不同生境下荠菜种群构件生物量分配特征
2022-08-04李恒艳楚合拉恩德马克黄婧姝张丽辉张彦文
李恒艳, 楚合拉·恩德马克,李 璐,黄婧姝,张丽辉,张彦文
(长春师范大学生命科学学院,吉林 长春 130032)
异速生长是指生物体某一特征与另一特征的相对生长速率不相同的现象,可用于表达生物体的某些特征与个体大小之间不成比例的幂函数关系[1]。通过异速生长分析可很好地阐释植物生物学的内在规律,对植物生活史策略的研究有十分重要的意义。植物异速生长的研究源于生物量分配[2]。而生物量作为植物的基本生物学特征和功能性状之一,是植物的物质和能量积累的基本体现[3]。因此,通过分析植物生物量分配情况,可以了解植物的生长状况、繁殖策略以及其在不同生境下的变化。
早春植物是指在上层林木仍处于休眠状态的早春时期就开始生长,于林分郁闭前开花、结果,林分郁闭后进入休眠期的多年生草本植物[4]。其普遍生存于温带落叶阔叶林下,是落叶阔叶林中的一个特殊的生态层片,并在森林生态系统的能量流动和物质循环中起着重要的作用[4-5]。研究表明,绝大多数的早春植物都具有很好的经济价值、观赏价值以及药用价值[6]。因此,关于早春植物的研究多集中在生物学特征、种群群落特征以及药理学方面,而对其生物量分配和生长分析的研究仍相对匮乏,其所蕴含的生物量分配特点和异速生长规律仍知之甚少。
荠菜(Capsellabursa-pastoris)又名地菜、护生草、菱角菜,是十字花科(Brassicaceae)荠属(Capsella)的一年生或二年生草本植物。其作为一种典型的早春植物,具有很高的营养价值和药用价值。目前,关于荠菜的研究主要体现在其生物学特性[7]、食品工艺、药理学以及重金属污染土壤治理[8]等方面,而对于其生物量的研究仅在张峰等人对四种早春植物生物量的动态研究中被涉及[9]。因此,本文以3种生境下的荠菜植株作为实验材料,通过对不同生境下荠菜各构件的生物量结构和构件间生长关系的定量分析,探究其在不同生长条件下所呈现出的生物量分配特点及生长、繁殖规律,为深入了解荠菜这类早春植物的生活史策略提供科学依据,并为该种植物资源的开发利用提供理论参考。
1 材料与方法
1.1 研究样地自然概况
研究区域位于吉林省长春市伊通河外围(东经125°21′24″~125°27′12″,北纬43°50′06″~43°57′36″)。此地区属于北温带大陆性半湿润季风气候,四季分明。年均温度4.6℃,四月均温7.8℃,年降水量600~700 mm,全年无霜期为140~150天,冰冻期为5个月。
1.2 植物样品采集与处理
2021年4月至5月期间,在对荠菜生长分布地区全面踏查后,于荠菜的花果期,分别选择了林缘(Forest edge,F)、路边(Roadside,R)、裸地(Naked land,N)3个生境。于每个生境中随机采集30株生长发育良好的荠菜植株,用小尖铲连带根系全株挖出,清水洗去植株上的泥土后,按生境将荠菜样品分别标上序号(1~30),测量每株样品的株高,而后将植株的根、茎、叶、花序分别包装,标好序号整合装袋,置于烘干箱105℃杀青30 min后,于60℃烘干24 h。将每株样品的根、茎、叶、花序单独称量,记录数据。天平精度为0.000 1 g。
1.3 数据统计与分析
以株为单位统计株高以及各构件的生物量及其分配指标,其中数据的整理、统计与图表的制作采用 Excel 软件,并利用 SPSS 16.0 统计软件对数据进行单因素方差分析和多重比较。此外在处理各生物量间关系时,选择相关系数最大的函数作为描述模型进行分析。
2 结果
2.1 不同生境下荠菜各构件的生物量结构分析
不同生境下荠菜种群各构件生物量特征见表1。从表1可知,林缘(F)、路边(R)、裸地(N)的平均株高分别为17.76 cm、28.47 cm、29.48 cm,其中林缘荠菜的平均株高显著低于路边和裸地荠菜(P<0.05),并以裸地荠菜的平均株高最高,路边荠菜次之。此外,根据数据可知这三种生境下的荠菜株高变异系数均低于50%,表明各生境下的荠菜种群的株高相对稳定。
通过表1分析这3种生境的生物量可知,裸地的荠菜种群总生物量最大,其构件生物量由大到小依次表现为叶、茎、花序、根,且各项生物量的变异系数均超过50%,以花序生物量的变异系数最高,为97.42%,茎生物量次之,为76.24%;而路边的荠菜种群总生物量次之,其构件生物量由大到小依次表现为茎、叶、花序、根,各项变异系数也均超过50%,以叶生物量的变异系数最高,为82.57%;林缘的荠菜种群总生物量最低,其构件生物量由大到小依次表现为叶、茎、根、花序,变异系数分别为56.49%、45.67%、56.31%、43.75%,相较于裸地和路边的荠菜植株变异系数最小。此外,对比各生境的生物量分配可知,各生境荠菜的生物量分配与其构件生物量呈现相同的规律,且其变异系数均低于50%。而对比这3种生境下荠菜种群各构件生物量分配的变异系数可知,林缘荠菜生物量分配变异系数由大到小依次表现为茎、花序、叶、根,路边则表现为根、花序、叶、茎,裸地则表现为花序、茎、叶、根。
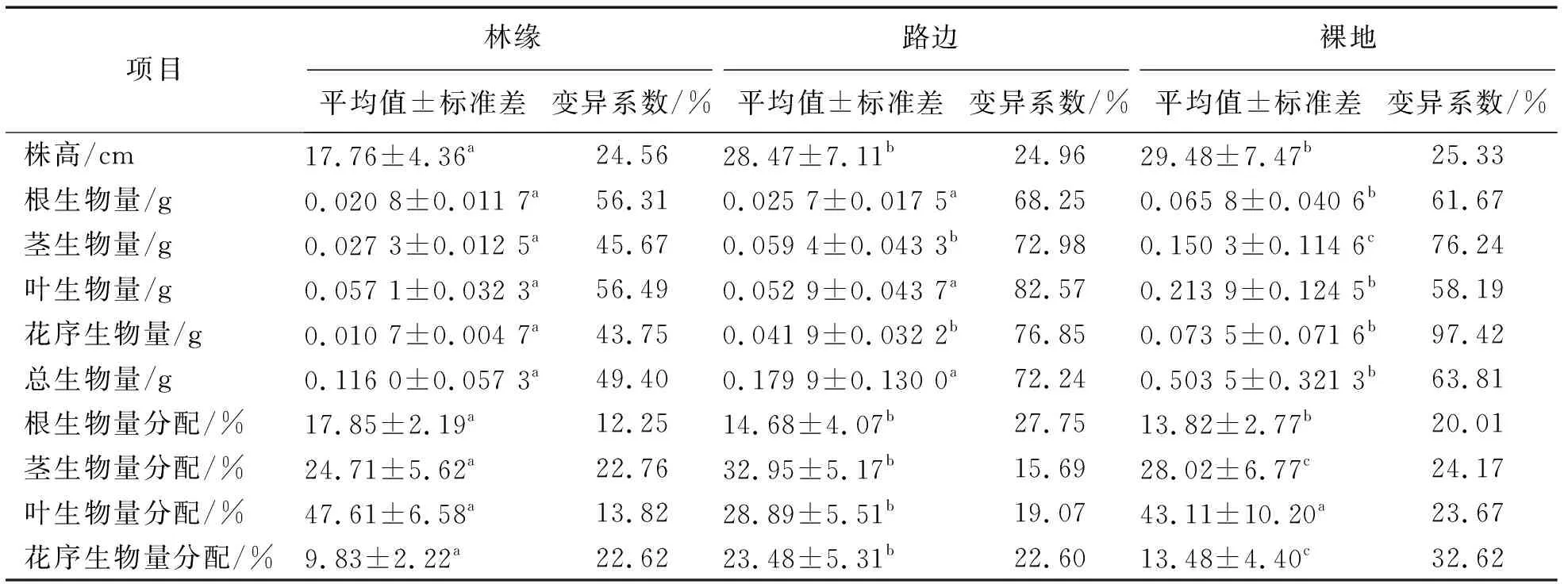
表1 不同生境下荠菜种群各构件生物量特征(n=30)
2.2 不同生境下荠菜各构件生物量与植株大小之间的关系
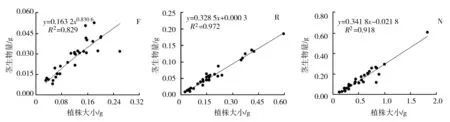
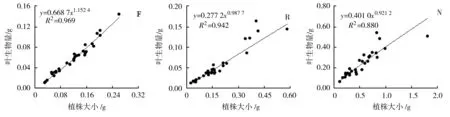
3种生境下荠菜各构件生物量与植株大小(总生物量)之间的关系见图1。

(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
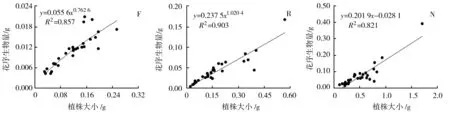
(j) (k) (l)图1 不同生境下荠菜种群各构件生物量与植株大小的观测值及拟合曲线(n=30)
由图1可知,荠菜在花果期其根、茎、叶以及花序的生物量均与其植株大小呈现极显著的正相关关系(P<0.01),即荠菜植株的各构件生物量随其植株大小的增加而增加。其中林缘生境下的荠菜根、茎、叶、花序的生物量与植株大小均呈现幂函数正相关,反映了该生境下荠菜种群各构件生物量与植株大小之间具有异速增长规律,但增长速率不同。据其方程b值可得,林缘生境下的荠菜植株与植株大小之间的异速生长指数由大到小依次表现为叶(1.152 4)、根(0.997 5)、茎(0.830 6)、花序(0.762 6),可见随着植株大小的增大,林缘荠菜植株会将更多的资源优先分配给光合构件(叶),根、茎及花序的生物量分配速率则逐渐减少。而路边生境下的荠菜根、叶、花序的生物量与植株大小呈现幂函数正相关,茎生物量与植株大小则呈极显著线性正相关;裸地生境下的荠菜根和叶与植株大小呈现幂函数正相关,茎和花序与植物大小则呈现极显著的线性正相关。幂函数关系体现其异速生长规律,而线性函数关系体现其同速生长规律。表明路边和裸地生境下的荠菜植株存在异速生长和同速生长并存的现象,并在物质能量有序分配的基础上,呈现了不同的表型可塑性调节。
2.3 不同生境下荠菜的繁殖器官与营养器官生物量之间的关系
三种生境下荠菜繁殖器官(花序)与营养器官(根、茎、叶)生物量之间的关系见图2。林缘和路边荠菜的繁殖器官与营养器官的生物量之间均呈现极显著的幂函数正相关关系(P<0.01),体现了这两种荠菜种群的繁殖器官与营养器官是异速生长关系,不同生境下生长速率不同。据方程b值可知,林缘和路边荠菜的繁殖器官和营养器官之间的异速生长指数分别为0.732 2、0.989 7,均小于1,说明二者荠菜植株的繁殖器官生物量分配速率均小于其营养器官。而裸地的荠菜种群繁殖器官与营养器官生物量之间呈现极显著的线性正相关关系,说明裸地荠菜植株的繁殖器官与营养器官成比例生长,即同速生长。据其方程b值可知,裸地荠菜的繁殖器官生物量对营养器官生物量的制约力为0.027 8。可见,各生境下的荠菜植株的营养物质积累及其向营养器官和生殖器官的分配是协调有序的,但所遵循的规律和速率不尽相同,具有不同过程的表型可塑性调节。

(a)林缘生境 (b)路边生境 (c)裸地生境图2 不同生境下荠菜种群繁殖器官与营养器官生物量的定量关系模型(n=30)
3 讨论与结论
在植物的生长过程中,为了确保其在不同生境下取得最大的生长适合度,往往会将从外界获得的有限资源合理分配到不同构件上。荠菜作为十字花科植物里较为娇小的一类,由于其枝条柔韧可以随风飘荡,地下部分受力不大,所以主根普遍短小[10]。因此,荠菜植株往往会更优先将资源投入到地上部分的生长发育上,而不是地下部分。研究表明,林缘、路边、裸地这3种生境的荠菜植株在生物量分配上都倾向于将更多资源分配给支持构件(茎)和光合构件(叶),并在植株各构件生物量分配上都表现出较低的变异系数(<50%)趋于稳定的表型。但不同生境下的荠菜种群为了更好地适应其生境在植株大小、构件生物量结构上呈现了显著差异。裸地生境的光照最为充足,土壤资源也相对丰富,因此,此生境下的荠菜种群长势最好,株高最高、总生物量最大,同时植株在构件生物量上也表现出较高的变异系数,尤其是花序生物量的变异系数高达97.42%,表现出很高的表型可塑性。其次是路边的荠菜种群,此生境下的荠菜植株为了更好地适应光照不足、土壤质地较硬、种群竞争激烈的生长环境,将外界获得的有限资源更多的分配给了支持构件(茎),因此株高近似于裸地的荠菜种群,但在总生物量上远远低于裸地荠菜种群,并在叶生物量上表现出了较高的变异系数。而林缘生境的荠菜种群较前两者是株高最矮、总生物量最小的,虽然该生境较前两个生境土壤更为肥沃、阳光更为充足,但该生境下种群间的竞争尤为激烈,从而使得荠菜在资源获取上更为困难,因此林缘生境的荠菜植株在繁殖构件(花序)和营养构件(根)中更倾向于将资源分配给营养构件,同时在各构件生物量的变异系数上较于前两者而言更低,在构件表型上更为稳定。
植物生长过程中存在着同速生长型和异速生长型两种模式[11]。同速生长型往往在构建的模型中表现为直线函数形式,而异速生长型则为非线性幂函数或指数函数形式[12]。不同生境的荠菜植株在构件生物量与植株大小之间所遵循的增长规律有所不同。林缘的荠菜植株大小随其根、茎、叶及花序生物量的增加均呈幂函数形式增长;而路边的荠菜植株大小随其根、叶及花序是生物量的增加呈幂函数形式增长,随其茎生物量的增加呈直线函数形式增长;裸地的荠菜植株大小则随其根、叶生物量的增加呈幂函数形式增长,随其茎、花序生物量的增加呈直线函数形式增长。可见这3种生境下的荠菜植株都存在异速生长规律,其中路边和裸地的荠菜种群甚至出现了异速、同速生长规律并存的现象,这是荠菜植株为了更好地适应各自的生境而做出的内外兼顾的生长策略。
植物在生长发育中,于生长与物质分配相协调的基础上,会不断地调整其生殖生长与营养生长的生物量分配策略[13-14]。研究表明,荠菜植株营养器官的生物量会随着繁殖器官生物量的增加而增加,于不同生境下呈现不同的生长规律。林缘和路边的荠菜植株的繁殖器官与营养器官之间遵循异速生长规律,且异速指数均小于1,表明随着荠菜营养器官生物量的增加,繁殖器官的生物量分配逐渐减少,更多的外来资源优先用于植株营养生长上。而裸地的荠菜植株的繁殖器官与营养器官之间遵循同速生长的规律,二者成比例生长。由此可见,在不同生境下,荠菜植物采取了在生殖生长和营养生长之间进行权衡的资源分配策略,以确保亲本的存活,同时最大可能地产生种子延续后代,蕴含了荠菜种群生长和繁殖的策略。
