井冈山地区莲鱼复合种养模式效益评价
2022-08-04米湘媛张朝硕张晓娟王海华郭小泽李思明赵大显洪一江段明
米湘媛 张朝硕 张晓娟 王海华 郭小泽 李思明赵大显 洪一江 段明
(1.中国科学院水生生物研究所淡水生态与生物技术国家重点实验室,武汉 430072;2.中国科学院大学,北京 100049;3.江西省水产科学研究所,南昌 330039;4.江西省农业科学院畜牧兽医研究所,南昌 330200;5.南昌大学,南昌 330047)
渔农复合种养(Integrated Agriculture-Aquaculture,IAA)是指利用生物群落共生原理、系统内组分之间相互协调和促进及物质循环利用原理,通过产品多样化来维持系统生产力和分散经济风险的生产模式[1]。不仅能提高单位面积利润[2],还能减少农业用水[3]、减少化肥和农药的使用、减少作物病害[4]、降低温室气体的排放[5]、减轻养殖池塘中的富营养化程度[6]及增加农村就业人口等[7],被认为是以小农经济为主的发展中国家的一种可持续农业模式[8]。稻田养鱼是我国一种传统的农渔复合种养模式,至今已经有两千多年历史[9],在此基础上发展出来的稻-鳖[10]、稻-虾[11,12]、稻-蛙[13]、稻-鱼[14]和稻-鳅[15]等复合种养在我国得到了广泛的生产实践,在少数莲藕资源丰富的地区,还因地制宜地开展了莲-虾[16]和莲-鳖复合种养[17]。渔农复合种养还为稻田相对分散、海拔较高、水温偏低、作物亩产量相对平原地区较低的山区提供了相比单一种植水稻更为经济高效、生态友好的可持续发展生产模式,成为我国四川广元[7]、贵州遵义[18]、浙江青田[4,19]和江西井冈山[17]等山区农业产业结构调整、稳粮增效、带动当地农户脱贫致富的有效手段。
浮游植物群落结构不仅能反映水体的营养状态,还是衡量水体生态系统健康程度的重要指示生物[20]。浮游植物对水体状况十分敏感,水体理化因子如营养盐、温度、pH和溶氧等都会对水体中的浮游植物产生影响,不同环境的主要影响因子各不相同且存在较大差异[21,22];而氮、磷长期以来被认为是影响浮游植物群落结构的重要环境因子[23]。有研究表明,复合种养系统中藻类不仅能通过光合作用增加水体中的氧气,还能加快尿素的水解,对氮元素转移与转化十分重要,然而过多的藻类又会因对氮元素的竞争作用影响作物的生长[24]。此外,复合种养系统中浮游植物群落多样性的增加,有助于形成更复杂的食物网结构,从而提升生态系统的稳定性和生产力[25]。研究不同种养系统中浮游植物群落结构及其与环境因子的关系可为制定合理的水质、藻相管理措施提供科学依据。
目前,关于渔农复合种养模式中水体的浮游植物群落特征研究,多以稻渔复合种养为主,缺乏对山区莲渔复合种养模式的研究;对渔农复合种养模式的效益评价多以直接收益法和产投比法[8,26]为主,或通过其化肥和农药减施量描述其生态效益[4],缺乏对其经济和生态效益统一量纲的、定量的综合评价。本文选择江西省井冈山地区具有代表性的莲鱼复合种养模式作为研究对象,以当地鱼单养和莲单种模式作为对照模式,研究该复合模式中浮游植物的群落结构特征,采用冗余分析研究该模式中驱动浮游植物群落动态变化的环境因子,基于生态足迹法[27,28]对其生态效益进行统一量纲的分析。为莲鱼复合种养模式的水质、藻相管理提供理论基础,为渔农复合种养模式的效益评价提供参考。
1 材料与方法
1.1 研究地点
研究地点位于中国江西省吉安市井冈山市(东经113°49′—114.23′,北纬26°27′—26°49′),该地区年平均温度19℃,年降水量1597.5 mm,年日照时数1698.7h,年平均湿度可达78%。其中,莲鱼复合种养模式及鱼单养模式均来自同一生产基地(大陇,图 1),莲单种模式来自该地区另一生产基地(田头,图 1),两研究地点相距41 km,采集的池塘面积均在(0.133±0.05) hm2,两个基地种植的莲品种均为太空莲36号。太空莲36号全生育期(移栽至采摘结束)200—210d,井冈山地区种植的太空莲在春季开始立叶,秋季下旬结束采摘,本研究在该时期进行。莲鱼复合种养模式环沟宽度在1—1.5 m左右,沟深1.5 m。莲鱼复合种养及鱼单养模式中放养的均为中科五号异育银鲫,莲鱼复合种养模式放养量为195 kg/hm2,鱼单养系统放养量为225 kg/hm2。
图1 研究地点位置Fig.1 The locations of two study sites
1.2 生产调查及样本采集
生产调查通过实地勘查、访问农户、查阅生产记录本及调查问卷获取生产数据。采集的数据包括水产品的放养量、收获量、人工成本、田租、投饲量、用水量、肥料及农(渔)药施用量,并通过其单位面积的收入减去单位面积的成本得到单位面积利润。
水样、浮游植物的采集、测定在4月(春季)、7月(夏季)和10月(秋季)对莲鱼种养、鱼单养和莲单种模式各三个池塘进行水样和浮游植物的采集。水样采集依据 HJ 494—2009《水质采样技术指导》和 SL 187—1996《水质采样技术规程》所要求的方法,采集的水样保存在1 L水样瓶中。除使用便携式水质测定仪YSI现场测定溶氧、水温和pH外;其他参数均通过现场固定后运回实验室进行测定。浮游植物的采集参照《淡水浮游生物研究方法》[29],浮游植物定性样品用25#浮游生物网在水深0.5 m处缓慢划“8”字进行采集,拖拉时间3min。定量样品使用5 L的有机玻璃采水器在水深50 cm处进行采集,取1 L水样加入鲁哥氏液固定后带回实验室进行分析。将水样沉淀并浓缩至50 mL后于显微镜下计数。参照相关文献[30]进行藻类种类鉴定,参照《淡水浮游生物研究方法》对浮游生物样品计数及计算。
1.3 计算公式
浮游生物多样性指数根据香农-威纳多样性指数(H',Shannon-Wiener index)、Pielou 均匀度指数(J,Pielou)和Mcnaughton 优势度指数(Y,Mcnaughton index)对不同系统水体中的浮游生物进行分析评价[31],各指数的计算公式如下:
(1)H'=-Σpi×lnpi
(2) Pielou 均匀度指数J=H'/log2S
(3) Mcnaughton 优势度指数Y=pi×fi
在此过程中,技术要点如下:(1)对电力系统的调峰能力进行科学合理的分析;(2)对系统的调频能力进行科学合理的分析;(3)对电力系统传输能力进行分析;(4)将消纳能力作为目标,对电力系统进行科学合理的规划。
式中,ni为i种的个体数,N为所有种类总个体数,H'为香农-威纳多样性指数,S为物种数,pi=ni/N,fi为该物种在各池塘中出现的频率。本研究中将Y>0.02的浮游植物定为优势种,Y>0.1为强优势种。
生态足迹生态足迹法通过计算一定区域内消费及排放废弃物所需要的生态生产性面积来表示发展造成的生态负荷,计算公式如下:
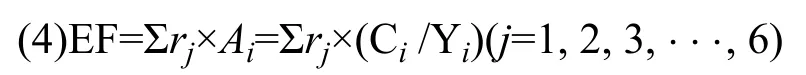
式中,EF为生态足迹总量,i为消费项目类型,j为生物生产性土地类型(分为耕地、林地、草地、水域、建筑用地和能源用地),rj为均衡因子,Ai为第i种消费项目折算的生物生产面积,Ci为第i种消费项目的消费量,Yi为生物生产性土地生产第i种消费项目该地区年平均产量。在本文中,饲料、苗种、水、农(渔)药、肥料氮、肥料磷和Ci值是通过生产调查得到的数据,水污染氮和水污染磷的Ci值是通过水质调查得到的水体中年平均总氮和总磷含量减去GB 3838-2002《地表水环境质量标准》中对该类水体规定的上限值得到的。该地区水资源年平均产量来源于2016—2019年《吉安市水资源公报》,饲料(粮食)、苗种(水产品)及生产性土地面积年平均产量来源于井冈山市2015—2019年《年经济和社会发展统计公报》。土地对农(渔)药、化肥中N、P的消纳能力及水体对N、P的消纳能力见文献[32]。
1.4 数据处理
数据处理及分析均在R 3.6.0中完成。采用单因素方差分析(ANOVA)中的多重比较(Tukey HSD)分析不同系统水体理化因子及浮游植物密度差异。采用Pearson检验分析不同水体理化因子之间的相互作用及水体理化因子对浮游植物密度、Shannon-Wiener指数和Pielou 均匀度指数的影响。
使用R 3.6.0 中的vegan包进行浮游植物与环境因子的排序分析,用于排序分析的物种数据筛选标准:(1)在各样点出现的频率>12.5%,(2)物种的Mcnaughton 优势度指数>0.02。对筛选出的物种密度数据进行Hellinger正态转化,对环境因子数据进行正态性检验,对不符合正态分布的数据进行转化。随后使用Decorana函数对物种数据进行除趋势对应分析,进一步得到各排序轴的物种变化梯度长度。
2 结果
2.1 浮游植物种类组成、密度及优势种
共采集到裸藻门(Euglenophyta)、隐藻门(Cryptophyta)、黄藻门(Xanthophyceae)、蓝藻门(Cyanophyta)、绿藻门(Chlorophyta)、硅藻门(Bacillariophyta)和金藻门(Chrysophytes)浮游植物7门103种,其中莲鱼种养模式90种,鱼单养模式85种,莲单种模式76种,3种模式的浮游植物种类数均为夏季>春季>秋季。在研究期间,采集到的绿藻门种类数最多,为54种,黄藻门种类数最少,仅有1种,各门种类数量排序为:绿藻门>蓝藻门>硅藻门>隐藻门>裸藻门>金藻门>黄藻门。

图2 莲鱼种养、鱼单养和莲单种模式水体中不同季节浮游植物细胞密度变化Fig.2 Seasonal densities of phytoplankton in the integrated lotusfish culture system,lotus monoculture system and fish monoculture system
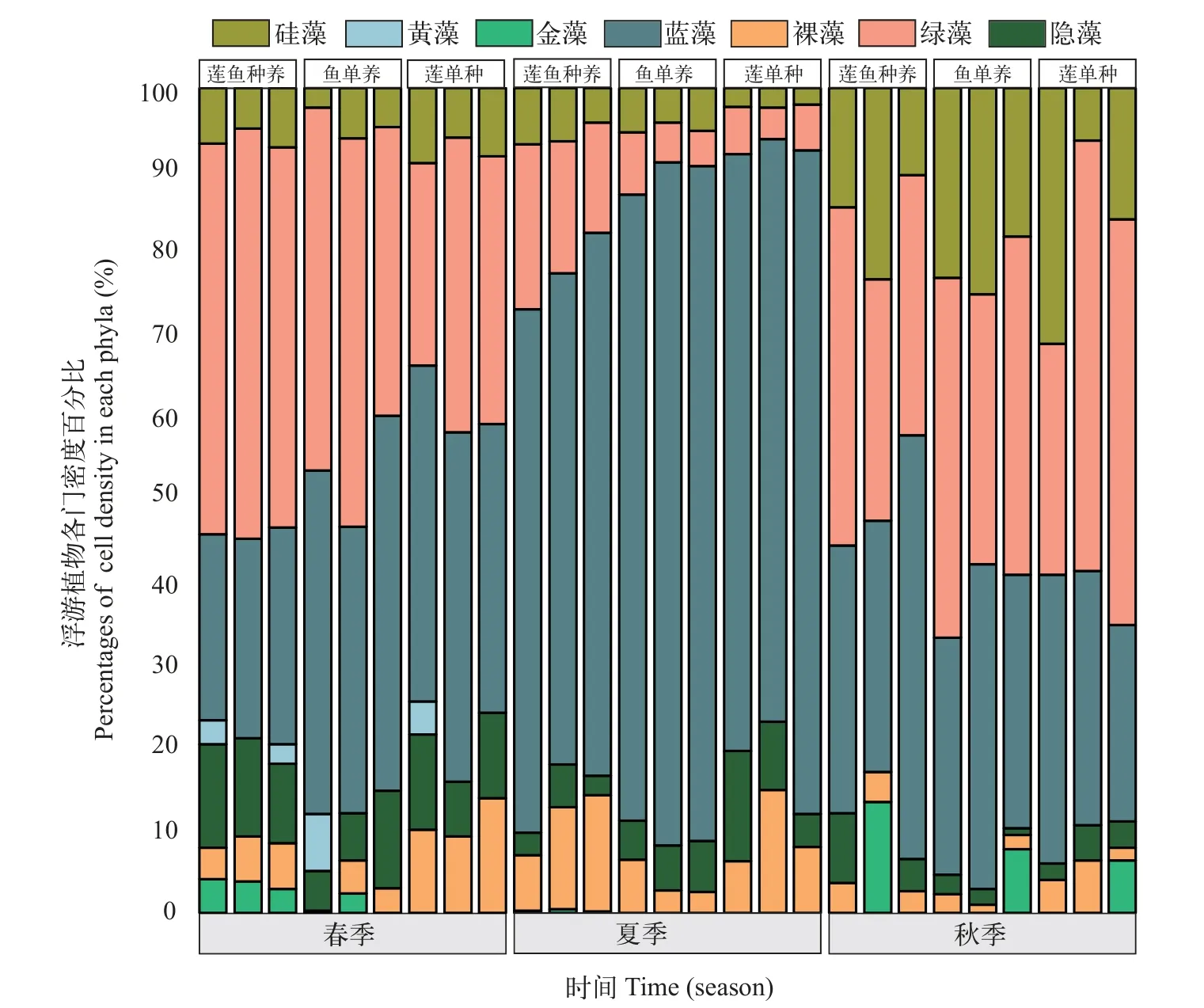
图3 不同季节莲鱼种养、鱼单养和莲单种模式水体中浮游植物细胞密度百分比Fig.3 Seasonal density in percentages for each phyla in the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
在调查期间,3种种养模式水体中的优势种组成及优势度具有明显差异(图4和表 1)。莲鱼复合种养模式水体中春季优势种为卵形隐藻(Cryptomons ovata;0.098)、美丽颤藻(Oscillatoria formosa;0.046)、针状蓝纤维藻(Dactylococcopsis acicularis;0.047)、四尾栅藻(Scenedesmus quadricauda;0.153)和双对栅藻(Scenedesmus bijuba;0.132);夏季优势种为矩圆囊裸藻(Trachelomonas oblonga;0.080)、小席藻(Phormidium tenus;0.097)、颤藻(0.140)、铜绿微囊藻(Microcystis aeruginosa;0.241)和四足十字藻(Crucigenia tetrapedia;0.052);秋季优势种为假鱼腥藻(Pseudoanabaenasp.;0.050)、空球藻(Eudorinasp.;0.022)、椭圆小球藻(Chlorella ellipsoidea;0.062)和异极藻(Gomphonemasp.;0.066)。在鱼单养模式中,夏季仅有蓝藻门的藻类形成优势种,且水华微囊藻、铜绿微囊藻和假鱼腥藻形成强优势种。在3种种养模式水体中均出现的优势种有:卵形隐藻、四尾栅藻、小席藻和铜绿微囊藻。
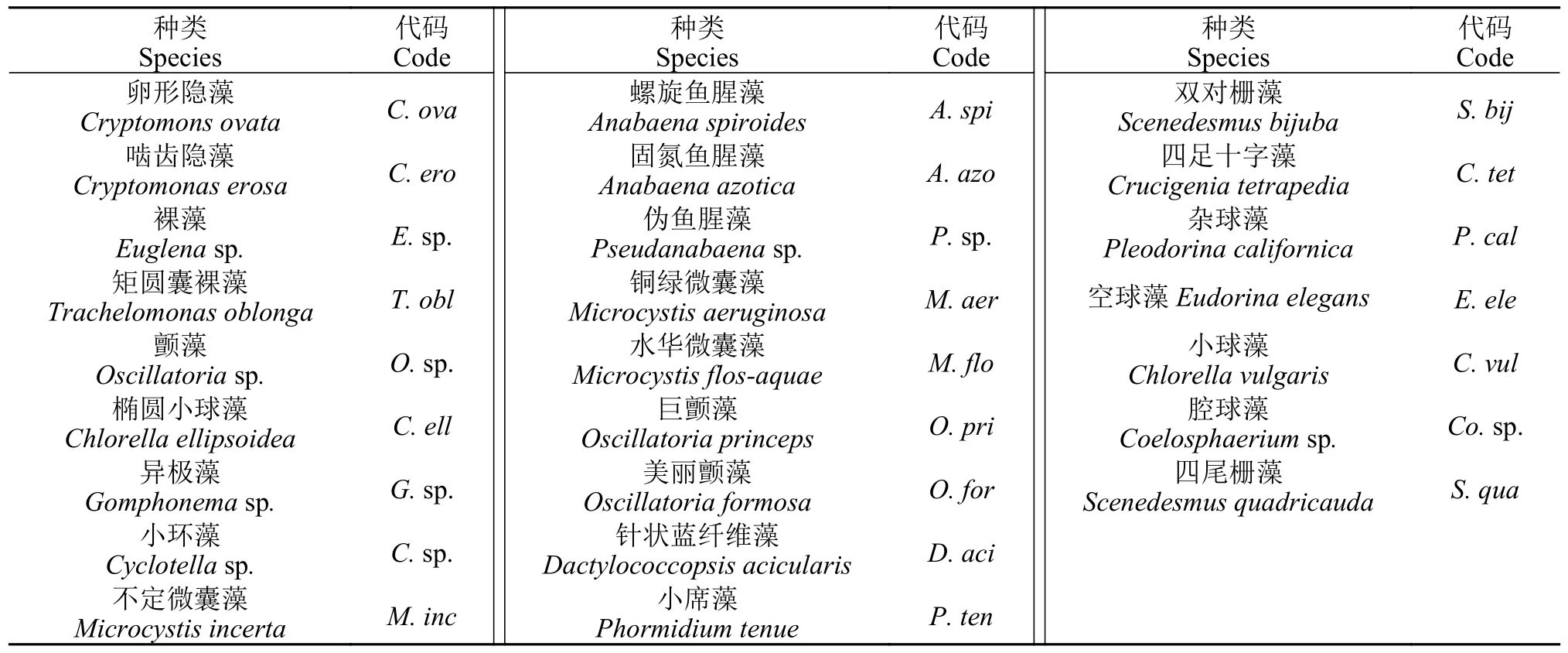
表1 浮游植物优势种代码Tab.1 Codes of dominant phytoplankton species

图4 不同季节莲鱼种养、莲单种和鱼单养模式中浮游植物优势度指数Fig.4 Seasonal variation of Mcnaughton dominance index in the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
2.2 浮游植物多样性指数
研究期间,各模式的香农威纳指数和均匀度指数均呈现出先下降后上升的趋势(表2)。其中,莲鱼复合种养模式的香农威纳指数变化为3.021—3.261;Pielou均匀度指数变化为0.502—0.645。鱼单养模式的香农威纳指数变化为2.649—2.882;Pielou 均匀度指数变化为0.454—0.613;莲单种模式的香农威纳指数变化为2.527—3.061;Pielou 均匀度指数变化为0.452—0.602。各模式香农威纳指数H′均值排序为莲鱼复合种养>莲单种>鱼单养,Pielou均匀度指数均值排序为莲鱼复合种养>莲单种>鱼单养,但3种模式水体中的浮游植物香农威纳指数和均匀度指数没有显著差异(P>0.05)。
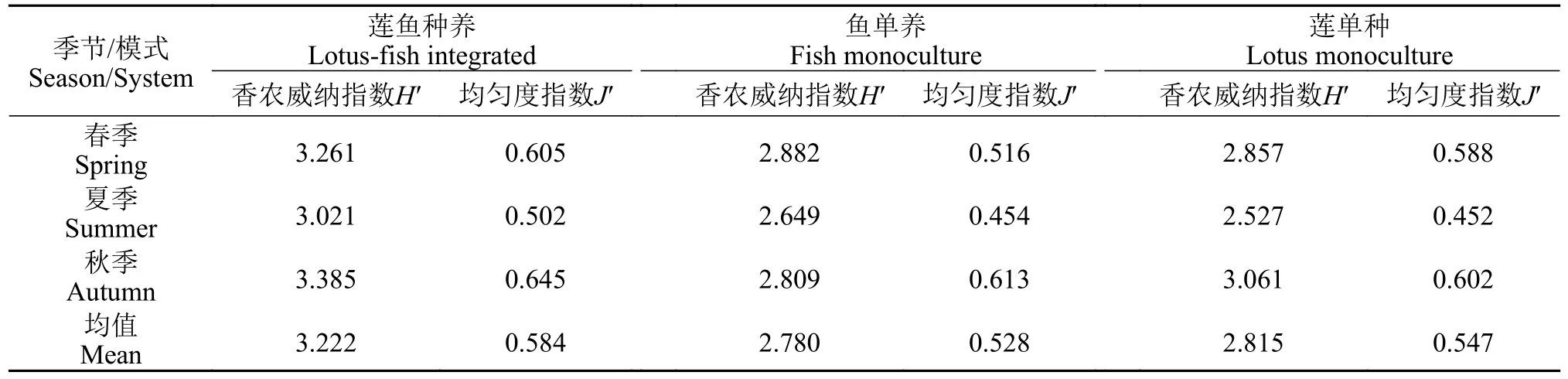
表2 不同季节莲鱼种养、莲单种、鱼单养模式浮游植物香农威纳指数及均匀度指数Tab.2 Seasonal variation of Shannon-Weiner index and Pielou index in the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
2.3 水体中的环境因子
莲鱼复合种养模式及其单种、单养模式水质呈现出一致的季节变化,水体中总氮(TN)、总磷(TP)、氨氮(NH4-N)、硝酸盐氮(NO3-N)、亚硝酸盐氮(NO2-N)和水温(WT)在复合种养期间均呈现出先升后降趋势,而pH和DO则呈现出先降后升趋势(图5)。单因素方差分析结果显示,春季莲鱼复合种养模式和莲单种模式的TN显著高于鱼单养模式,莲鱼复合种养模式水体中的TP含量显著低于鱼单养模式(P<0.05),莲鱼复合种养模式水体中的NO3-N含量显著低于鱼单养模式(P<0.001),但显著高于莲单种模式(P<0.001),水体中NO2-N含量显著高于莲单种模式(P<0.001),水体中溶氧显著高于鱼单养模式(P<0.01),水体中pH显著高于莲单种模式(P<0.05);此外,春季各模式间的WT和NH4-N没有显著差异。夏季莲鱼复合种养模式水体中TN显著高于鱼单养模式(P<0.001)。与春季相反,在夏季莲鱼复合种养模式水体中TP含量显著高于鱼单养模式(P<0.05)。秋季莲鱼复合种养模式的TN含量显著高于鱼单养模式(P<0.05),TP含量在各模式中没有显著差异(P<0.05),NO3-N含量显著低于鱼单养和莲单种模式(P<0.001),NH4-N含量显著低于鱼单养模式(P<0.001),溶氧显著高于鱼单养和莲单种模式(P<0.001;图 5)。
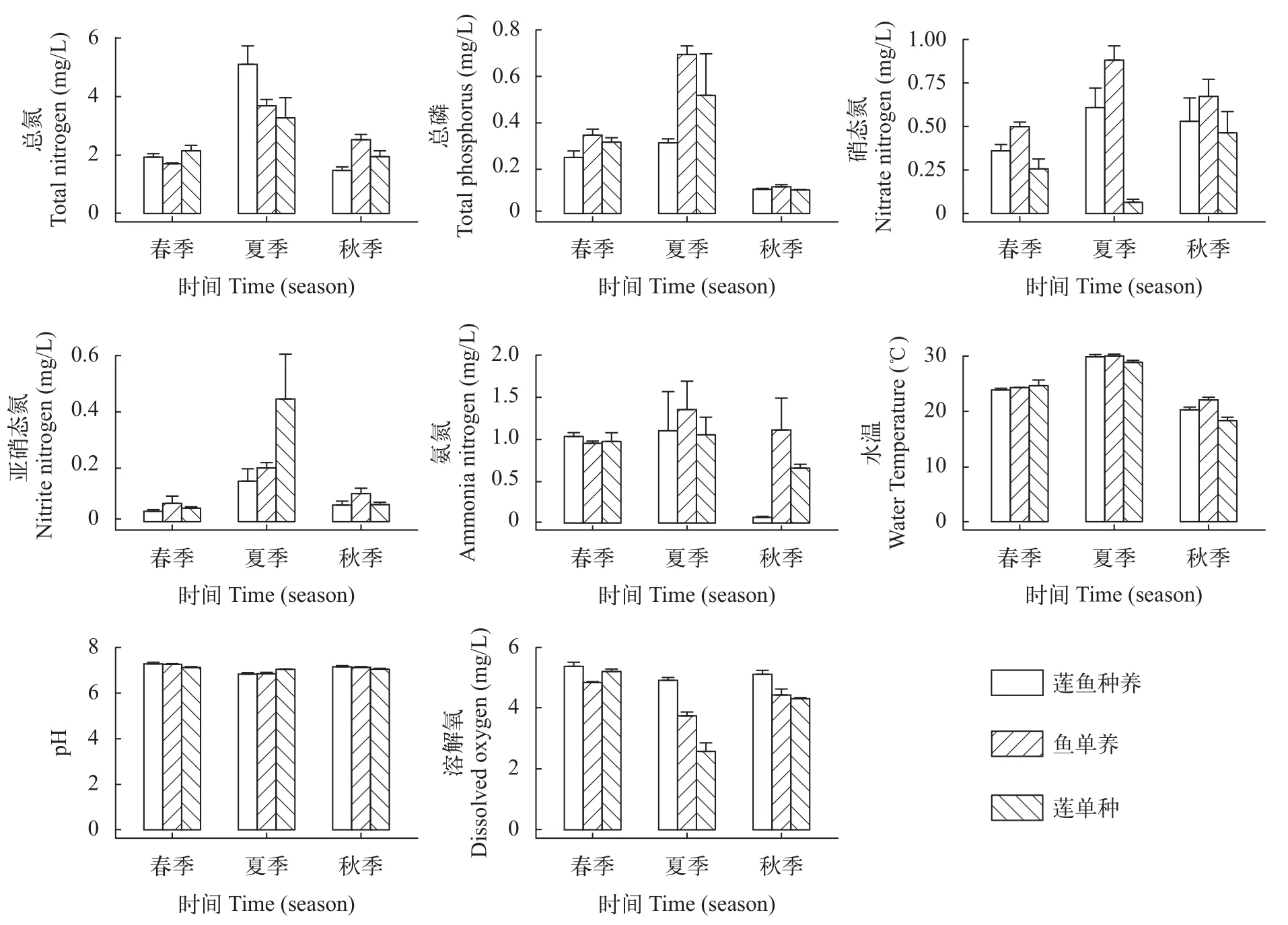
图5 莲鱼种养、莲单种和鱼单养模式水体中不同季节水体理化因子Fig.5 Seasonal variation of environmental factors in the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
2.4 环境因子和浮游植物群落的关系
本研究中莲鱼复合种养模式水体中环境因子和浮游植物群落的关系排序轴中梯度长度的最大值为2.142,使用RDA模型结果更为准确;鱼单养和莲单种模式中排序轴中梯度长度的最大值分别为3.775和3.368,既可使用CCA模型又可使用RDA模型进一步分析。为确保分析方法的统一性,本研究均采用RDA模型进行分析。进行初步RDA分析后,使用ordistep函数进行前向选择,保留显著影响浮游植物群落结构特征变化的环境因子(P>0.05),并进一步构建主要环境驱动因子和浮游植物群落结构特征相互关系的简约模型。随后对模型的R2进行矫正,并对约束轴进行蒙特卡罗置换检验,确保RDA模型、第一约束轴和第二约束轴均显著(P<0.05)。简约模型显示,各季节样方水体中浮游植物群落具有明显差异(图6)。其中,莲鱼复合种养模式浮游植物群落的主要驱动因子为WT和DO,对浮游植物群落特征的解释率分别为28.48%和24.36%;鱼单养模式浮游植物群落的主要驱动因子为TP和DO,对浮游植物群落特征的解释率分别为39.47%和33.52%;莲单种模式游植物群落的主要驱动因子为WT、NO3-N和DO;对浮游植物群落特征的解释率分别为45.56%、17.37%和5.06%。
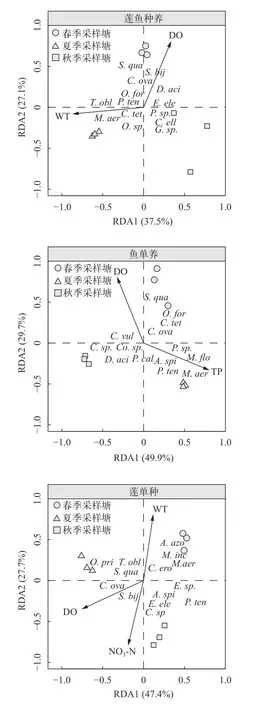
图6 莲鱼种养、莲单种、鱼单养模式水体中浮游植物群落和环境因子的冗余分析Fig.6 RDA analysis of phytoplankton communities and environmental factors of the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
2.5 经济效益和生态足迹
莲鱼种养每公顷可产生经济效益2.52万元,每公顷利润为鱼单养>莲鱼种养>莲单种(表3),而每公顷莲鱼模式的总生态足迹为11.27 hm2,每公顷莲单种的总生态足迹为18.91 hm2,每公顷鱼单养模式的总生态足迹为19.54 hm2,鱼单养和莲单种带来的生态负荷分别为莲鱼复合种养模式的1.73倍和1.68倍。此外,莲鱼复合种养模式单位利润的生态足迹量为4.48 hm2,莲单种模式单位利润的生态足迹量为13.48 hm2,鱼单养模式单位利润的生态足迹量为5.54 hm2。即莲鱼模式每产生1万元的经济效益需要该地区4.48 hm2的生产土地来提供其所需投入品或消纳其产生的废物,鱼单养模式则需要5.54 hm2生产土地,莲单种模式则需要13.48 hm2生产土地。

表3 莲鱼种养、鱼单养、莲单种模式的生态足迹Tab.3 Ecological footprint of the integrated lotus-fish culture system,lotus monoculture system and fish monoculture system
3 讨论
3.1 水体理化因子和浮游植物群落结构特征
本研究发现莲鱼复合种养模式及其单种、单养模式水体中TN、TP、NH4-N、NO3-N、NO2-N和WT在研究期间均呈现出先升后降趋势,而pH和DO则呈现出先降后升趋势。在本研究中鱼单养模式和莲单作模式水体中溶解氧在调查期间,尤其是夏季相比莲鱼种养模式较低,表明水体内有机负荷相对莲鱼种养模式较高[33];而莲单种模式水体中溶氧在夏季显著低于其他模式则更可能与莲叶的过度生长及莲叶和浮游植物对阳光的竞争有关。氮、磷等营养物质是浮游植物生长繁殖的物质基础,也是在水质监测评价中重点关注的指标[34]。在本研究中,夏季莲鱼种养模式水体中TN含量达到高峰,且显著高于单养/种模式,可能是由于共作过程中的施肥和投饵共同导致的。而在莲鱼种养模式水体中TP含量始终低于鱼单养模式,这可能与莲生长期对磷元素的旺盛需求有关。
浮游植物群落存在明显的季节演替现象,研究期间各模式水体中浮游植物的演替规律与Sommer等[35]提出的演替模式基本一致。在各模式水体中,隐藻常在春夏季形成优势种,这可能与隐藻能通过鞭毛调节自身位置并适应低光条件有关[36]。而蓝藻门在各个季节均存在优势种,与蓝藻具有的可调节细胞浮力的气泡有关,这种能力使得蓝藻可以适应不同季节并广泛分布[37]。秋冬季优势度较高的异极藻和小环形藻均属于硅藻门,而这些藻类的最适温度一般低于25℃。在研究期间,春季鱼单养和莲鱼种养模式水体中浮游植物密度均显著高于莲单作模式水体中浮游植物密度,一方面与鲫对浮游植物的摄食压力弱有关[38],另一方面可能与鲫作为底层鱼类,可通过生物扰动作用促进营养物质从底泥中释放,从而引起水体中浮游植物生物量增加有关[39]。此外,复合种养模式各个季节的香农威纳指数均比单养/种模式高,说明种养模式中浮游植物群落多样性高;而复合种养模式中浮游植物群落多样性的增加,有利于形成更复杂的食物网结构,提升模式的稳定性和生产力[25]。而在鱼单养模式中,夏季仅有蓝藻门的藻类形成优势种,对水体中浮游植物密度的贡献最大,优势度较高的种类将占据更多生态位[40],从而导致群落结构稳定性下降,增加发生水华的风险。
3.2 水体理化因子对浮游植物群落结构特征的影响
浮游植物群落结构受外界环境中温度、营养盐等环境因子的驱动,始终处于动态变化过程之中[41]。温度和营养盐是影响浮游植物生长的两大重要环境因素。水温作为显著影响莲鱼种养和莲单种模式中浮游植物群落特征的共同环境因子,一方面影响浮游植物的新陈代谢速率,另一方面也通过影响各类营养物质在水体中的溶解度影响浮游植物生长和繁殖[42]。通常浮游植物密度会随着温度上升而上升,且较高的温度有利于绿藻门和蓝藻门的生长和繁殖[43]。由于蓝藻对温度的耐受性高于其他藻类,在高温下通常以快速生长的r对策获得竞争优势[44],这与本研究中蓝藻门中的铜绿微囊藻、水华微囊藻、不定微囊藻和颤藻在高温季节的高细胞密度结果一致。众多研究表明水体中氮、磷等营养盐对藻类的生长均具有限制作用,氮磷比(TN/TP)通常用来判断氮或磷是否为限制性因素,当氮磷比>16时,则认为磷是限制性因素;反之,则认为氮是限制藻类生长的主要因素[45,46]。在本研究中鱼单养和莲单种模式中秋季氮磷比均大于16,莲鱼模式中秋季氮磷比接近16(13.36—15.43),秋季浮游植物密度较低,水体均处于极贫营养状态。冗余分析表明,氮、磷营养盐并不是影响莲鱼种养模式水体中浮游植物群落特征的主要影响因素;而水体中总磷含量则是影响鱼单养模式水体中浮游植物群落特征的主要影响因素,且与微囊藻等蓝藻的高密度密切正相关。而在莲单种模式中,浮游植物还受到硝酸盐的调控。冗余分析结果显示,铜绿微囊藻、不定微囊藻与水体中NO3-N含量呈负相关,这可能与微囊藻细胞能将NO3-N转化为其他形式的氮排出细胞外有关[47]。在鱼单养模式中,蓝藻类中的假鱼腥藻、铜绿微囊藻、水华微囊藻、小席藻和TP高度正相关,且夏季水体中总磷含量显著高于莲鱼种养模式。而随着温度上升底质磷的释放可导致水体中可利用的磷酸盐增多[48],导致上述浮游植物生物量增加。
3.3 水体富营养化和污染状况
浮游植物群落结构特征与水体环境密切相关,且能对水体变化做出快速响应,主要表现在种类组成、细胞密度和优势种的变化。因此,浮游植物群落结构和多样性的变化常用于水体营养状态、污染程度评价[49]。根据浮游植物密度[50],春季莲鱼种养模式水体属于贫中营养,鱼单养和莲单种模式均属于贫营养状态;夏季莲鱼种养模式和莲单作模式均处于中营养状态,鱼单养模式处于富营养状态;秋季3种水体均处于极贫营养状态。为避免单一多样性指数评价的局限性,本研究选用Shannon-Wiwner多样性指数和Pielou均匀度指数共同进行评价。根据Shannon-Wiwner多样性指数评价标准[50],莲鱼种养模式3个季节水体均处于和均匀度指数得到的莲鱼种养模式各季节均处于轻污染-无污染状态,鱼单养各季节均处于β-中污状态,莲单种模式除秋季水体属于轻污染-无污染状态外,其他季节均处于β-中污状态。根据Pielou均匀度指数,莲鱼种养模式3个季节水体均处于和均匀度指数得到的莲鱼种养模式各季节均处于轻污染-无污染状态,鱼单养和莲单种模式除夏季属于β-中污状态,其他季节均属于轻污染-无污染状态。在本研究中,较高的浮游植物多样性指数与水体的富营养化程度并不完全一致,与前人研究结果一致[51,52]。一方面可能与水体受到干扰导致其生物多样性和水体的水质情况有悖有关,另一方面本研究中未对浮游植物进行功能群分类,根据生物多样性指数得到的水质评价结果可能比实际偏高。虽然,浮游植物密度和两种指数的评价结果显示,各模式中水体富营养化程度不完全一致,但总体而言各模式中水体富营养化程度的季节变化均为夏季>春季>秋季,且莲鱼种养模式的水质优于单养/单种模式,可能与莲鱼种养模式中的莲作为高等水生植物具有一定的抑藻效应[53],能抑制藻类优势种的生长、扩散有关。
3.4 莲鱼种养模式的生态成本
生态足迹模型[27,28]从1999年引入我国后,被广泛应用于国家、地区、社区、学校、行业乃至个人的生态负荷计算,为度量可持续发展提供了一个“公平秤”,并逐渐开始应用在渔业和种养业的可持续发展衡量中,进而为政策制定提供参考依据。如通过计算渔业集约化养殖模式的生态足迹为具体生产管理提供理论基础,通过计算大菱鲆流水养殖和循环水养殖养殖场生态足迹为中国大菱鲆循环水养殖提供参考建议等[54,55]。在本研究中莲鱼种养每公顷可产生经济效益2.52万元,平均每公顷利润低于鱼单养模式的3.53万元,高于莲单种模式的1.40万元,经济效益适中;然而莲鱼复合种养模式每公顷所产生的生态足迹为11.27 hm2,其用来提供其所需投入品或消纳其产生的废物的生产土地远低于单养/种模式,表明复合系种养统具有更好的可持续性,生态效益和经济效益更高,符合循环经济理念。在井冈山地区,太空莲虽然被广泛种植,然而莲子和莲藕的产量极易受天气影响,给种莲的农户们带来了更多的成本风险和收益不确定性。而莲鱼种养则提供了一种更为经济高效、生态环保的可持续发展模式。
