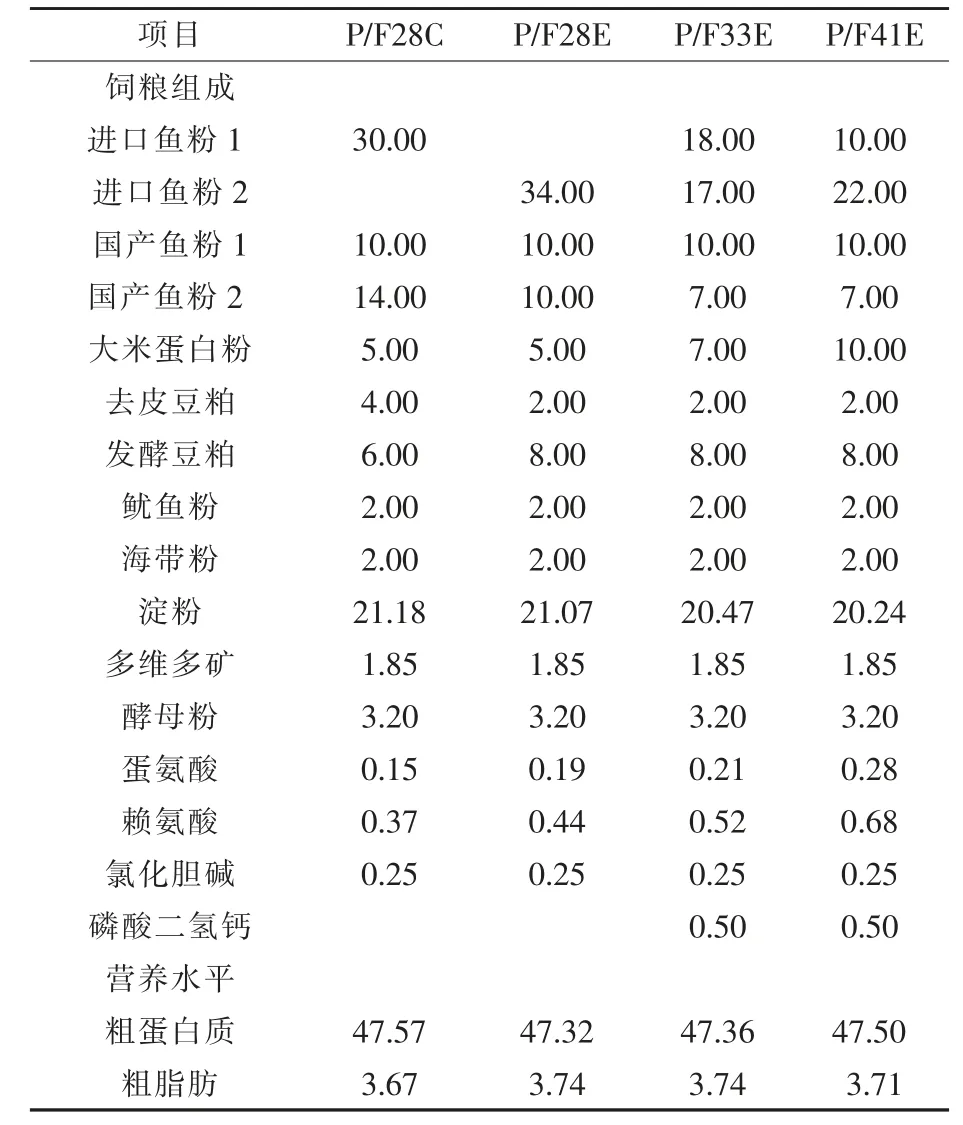
表1 饲料组成及营养水平(风干基础) %
1.1.2 养殖条件及日常管理 养殖试验在杭州海皇科技股份有限公司的养殖场温棚中进行。将体质健壮的16000 只稚鳖随机分到16 个水泥池,养殖池面积20 m2,水深50 cm,池中有增氧设备,棚中有加热锅炉,黑暗养殖。水温保持在(31±1)℃。
对试验鳖进行2 周驯化,用商品稚甲饲料饱食投喂,每天3次,时间为8:00、13:00、18:00,直至正常摄食。禁食24 h,擦干鳖体,称重作为平均初始体重(W0)。试验期为49 d,于试验末期对中华鳖进行一次抽样称重,记录为平均终重(W1),并统计成活率。所用粉状饲料在每餐投喂时加其重量2%的油脂和50%的地下水混合均匀,用软颗粒机制成软颗粒后供投喂使用,每天分别于8:00、13:00、18:00 饱食投喂,并记录投喂量。
成活率(SR)/%=St/S0×100;
式中:St为生长试验末期中华鳖成活数;S0为生长试验开始时的中华鳖总数。
1.2 抗应激试验(冬眠)继生长试验后(11 月份),拆除温棚,于原水泥池进行中华鳖有沙冬眠,期间停止增氧和喂食。以生长试验末期作为冬眠初期,记体重W1,之后,分别于次年2 月(冬眠中期)和4 月(冬眠末期)采样,分别记体重W2、W3,试验为期6 个月。
相对增重率(WGR)/%=(Wt-Wt-1)/Wt-1×100;
式中:Wt为试验第t 期时中华鳖体重,t=1、2、3,分别对应冬眠初期、中期和后期;Wt-1为试验第(t-1)期时中华鳖体重。
1.3 样品分析
1.3.1 形体指数 在养殖末期(即冬眠初期)、冬眠中期和冬眠末期分别量取每组中华鳖的体长,分别记为L1、L2和L3;称取每组中华鳖肝脏重量,分别记为WH1、WH2和WH3。
肥满度/(g/cm3)=Wt/Lt3;
式中:Wt为试验第t 期时中华鳖体重,t=1、2、3;Lt为试验第t 期时中华鳖体长,t=1、2、3。
肝体比/%=WHt/Wt×100;
式中:WHt为第t 期肝脏重量,t=1、2、3。
1.3.2 体成分测定 冬眠期间,分离中华鳖鳖体和内脏,于60 ℃恒温烘干,制得风干样,测定其粗蛋白质、粗脂肪和粗灰分,测定方法分别参考凯氏定氮法(GB5009.5-2010)、索氏抽提法(GB/T5009.6-2003)和高温灼烧法(GB5009.3-2010)。
1.3.3 血清非特异性免疫相关酶 冬眠期间,每组取10 只中华鳖,断头取血,离心得血清,于-20℃冷藏,以检测血清中碱性磷酸酶、溶菌酶和TSOD,检测方法参考自南京建成试剂盒说明书。
1.3.4 组织消化酶 冬眠期间,每组取3 只中华鳖,取出肝脏、肠道和胃于-20 ℃冷藏,以检测组织中的淀粉酶和蛋白酶,淀粉酶检测方法参考南京建成试剂盒说明书,蛋白质酶检测方法为福林酚法。
1.4 数据处理及统计分析 结果用“平均数±标准差”表示,并用SPSS 17.0 软件进行单因素方差分析(Anova)。当差异显著时(P <0.05),进行Duncan's 多重比较。
2 试验结果
2.1 生长性能 由表2 可知,养殖末期各组中华鳖的相对增重率、肥满度、肝体比、饲料系数和成活率均无显著性差异(P >0.05),但P/F28E增重率稍显弱势。由表3 可知,在冬眠的抗应激试验中,所有试验组的增重率均表现为先下降后上升,但试验组P/F33E 能在冬眠复苏后快速适应环境而增加体重,相对增重率较对照组提高1.04%(P >0.05),试验组P/F28E 和P/F41E相对增重率较对照组分别降低13.02% 和25.96%(P <0.05)。冬眠后中华鳖肥满度为先下降后上升,肝体比则持续下降。
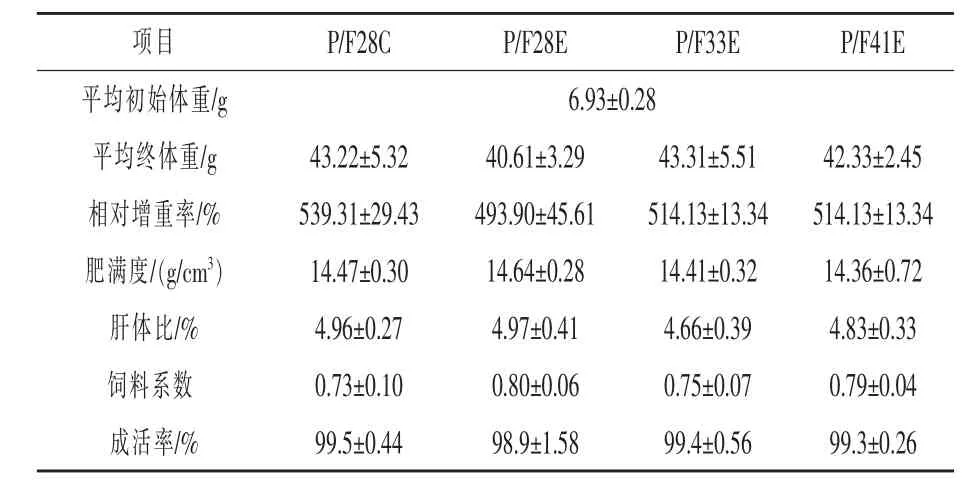
表2 不同配合饲料对中华鳖生长和形体指数的影响
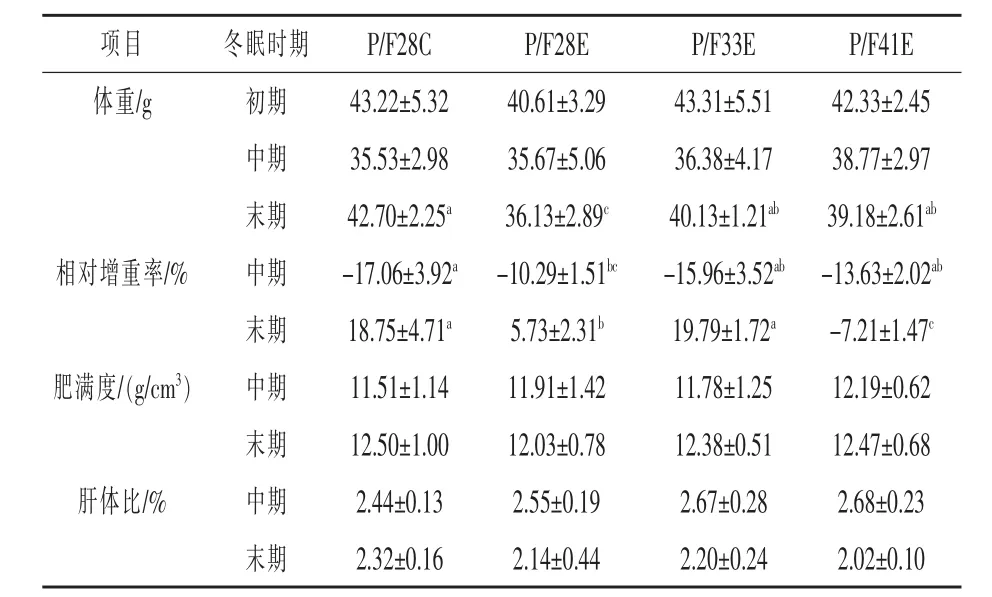
表3 不同配合饲料对冬眠期间中华鳖生长和形体指数的影响
2.2 体成分分析
2.2.1 冬眠中华鳖全鳖常规营养成分分析 由图1 可知,中华鳖全鳖粗蛋白质含量随冬眠时间的延长显著性上升(P <0.05),粗脂肪含量显著性下降(P <0.05)(图2);P/F28C 中华鳖在冬眠末期时,较冬眠中期全鳖粗蛋白质含量有所下降,粗脂肪含量有所上升,但均无显著性差异(P >0.05)。试验各组粗灰分含量在冬眠期间均显著性上升(P <0.05),且在冬眠初期各组之间无显著性差异(P >0.05),冬眠中期P/F28E 组低于其他试验组,冬眠末期P/F41E 组显著高于其他试验组(P <0.05)(图3)。
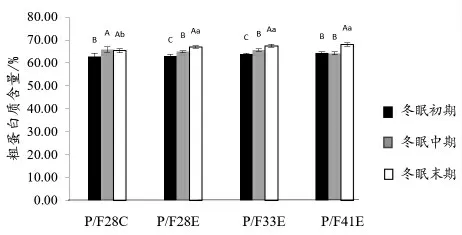
图1 中华鳖在冬眠不同时期全鳖粗蛋白质含量比较

图2 中华鳖在冬眠不同时期全鳖粗脂肪含量比较
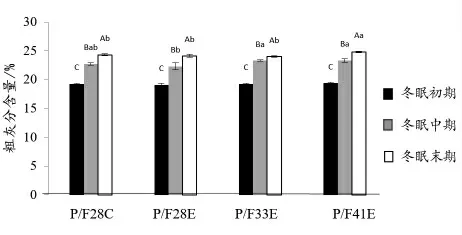
图3 中华鳖在冬眠不同时期全鳖粗灰分含量比较
2.2.2 冬眠中华鳖内脏常规营养成分分析 由图4 可知,中华鳖内脏粗蛋白质含量随冬眠时间的延长,逐渐上升,P/F28E 上涨趋势较小,粗脂肪含量逐渐下降,但P/F28C 在冬眠末期时,粗脂肪含量显著性上升(P <0.05)(图5),粗灰分含量整体趋势为先下降,再上升(图6),除P/F28E外,其他试验组均表现为冬眠末期时中华鳖内脏灰分显著性高于冬眠中期(P <0.05),低于冬眠初期(P <0.05)。
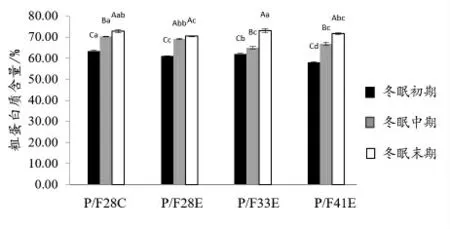
图4 中华鳖在冬眠不同时期内脏粗蛋白质含量比较

图5 中华鳖在冬眠不同时期内脏粗脂肪含量比较
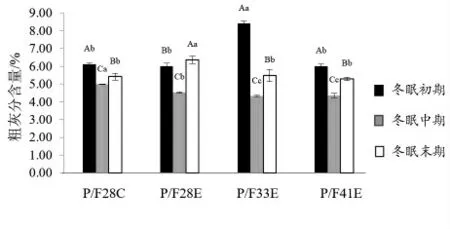
图6 中华鳖在冬眠不同时期内脏粗灰分含量比较
2.3 中华鳖肝脏、肠道和胃在冬眠不同时期的消化酶活性
2.3.1 蛋白质酶活性 由表4 可知,冬眠期间,中华鳖肝脏、肠道和胃的蛋白质酶活力整体表现为先下降后上升,冬眠初期,P/F28C 和P/F33E 组肝脏蛋白质酶活显著性高于其他两组(P <0.05),P/F28E 组肠道蛋白质酶活最低,显著性低于对照组和P/F33E 组(P <0.05),P/F28E 组胃蛋白酶活性较对照组高2.07(P >0.05),P/F33E 和P/F41E 组较对照组分别低10.02 和14.03(P <0.05);冬眠中期,P/F28E 和P/F41E 组肝脏蛋白酶活性较对照组分别高83.33 和77.28(P <0.05),P/F33E 组肝脏蛋白酶活性较对照组低17.73(P >0.05);冬眠末期,P/F41E 和P/F28E 两组肝脏、肠道和胃蛋白质酶活均显著性低于对照组(P <0.05),P/F33E组肝脏蛋白质酶活较对照组低37.42(P <0.05),但肠道和胃蛋白质酶活较P/F28C 组无显著性差异(P >0.05),较P/F41E 和P/F28E 两组肝脏、肠道和胃蛋白质酶活显著性升高(P <0.05)。P/F41E组肝脏、肠道和胃蛋白质酶活最低(P <0.05)。
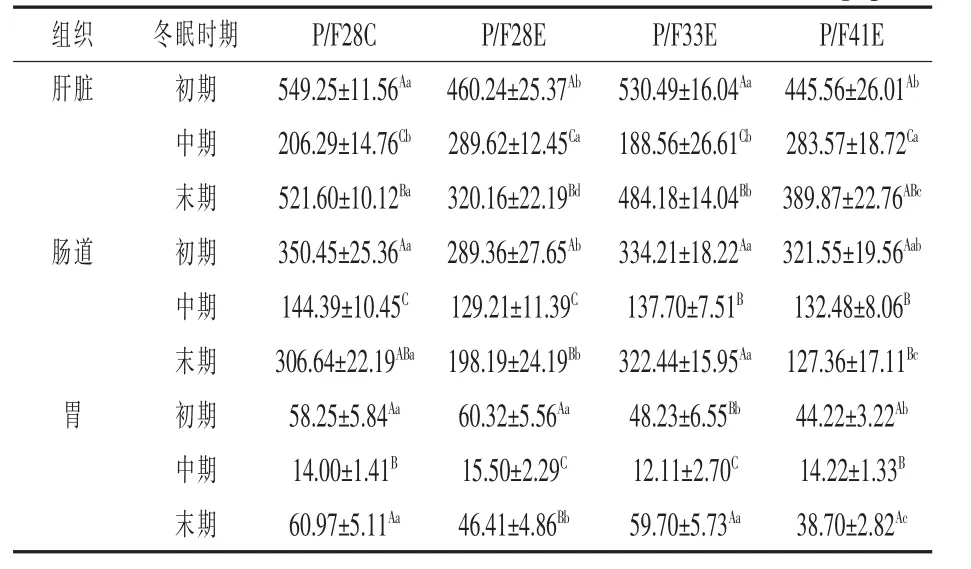
表4 不同配合饲料对冬眠期间中华鳖肝脏、肠道和胃蛋白质酶的影响U/g·pro
2.3.2 淀粉酶活性 由表5 可知,冬眠期间中华鳖肝脏、肠道和胃的淀粉酶活整体表现为先上升后下降,均表现为冬眠中期淀粉酶活性显著性高于冬眠初期和末期。P/F33E 组肝脏淀粉酶活在冬眠前、中、后期均显著性高于其他试验组(P <0.05),P/F41E 组肠道淀粉酶活性在冬眠期间均显著性高于其他试验组(P <0.05),P/F28E 组在冬眠初期胃淀粉酶活性显著性高于其他试验组(P <0.05),P/F28E 和P/F41E 组在冬眠中期胃淀粉酶活性显著性低于其他两组(P <0.05),P/F33E 组在冬眠末期胃淀粉酶活性显著性高于其他试验组(P <0.05)。
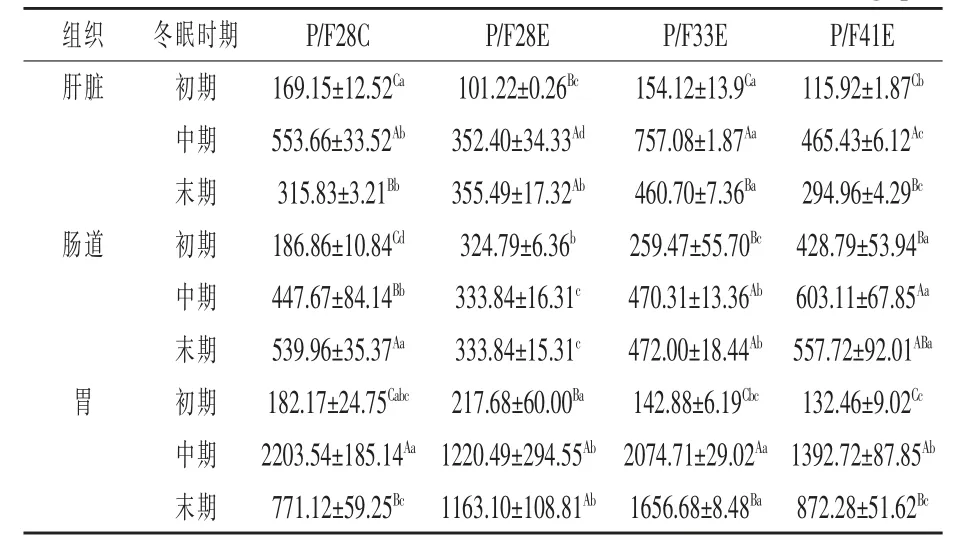
表5 不同配合饲料对冬眠期间中华鳖肝脏、肠道和胃淀粉酶的影响U/mg·pro
2.4 血清生化指标 由图7 可知,冬眠前期P/F28C 和P/F33E 组血清中溶菌酶活力显著高于P/F33E、P/F41E 两组(P <0.05);由图8 可知,冬眠期间,中华鳖血清中碱性磷酸酶活性整体表现为先下降后上升,P/F33E 组在冬眠初和冬眠中期血清中碱性磷酸酶活力显著性高于其他试验组(P <0.05)。从图9 可知,在冬眠前期,P/F41E 组总超氧化物歧化酶(T-SOD)显著高于其他试验组,中华鳖冬眠期间,血清中超氧化物歧化酶(SOD)活力整体表现为冬眠中期最低,冬眠末期最高,P/F33E组冬眠期间血清SOD 活力波动最小。

图7 不同饲料组中华鳖血清溶菌酶活力比较
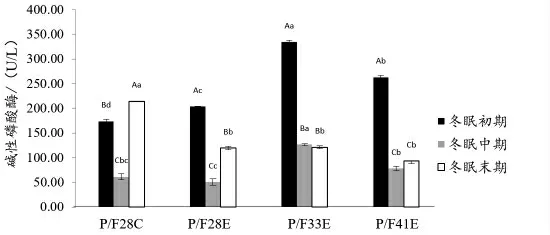
图8 中华鳖在冬眠不同时期血清碱性磷酸酶活力
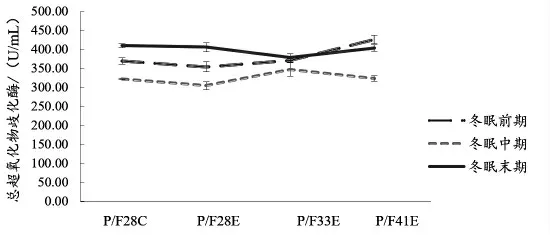
图9 中华鳖在冬眠不同时期血清T-SOD 活力
3 讨论
Ringo 等(2004)研究表明,豆粕中的抗营养因子中抗胰蛋白酶因子和植酸会抑制机体对脂肪和蛋白质的转化和沉淀,发酵豆粕因抗营养因子较低和良好的适口性,被认为是取代去皮豆粕的最佳饲料原料,并已在畜禽和鱼虾上取得良好的应用效果,比如,使用50%~ 70%的发酵豆粕替代鱼粉不影响牛蛙的生长性能(陈明哲等,2021);用8%的发酵豆粕替代鱼粉对罗氏沼虾的生长没有影响(杨景丰等,2018)。大米蛋白粉是一种新型的植物蛋白质原料,已有研究表明,饲料中使用大米蛋白粉替代鱼粉对保育猪的生长性能没有影响(田冬冬等,2016);畜禽和鱼虾对大米蛋白粉的营养物质消化率优于豆粕和发酵豆粕(刘卫东等2010),因此,在饲料配方中提高发酵豆粕和大米蛋白粉,并辅以降低去皮豆粕和国产鱼粉的用量,来降低进口鱼粉使用量,从而降低饲料生产成本,是一种相对稳妥可行的方案。
在本次养殖试验中,P/F33E 组较P/F28C 组在生长、形体指数、体成分方面均无显著性差异。这是因为P/F33E 组在降低进口鱼粉1 的使用量时,提高了进口鱼粉2、发酵豆粕和大米蛋白粉的使用量,来补充饲料中的蛋白质,随之降低了国产鱼粉2 和去皮豆粕的使用量,规避了国产鱼粉和豆粕对中华鳖机体的损伤。P/F41E 组较P/F33E 组和P/F28C 组在生长和形体指数均无显著性差异,但却显著性降低了鳖体粗脂肪,提高了内脏粗脂肪含量。可能是因为在进一步降低进口鱼粉1 的使用量时,采用在P/F33E 组的基础上增加进口鱼粉2和大米蛋白粉使用量的方法,从总体上提高了饲料中植物蛋白质源的比例,促使中华鳖体内糖原异生转化为脂肪,储存于肝脏中,随着投喂时间延长,导致该组中华鳖患脂肪肝的可能行越大,不利于中华鳖健康。P/F28E 组在生长、形体指数和体成分(除鳖体粗脂肪和内脏粗蛋白质)均值均劣于P/F28C组,但无显著性差异,而鳖体粗脂肪显著性高于其他试验组,内脏粗蛋白质显著性低于P/F33E 组和P/F28C,可能是因为在仅使用进口鱼粉2 的情况下,提高发酵豆粕使用量,不足以维持氨基酸平衡,影响了蛋白质消化吸收和利用。
水产动物体内的溶菌酶(LSZ)是一种重要的非特异性防御因子,也是生理防御水平的重要标志之一(Paulsen等,2001),碱性磷酸酶(AKP)是溶酶体酶的重要组成部分(Hashimoto等,1997),与脂类、葡萄糖、钙和无机磷的吸收存在正相关性(冯晓燕等2003),并具有防御和消化的双重作用(周永灿,2000)。在养殖试验期间P/F33E 组和P/F28C 组溶菌酶活力显著性高于其他两组,P/F33E组碱性磷酸酶显著性高于其他试验组,说明仅仅依靠提高发酵豆粕或大米蛋白粉的用量来降低进口鱼粉1 的用量可能会降低机体免疫水平。对大西洋鲽(Aksne等,1997)、大菱鲆(Oliva-Teles等,1999)和半滑舌鳎(柳旭东等,2006)的研究也表明,低质鱼粉会显著性降低机体的免疫力水平。总超氧化物歧化酶的活性可以表征机体抗氧化状态(Drail等,2001),相关研究表明低质鱼粉的氧化油脂会致使水产动物的总超氧化物歧化酶活性升高(Mourente等,2002)。在本试验中P/F41E 组TSOD(即冬眠前期)显著高于其他试验组,可能是因为低质鱼粉中不利因子含量和植物蛋白用量超过了中华鳖的耐受度,对其造成了胁迫。因此,在养殖试验期间,就生长、形体指数、体成分、血清免疫酶活和价格等因素而言,P/F33E 组配方最优。
冬眠是生物对外界不良环境条件的一种适应现象。冬眠期间,中华鳖主要通过皮肤呼吸和消耗体内积累的营养物质维持微弱的新陈代谢。经过冬眠的鳖体质量通常会减轻10%~15%(张幼敏等,1993)。已有研究表明,冬眠期低温降低了蛙类免疫等保护机能,同时也限制了病原微生物的内禀增长率,但体质较差的个体易患过冬死亡症(井润贞等,2011)。在冬眠期间,P/F33E 组和P/F28C组中华鳖体重变化幅度最大,且能在冬眠末期(复苏时)体重快速增长,表明两组中华鳖既能有效利用能量以顺利度过冬眠期,也能在复苏时快速适应环境和摄食。
饲料中鱼粉的组胺含量低时,可以促进消化酶的分泌以及提高其活性(Sugahara等,1988),但含量高时会导致消化道损伤,抑制消化酶的分泌和活性,从而降低饲料利用效率和养殖动物生长(Okazaki等,1983)。国产鱼粉的组胺含量高于进口鱼粉,豆粕中的胰蛋白酶抑制因子会降低机体蛋白质酶的活性(Yamamoto等,1998)。在本次养殖试验中,P/F28C 和P/F33E 组肝脏、肠道和胃的蛋白质酶活显著性高于其他两组,P/F28E 可能是因为饲料中国产鱼粉含量较高,其中的组胺抑制了机体蛋白质酶活力;而P/F41E 组蛋白酶活力低,可能与动植物蛋白源使用比例不合理有关;P/F33E 组蛋白质酶活力较高的原因,可能是饲料中复合鱼粉的使用降低了饲料中组胺的含量,复合植物蛋白质源抑制了饲料中胰蛋白酶抑制因子的活性。中华鳖是一种变温爬行动物,环境温度的变化会影响动物机体代谢过程中的酶活和酶浓度的变化(Crawford等,1999)。因此在本次冬眠试验中,蛋白质酶活力呈现先下降后上升的趋势,P/F28C 和P/F33E 组在冬眠末期各组织蛋白质酶活均上升至冬眠初期水平,可能是该两组中华鳖体质较好,能快速恢复活力并摄食。
淀粉酶主要是由肝脏产生,并在肠道中激活(Debnath等,2007),且长期摄食高植物蛋白质饲料可提高机体淀粉酶活性。在本次养殖试验中,中华鳖机体淀粉酶活性整体表现与饲料中植物蛋白质源呈正相关。葡萄糖是生物机体的主要防冻剂,可降低生物在冬眠期间的死亡风险(Storey等,1987)。当年气温变化无常,扰乱了中华鳖机体代谢,使得中华鳖过早恢复摄食,然而此时并未及时投喂,只能摄食水体中枯枝败叶和已死亡的中华鳖,在本次冬眠试验期间,淀粉酶活力整体表现为先上升后下降,但后期仍高于初期,推测与整个冬眠期糖类的分解供能有关,也可能与复苏的中华鳖主要摄食水体中的植物有关。中华鳖的脂肪主要存储于肝脏和四肢脂肪腺中。冬眠期间,中华鳖鳖体和内脏中粗脂肪含量显著性减少,且肝体比也在显著性下降,而中华鳖鳖体和内脏中粗蛋白质和灰分含量显著性上升,表明冬眠期间,中华鳖消耗的能源物质除糖类外还有脂肪。
中华鳖是一种变温爬行动物,所以免疫功能具有明显的温度依赖性,且冬眠会抑制机体免疫(Bouma等,2010)。因此,冬眠期间中华鳖碱性磷酸酶和总超氧化物歧化酶活力整体表现为先下降后上升趋势。已有研究表明(Gregory等,1997),蛙类在低温驯化初始阶段会导致免疫抑制,适应后会恢复至或上升到更高水平,而P/F41E 组冬眠末期T-SOD 显著性低于冬眠前期,可能是此时中华鳖机体免疫器官受冬眠影响而遭到破坏,故不建议采用这样的组合替代方法降低进口鱼粉的使用量。因此,在冬眠试验期间,综合生长、形体指数、体成分、血清免疫酶活力和价格等因素,P/F33E组配方最优。
4 结论
采用提高大米蛋白粉和发酵豆粕使用量,并同时降低去皮豆粕和国产鱼粉使用量的组合方式来降低配方中进口鱼粉的使用量,进而降低饲料生产成本的方法是可行的,综合生长、形体指数、体成分、血清免疫酶活和价格等因素,P/F33 E 组配方最优。
