山东近海三种底层鱼类的摄食生态与食物竞争
2022-08-04韩方成李凡冯媛刘利军钮春香许叶鹏任一平薛莹
韩方成 李凡 冯媛 刘利军 钮春香 许叶鹏 任一平, 薛莹,
(1.中国海洋大学水产学院,青岛 266003;2.山东省海洋资源与环境研究院,山东省海洋生态修复重点实验室,烟台 264006;3.青岛海洋科学与技术试点国家实验室,海洋渔业科学与食物产出过程功能实验室,青岛 266237;4.海州湾渔业生态系统教育部野外科学观测研究站,青岛 266003)
鱼类摄食生态研究是现代鱼类生态学的基础理论之一,同时也是解析海洋生态系统结构和功能的基础。开展鱼类摄食生态与食物竞争的研究,是深入理解鱼类捕食行为和饵料选择性的关键所在,也是了解鱼类群落结构和生态系统功能的重要途径[1]。
得益于独特的地理环境和水文因素,山东半岛近岸海域是黄海多种渔业生物的产卵场、索饵场和传统渔场[2]。近年来,由于高强度的捕捞压力,导致渔业资源严重衰退,渔获物中传统经济种类比例也明显下降,群落结构发生了较大的变化[3]。根据本课题组2016—2017年在山东近海进行的季节性渔业资源底拖网调查,小眼绿鳍鱼(Chelidonichthys spinosus)、黄鮟鱇(Lophius litulon)高眼鲽(Cleisthenes herzensteini)是山东近海重要的优势底层鱼类,它们的生态位相近,分布区域有所重叠,因而有可能存在激烈的种间竞争。国内学者对这3种鱼类的研究主要集中在资源密度、生长和摄食等方面[4—7],而有关其种间食物竞争的研究较少,亟待开展相关研究。
为此,本研究应用分类树和生态位重叠指数法等,研究了山东近海3种主要底层鱼类的摄食生态及其营养-空间二维生态位的重叠情况,通过典范对应分析研究环境因素对其摄食的影响,旨在深入了解种间竞争对鱼类时空分布、种群数量变动以及海洋生态系统稳定性的影响[5],并为该海域渔业资源的可持续利用和科学管理提供基础资料[6]。
1 材料与方法
1.1 样品采集
样品采自2016—2017年在山东近海进行的四个季节的底拖网调查(图1)。利用功率为220 kW的单拖网渔船以2 kn的拖速开展调查,每站拖网时间不少于1h,拖网时网口宽度约为25 m,囊网网目尺寸为17 mm[7]。严格按照《海洋调查规范》[8]进行样品的采集与分析。本调查共采集3种鱼类样品1630尾,其中440尾为空胃,样品数、平均体长及摄食率如表 1所示。
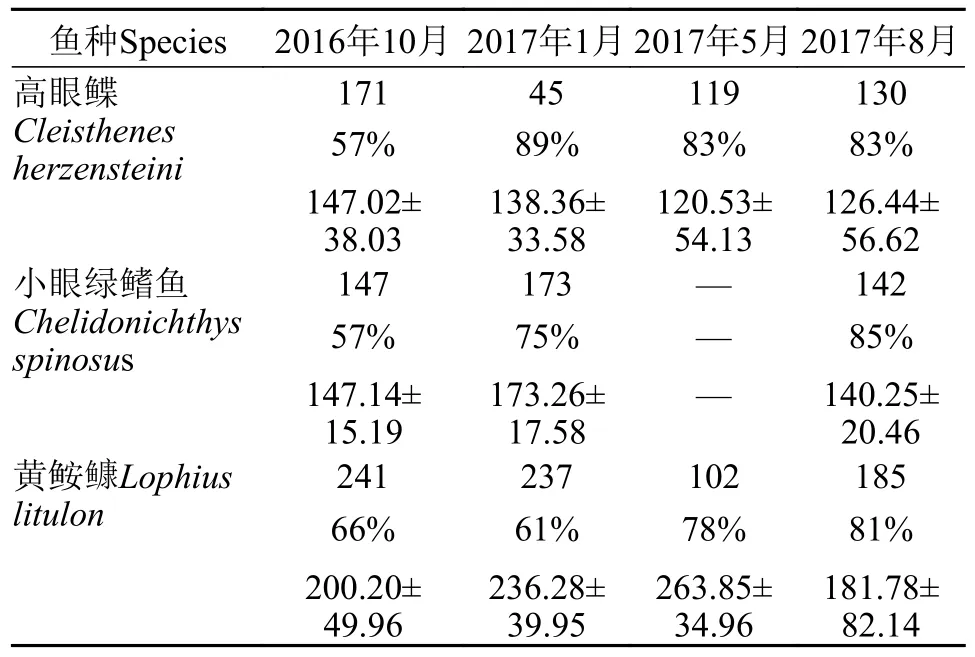
表1 山东近海3种鱼类的胃含物样品数、摄食率和平均体长Tab.1 Sample number,feeding rate and mean size of three fish species in the coastal waters of Shandong Province (mm)

图1 山东近海调查站位图Fig.1 Sampling stations in the coastal waters of Shandong Province
1.2 胃含物分析
鱼类样品经生物学测定后,取出消化道并立即速冻保存[9]。将胃含物样品解冻后,用吸水纸吸去水分后,再在双筒解剖镜下鉴定出饵料生物的最低分类单元,并分别计数和称重,称量所用天平精确到0.001 g[10]。
1.3 数据分析
通过绘制累积饵料生物曲线(Cumulative prey curves)来确定胃含物样品数量是否充足[11]。随着样品量的增加,饵料生物种类数的变化会变小,当累积饵料生物曲线接近一条渐近线时,表明已分析足够数量的样品来反映其摄食状况[7]。
相对重要性指数(IRI)[12]综合了质量百分比(W%)、个数百分比(N%)和出现频率(F%)3方面的信息,是描述特定饵料种类重要性的综合性指标,本研究采用相对重要性指数百分比(IRI%)评价各饵料生物的重要性。计算公式如下:

应用分类树和相似性分析(ANOSIM)比较摄食的种间差异。分类树是一种递归算法,每次只评估一个解释变量。此过程在变量中设置阈值,该阈值解释每个实例中方差的最大比例。重复这一过程,直到所解释的方差没有进一步增加,或者剩下的子组是同质的[13]。我们将物种种类作为响应变量,将饵料生物的百分比作为解释变量,通过分析,评估其食物组成是否存在差异。ANOSIM是一种多元统计分析方法[14],它通过评估距离矩阵中的组间差异是否大于组内差异,来评估组间(即物种之间)的差异[15]。
典范对应分析(CCA)是一种非线性多元直接梯度分析方法,可以结合多个环境因子一起分析,信息量大且结果直观明显[16],可以更好地反映鱼类摄食与环境因子的关系。应用典范对应分析研究影响摄食的主要因素,利用Bray-Curtis相似性指数作为矩阵计算的基础,通过从相同的输入数据确定食物组成数据和响应变量之间的关联度。选取体长、体重、季节和地理位置(纬度、经度)作为解释变量。按个数百分比(N%)进行计算。通过排列(9999次模拟)确定了整个模型、每个坐标轴和每个相应变量的重要性,并沿着前两个坐标轴构建了相应变量和饵料组成的双线图。
在生态学研究中,用于描述生态位宽度的指数较多,但Shannon-Wiener指数凭借其对稀有饵料生物比较敏感[17]的特性,而被广泛地应用于鱼类生态位宽度的研究。因此本研究采用Shannon-Wiener指数(H′)来计算3种鱼类的营养和空间生态位宽度,应用Pianka生态位重叠指数(Ojk)来研究其生态位重叠[7]。由于不同种类及同种类个体之间差异较大,使用质量百分比更能表示种类间能量的分布[7]。
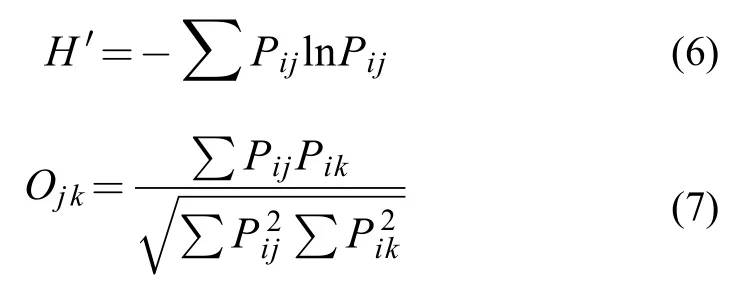
式中,Pij在计算营养生态位时,为在捕食者j食物组成中饵料生物i所占的质量百分比;在计算空间生态位时,捕食者i在第j个站位总生物量中所占的比例。Ojk的变化范围为0—1,值越大,说明重叠程度越高,Oij>0.3视为有重叠,Oij>0.6视为重叠显著[18]。各个站位的鱼类生物量以拖速2 kn和拖网时间1h为基准进行标准化处理。
由于营养和空间资源相互独立,因此可以使用鱼类营养、空间生态位重叠指数的乘积来表示鱼类之间的营养-空间二维生态位重叠指数,作为评价鱼类种间竞争程度的指标[19]。
2 结果
2.1 食物组成
本研究发现,山东近海3种鱼类的累积饵料生物曲线都达到或接近一条渐近线(图2),说明胃含物样品数量足以反映其摄食情况。
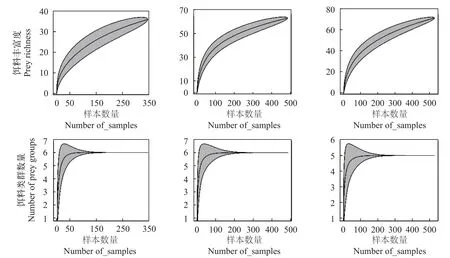
图2 山东近海3种鱼类的累积饵料生物曲线Fig.2 Cumulative prey curves of three fish species in the coastal waters of Shandong Province
根据IRI%的值(表2),高眼鲽主要以虾类(91.41%)为食,此外也摄食一定数量的端足类(1.52%)、棘皮动物(2.67%)和鱼类(3.06%),而对蟹类、糠虾等摄食较少。脊腹褐虾(Crangon affinis;60.95%)和中华安乐虾(Eualus sinensis;23.69%)为其主要的饵料生物(表2)。
小眼绿鳍鱼主要以虾类(94.59%)为食,同时也摄食一定量的端足类(1.61%)、鱼类(2.28%)和蟹类(1.03%)。在虾类中,细螯虾(L.gracilis;43.37%)、脊腹褐虾(18.55%)和戴氏赤虾(M.dalei;20.11%)所占比例较大,为优势饵料(表2)。
黄鮟鱇主要以鱼类(85.91%)和虾类(12.83%)为食,其他饵料则摄食的比较少。脊腹褐虾(30.70%)和方氏云鳚(Enedrias fangi;34.10%)为其优势饵料(表2)。
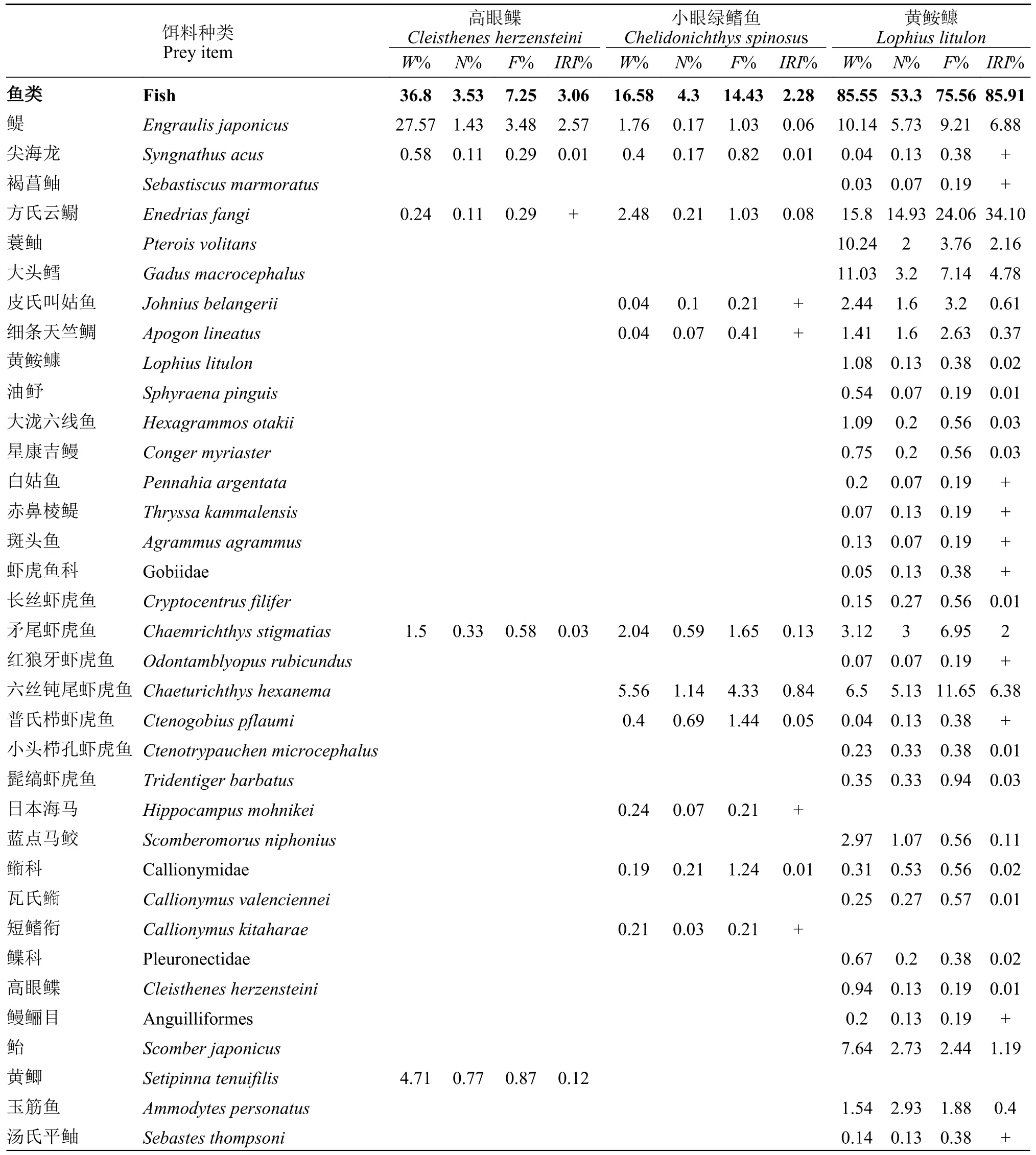
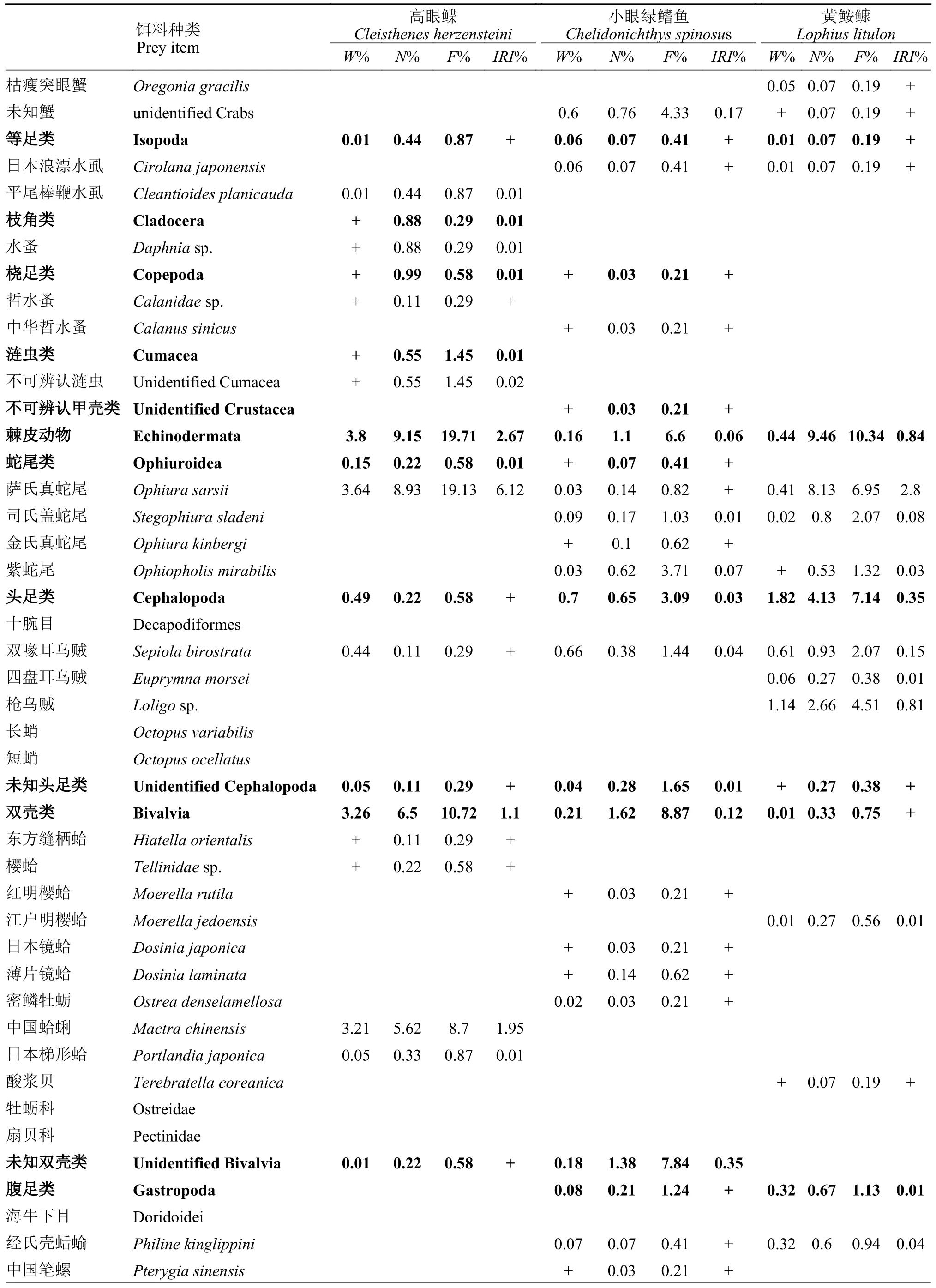
表2 山东近海3种底层鱼类的食物组成Tab.2 Food composition of three demersal fish species in the coastal waters of Shandong Province

续表 2
续表 2

续表 2
分类树表明,这3种鱼类的食物组成存在较大差异(图3),黄鮟鱇摄食鱼类的比例最高,小眼绿鳍鱼则摄食比较多的虾类,而高眼鲽则主要以软体动物为食。ANOSIM分析也表明,这3种鱼类的摄食存在极显著差异(R=0.356,P<0.01)。
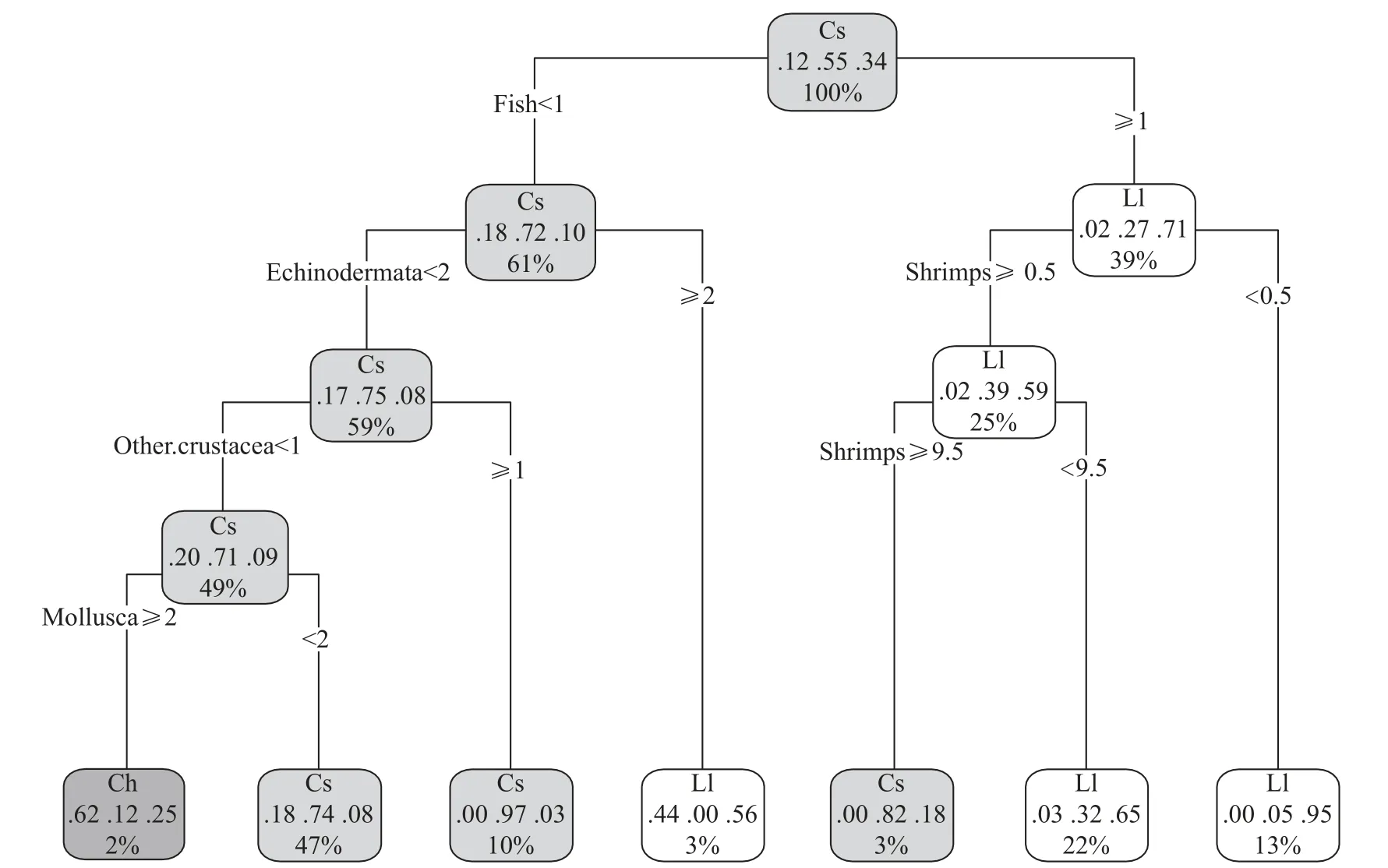
图3 山东近海3种鱼类食物组成的分类树Fig.3 Classification tree of the diet composition of three fish species in the coastal waters of Shandong Province
2.2 影响摄食的因素
CCA分析表明,高眼鲽的摄食习性主要与体长有关。在模型中,CCA1(45.49%)和CCA2(32.66%)解释了最大的摄食差异(图4)。随着体长的增加,高眼鲽会摄食更多的鱼类和虾类,而摄食其他甲壳类的比例有所减少;随着经度的增加,高眼鲽摄食更多的软体动物和棘皮动物。
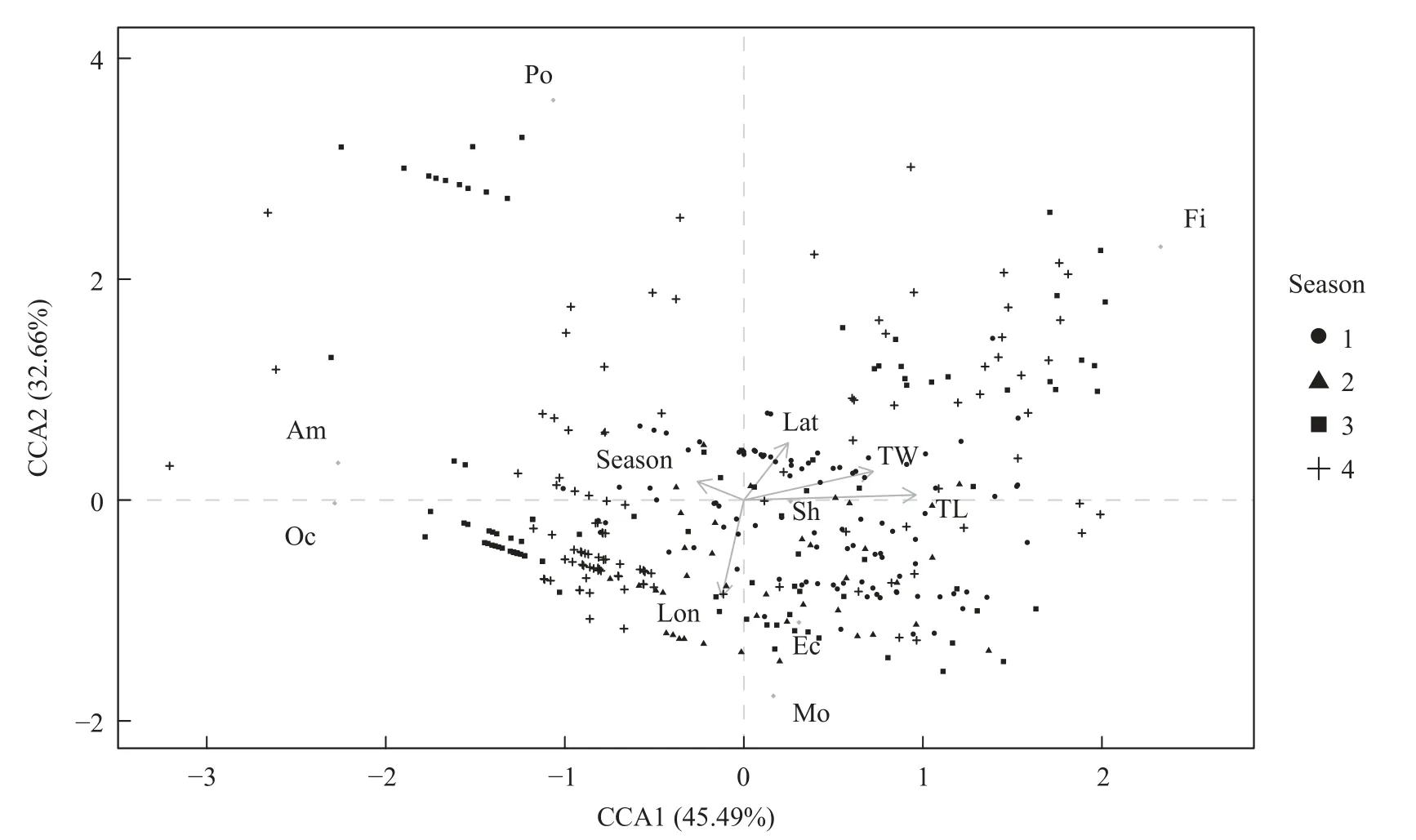
图4 高眼鲽摄食习性与影响因素之间关系的CCA分析图Fig.4 CCA biplot of the relationships between response variables and feeding habits
CCA分析表明,小眼绿鳍鱼的摄食习性主要与季节变化有关(图5)。在模型中,CCA1(61.1%)和CCA2(20.43%)中解释了最大的摄食变异性,并且CCA1解释的程度要明显高于CCA2。在CCA2中,小眼绿鳍鱼摄食鱼类、虾类的比例与季节变化有关。
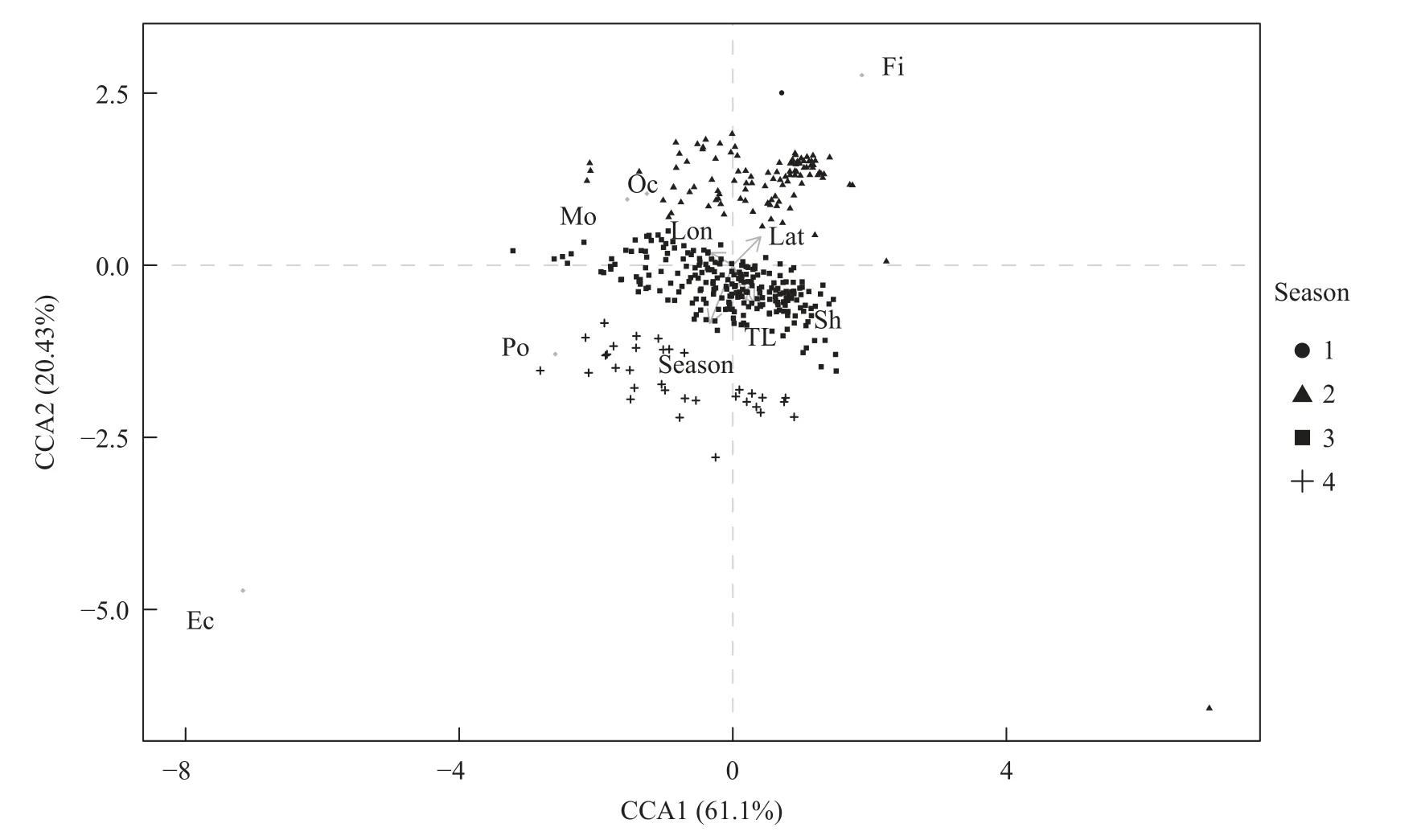
图5 小眼绿鳍鱼摄食习性与影响因素之间关系的CCA分析图Fig.5 CCA biplot of the relationships between response variables and feeding habits
CCA分析表示,黄鮟鱇的摄食主要受空间分布的影响(图6)。在模型中,CCA1(63.23%)和CCA2(29.76%)中解释了最大的摄食变异性。CCA1表示黄鮟鱇摄食虾类和鱼类的比例随着空间位置的变化而变化。模型显示,高纬度的黄鮟鱇摄食更多的虾类,而随着经度的增加,黄鮟鱇会摄食更多的鱼类。
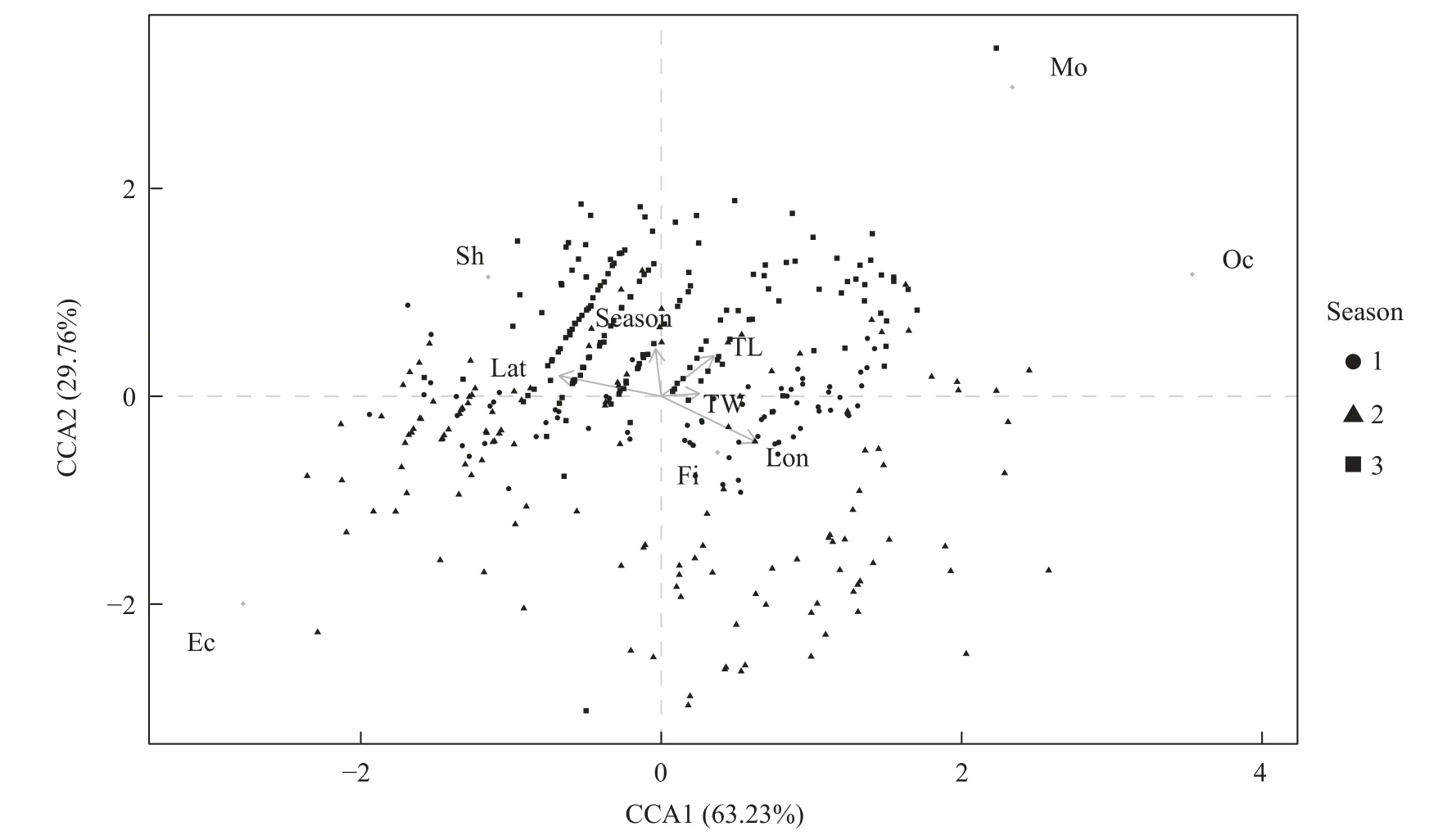
图6 黄鮟鱇摄食习性与影响因素之间关系的CCA分析图Fig.6 CCA biplot of the relationships between response variables and feeding habits
2.3 营养和空间生态位竞争
如表 3所示,高眼鲽的营养生态位宽度最低(1.65),黄鮟鱇的营养生态位宽度最高(3.10)。对于空间生态位,小眼绿鳍鱼和黄鮟鱇的空间生态位都较大,均大于3.5,而高眼鲽的空间生态位都较低(2.95)。
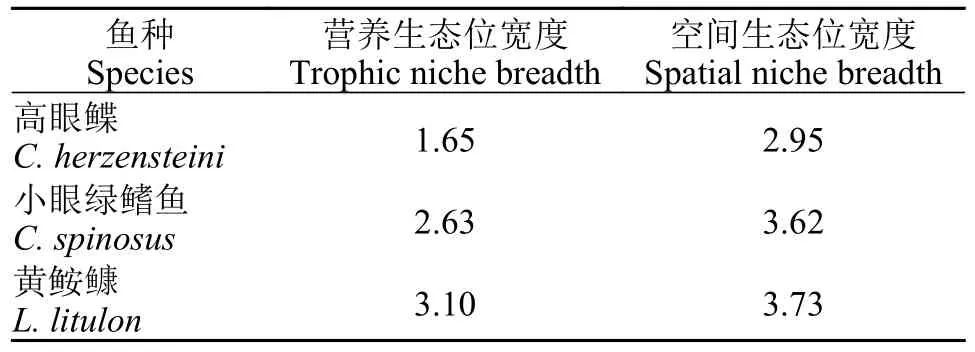
表3 山东近海3种鱼类的营养和空间生态位宽度Tab.3 Trophic and spatial niche breadth of three fish species in the coastal waters of Shandong Province
对于营养生态位重叠指数(表4),高眼鲽与小眼绿鳍鱼(0.858)具有最高的营养生态位重叠(0.858),而小眼绿鳍鱼与黄鮟鱇的营养生态位重叠最小(0.399)。相较于营养生态位重叠,3种鱼类的空间生态位重叠并不明显(表4),高眼鲽与黄鮟鱇具有最高的空间生态位重叠(0.478),而小眼绿鳍鱼与黄鮟鱇的空间生态位重叠则很小(0.115)。
而对于营养-空间二维生态位重叠(表4),虽然高眼鲽与小眼绿鳍鱼具有最高的营养生态重叠,但由于其空间生态位重叠较小,所以导致其营养-空间二维生态位重叠较小;相反,虽然高眼鲽和黄鮟鱇的营养生态位重叠不是很高,但是其较大空间生态位重叠导致其营养-空间二维生态位重叠最大(表4)。
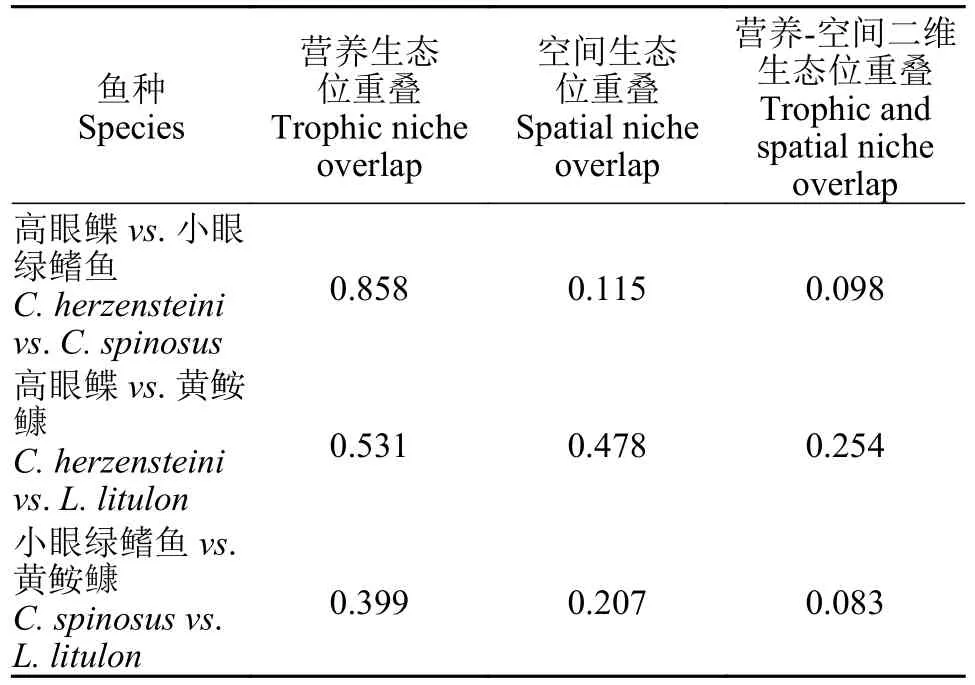
表4 山东近海3种鱼类的营养和空间生态位重叠Tab.4 Trophic and spatial niche overlap index of three fish species in the coastal waters of Shandong Province
3 讨论
3.1 摄食习性
本研究发现,山东近海高眼鲽属于底栖动物食性,主要摄食虾类,以脊腹褐虾和中华安乐虾为优势饵料。窦硕增等[20]研究表明,渤海的高眼鲽主要以鲜明鼓虾(Alpheus distinguendus)、口虾蛄(Oratosquilla oratoria)、日本鼓虾(Alpheus japonicus)及火枪乌贼(Loligo beka)为食,说明不同海域高眼鲽的食物组成存在差异。山东近海小眼绿鳍鱼摄食的饵料生物达60余种,属于底栖动物食性。根据以往的研究,渤海小眼绿鳍鱼以脊腹褐虾、鹰爪虾(Trachypenaeus curvirostris)、口虾蛄、细螯虾及六丝钝尾虾虎鱼(Chaeturichthys hexanema)为优势饵料[21];海州湾的小眼绿鳍鱼主要摄食细螯虾、戴氏赤虾、疣背宽额虾(Latreutes planirostris)等[22]。综合看来,小眼绿鳍鱼在不同海区都主要以虾类为食。山东近海黄鮟鱇摄食的饵料生物达70余种,主要以鱼类为主,但不同季节的优势饵料却有差异。薛莹等[23]的研究表明,北黄海秋季黄鮟鱇的优势饵料生物是矛尾虾虎鱼(Chaemrichthys stigmatias)和脊腹褐虾,与本研究的优势饵料(方氏云鳚和脊腹褐虾)存在差异,这表明鱼类摄食的优势饵料生物会随自然海域中饵料生物组成或者数量的变化而发生变化[24]。本研究发现,山东近海这3种鱼类虽然都属于底栖动物食性,但是其优势饵料的种类和所占比例不同,在一定程度上减缓了对饵料生物的竞争,有利于种间共存。
3.2 影响摄食的因素
本研究表明,高眼鲽的摄食与体长有显著的关系,随着体长的增加,高眼鲽会摄食更多的鱼类和虾类,而摄食其他甲壳类的比例有所减少。张波等[25]发现,不同体长组高眼鲽摄食的食物种类有一定差异,体长<119 mm的主要以磷虾类和底层虾类为食;当体长增大时,主要摄食底层虾类和鱼类,虾类所占比例减小,与本研究的结论相吻合。鱼类的生长发育往往伴随着口裂大小的增加,游泳速度的加快,从而导致其捕食大型饵料生物的能力也在不断地增加,因此个体较大的鱼类捕食饵料鱼类的比例也较高[26]。本研究还发现,山东近海高眼鲽摄食的饵料生物达30余种,但是每个季节摄食的饵料生物种类只有10余种,说明高眼鲽的摄食习性会随着季节变化而发生较大的变化,这可能与山东近海饵料生物的季节变化有关[27—28]。此外,高眼鲽的体长组成存在显著的季节差异(P<0.05),这也是导致其摄食习性存在季节变化的原因之一。
本研究还发现,黄鮟鱇的摄食与其空间分布有着密切的联系。以往的研究表明[23],体长、性别和性腺成熟度对北黄海黄鮟鱇的食物组成没有显著影响,水深、经度和纬度是影响北黄海黄鮟鱇食物组成的关键因素,与本研究的结论相吻合。饵料生物受到栖息环境和生活习性等因素的影响,其分布存在较大的空间异质性。鱼类总是倾向于摄食该海域可获得性较高的饵料生物,因此就导致了同种鱼类摄食的空间差异[29]。
3.3 营养和空间生态位重叠
生物对环境的适应能力和耐受程度主要通过生态位宽度来衡量[30]。在本研究的3种鱼中,高眼鲽的营养生态位宽度最低,说明高眼鲽所摄食的饵料种类数最少,对饵料的利用范围最小,其对饵料资源的利用能力相对较弱。空间生态位宽度是对物种空间分布范围的反映[31]。本研究发现,黄鮟鱇的空间生态位宽度最高,说明其适应环境的能力相对较高,这也与其在山东近海的分布范围较广相吻合。而其余两种鱼出现的站位相对较少,因此它们的空间生态位宽度也较小。
生态位重叠指数通过反映物种间对资源利用的相似程度,来反映物种间潜在的竞争关系[32]。其中,营养生态位重叠指数反映了物种间食物组成的相似程度[7]。在本研究中,高眼鲽和小眼绿鳍鱼的营养生态位重叠最高,说明二者的食物组成较为相似。空间生态位重叠指数反映的是不同物种在空间分布上的重叠程度[9]。本研究发现,山东近海这3种鱼类的空间生态位重叠较低,在一定程度上能够缓和因营养生态位重叠过高而导致的种间竞争。
营养-空间二维生态位重叠指数结合了营养生态位和空间生态位重叠指数的特征,反映了物种之间对食物和空间资源利用的相似程度。物种之间,高的营养-空间二维生态位重叠指数一般对应着高的空间生态位重叠指数和高的营养生态位重叠指数,任一维度的重叠指数偏低都会导致营养-空间二维生态位重叠指数降低[7]。在资源有限的环境中,鱼类需要通过对营养和空间等生态位利用的平衡来实现共存[33]。本研究表明,山东近海这3种鱼类都没有较为明显的营养-空间二维生态位重叠,虽然3种鱼类的营养生态位重叠都较为显著,但是空间生态位重叠较小,导致它们的营养-空间二维生态位重叠也较小。这说明3种鱼类通过不同程度的空间生态位分化,在一定程度上缓解了因选择相似的饵料生物而产生的种间竞争。
