毛竹林和阔叶林凋落物互置对土壤氮矿化的影响及微生物贡献
2022-08-03卢小妮陈露雨李永春胡晓嫣李永夫俞叶飞
卢小妮,陈露雨,李永春,*,方 韬,胡晓嫣,李永夫,俞叶飞
1 浙江农林大学环境与资源学院/浙江省森林生态系统碳循环与固碳重点实验室/亚热带森林培育国家重点实验室, 杭州 311300 2 浙江省大盘山国家级自然保护区管理局, 磐安 322300
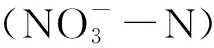
微生物是连接植物与土壤的纽带,在土壤生物地球化学循环中起着关键作用[11];添加凋落物的量决定了土壤微生物群落的结构及底物利用模式[12]。高质量的凋落物含有较多相对易降解的化学物质,因此易于被大多数土壤微生物群落降解;而低质量的凋落物往往含有较多不易被微生物快速降解的高抗性或有毒成分,所以具有较强的主场效应[13]。已有研究表明,土壤细菌和真菌群落相对丰度与凋落物初始木质素与氮的比率及碳氮比有关[14]。土壤微生物作为土壤有机质分解和氮矿化的总工程师,细菌和真菌参与了土壤净氨化和净硝化过程[15]。使用放线菌酮和链霉素选择性抑制真菌和细菌群落,可以更好地探明真菌和细菌在氮素矿化中的贡献[16]。添加放线菌酮或链霉素可促进氮素矿化[17],而添加放线菌酮会使旱雀麦和三齿蒿土壤中总氮矿化降低[18],表明真菌在土壤总氮矿化中发挥主要作用。采用抗生素解析真菌和细菌群落在氮素矿化中的贡献已有报道,但结合抗生素抑制和凋落物输入质量变化探究真菌和细菌对土壤氮素矿化的贡献尚鲜见报道。
毛竹(Phyllostachysedulis)主要分布在我国亚热带地区,由于其快速生长、强大的繁殖和再生能力[19],可以快速扩张到邻近的天然林[20]。由于氮是植物生长重要的限制性养分,入侵植物如何影响土壤氮素水平及有效性是理解入侵机制的重要环节[21]。毛竹入侵阔叶林使土壤的净氮矿化增加[22],且毛竹苗在铵态氮为主的氮源时生长更占优势[23]。此外,毛竹林土壤氮矿化速率与凋落物数量呈正相关,与凋落物碳氮比呈负相关[20]。与凋落物降解存在主场效应一样,土壤有机质分解的“激发效应”也存在主场效应,表明特定的凋落物降解群落驱动的激发效应[24],可能通过增强诱导“激发效应”从而影响土壤氮矿化。为此,本研究选取次生常绿阔叶林和毛竹入侵后形成的毛竹纯林,分别代表未被入侵和受到重度入侵的植被类型,对林地凋落物进行保留、去除和置换处理;并采集处理小区内的土壤样品,进行室内氮素矿化培养试验测定土壤净氮矿化速率,结合添加抗生素解析真菌和细菌群落对土壤氮素矿化的相对贡献。本研究拟回答以下科学问题:(1)在毛竹入侵次生阔叶林过程中,输入凋落物化学性质和降解生境的改变如何影响土壤氮矿化速率?(2)前述凋落物化学性质和降解生境改变后,真菌和细菌群落对土壤氮素矿化的相对贡献是否改变?本研究旨在探明毛竹林和阔叶林地上凋落物对土壤氮素矿化的影响及微生物群落的贡献,以期为阐明毛竹入侵生境不同类型凋落物输入调控土壤氮矿化过程提供理论依据。
1 研究区域与研究方法
1.1 研究区域概况
实验区域位于浙江省杭州市临安区天目山国家级自然保护区(30°18′30″—30°21′37″N, 119°24′11″—119°27′11″E),天目山保护区是中国亚热带生态系统中植物物种最丰富的区域之一。本研究区域气候为亚热带季风气候,年均气温为8.8—14.8℃,年均降水量为1390—1870 mm,平均无霜期为209—235 d。亚热带常绿阔叶林为本区域的地带性植被,区域内阔叶林优势树种主要为青冈(Cyclobalanopsisglauca)、木荷(Schimasuperba)和苦槠(Castanopsissclerophyll)等,自20世纪70年代以来经历了毛竹入侵[25]。在毛竹入侵典型区域,根据毛竹扩张水平方向和海拔由低到高,依次划分为入侵后形成的纯毛竹林、毛竹-阔叶混交林和尚未被毛竹入侵的次生阔叶林样地,每种林分各设置4个标准样地(20 m×20 m),样地之前间隔不小于20 m。
1.2 试验设计
1.2.1野外凋落物置换试验
2019年3月在上述次生阔叶林和毛竹纯林的每个标准样地内,随机设置3个2.5 m×2.5 m的小区,小区与小区之间间隔不少于10 m;每个样地内的3个小区对应3种凋落物输入类型,本研究添加的凋落物包含叶凋落物和枯落细枝。小区建立时,清除小区内全部凋落物,保持一致状态下缓冲3个月。小区设置时具体处理措施如下:四周埋设4根1 m高的PVC管;靠近地面围设0.5 m高的尼龙网,防止周围的凋落物进入小区内;顶部用尼龙网覆盖,以阻止上方凋落物落入小区内,同时在小区边缘设置0.5 m的缓冲带。两种林分每个样地的3个小区内随机布设对照、去除地上凋落物、置换凋落物3种处理(详细说明见表1),每种林分的4个标准样地作为4次重复,两种林分共计24个小区。2019年5月起将顶部尼龙网上收集到的凋落物混匀后,等量均匀地添加到对照与置换处理的小区内,凋落物处理每个月进行1次。

表1 凋落物处理实验设置Table 1 The experimental setup for litter treatment
1.2.2室内氮素矿化培养试验
为了阐述凋落物输入对土壤氮素矿化的影响,对上述实验小区内采集的土壤样品,进行为期7天的室内氮素矿化培养试验。同时为了探讨凋落物输入改变后真菌和细菌群落对土壤氮素矿化相对贡献的变化,对野外采集的毛竹林和阔叶林对照和置换土壤样品进行添加链霉素(抑制细菌)和放线菌酮(抑制真菌)抑制剂处理。添加抑制剂处理具体为:(1)对照,土壤中不加任何抗生素;(2)放线菌酮,2 mg/g的放线菌酮注入装有10 g土壤的培养瓶;(3)链霉素,1 mg/g的链霉素注入装有10 g土壤的培养瓶;(4)放线菌酮+链霉素,2.0 mg/g的放线菌酮和1 mg/g的链霉素注入装有10 g土壤的培养瓶。
1.3 样品的采集与处理
1.3.1土壤样品采集
在凋落物处理后3个月即2019年8月25日,五点取样法用土钻采集0—20 cm的土壤样品,采集土壤样品时尽量保持各采样点的采样量一致。将土壤样品放入冰盒带回实验室,挑出石子和细根后混匀过2 mm的筛。过筛后的土壤样品分为两份;一份经含水量和持水量测定后,置于室内自然风干,磨碎过筛后用于测定土壤理化性质;另一份保存在4℃冰箱,用于室内氮素矿化培养试验。
1.3.2凋落物收集
在第一次进行凋落物置换处理时,根据实验处理将同一林分各小区凋落物收集网上的凋落物收集后混匀并称重,一部分凋落物按每月的凋落物添加量均匀撒入对照和置换处理的小区内,另一部分装入无菌自封袋,带回实验室在65℃(48 h)烘干直至恒重。称重后使用粉碎机将其粉碎,过0.15 mm筛用于测定木质素、纤维素、全碳(TC)、全氮(TN)的含量。
1.4 指标测定
1.4.1土壤理化性质和凋落物化学性质测定
土壤化学性质的测定参照《土壤农业化学分析方法》[26]。复合电极方法测定土壤pH值,水土比为2.5∶1;重铬酸钾法-外加热法测定土壤有机碳(Soil organic carbon,SOC);凯氏定氮仪-蒸馏法测定土壤全氮;靛酚蓝-比色法测定铵态氮;紫外分光光度计-比色法测定硝态氮。
凋落物化学性质测定,采用酸水解法测定纤维素和木质素含量[27];重铬酸钾法-外加热法测定凋落物全碳[26];凯氏定氮仪-蒸馏法测定凋落物全氮[26]。
1.4.2土壤净氨化、净硝化速率测定
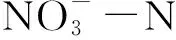
土壤净硝化速率=(培养后的硝态氮含量-培养前的硝态氮含量)/培养天数;
土壤净氨化速率=(培养后的铵态氮含量-培养前的铵态氮含量)/培养天数;
土壤净氮矿化=[(培养后的铵态氮+硝态氮含量)-(培养前的铵态氮+硝态氮含量)]/培养天数
1.5 数据处理
使用SPSS 18.0统计软件对数据进行统计分析,使用ANOVA单因素方差分析、Duncan和最小显著性差异法(LSD),对土壤化学性质以及凋落物的化学性质进行显著性分析与多重比较(P<0.05)。通过一元线性回归评估土壤氮矿化速率与凋落物化学性质和土壤理化性质之间的相关性。采用Origin 2017软件进行作图。
2 结果与分析
2.1 两种林分凋落物化学性质及凋落物处理对土壤理化性质的影响
由表2可知,与阔叶林(EBF)相比,毛竹林(PEF)凋落物全氮、木质素含量、木质素/全氮及木质素/纤维素分别高12.1%、33.4%、19.0%、69.5%,但纤维素含量低26.6%。
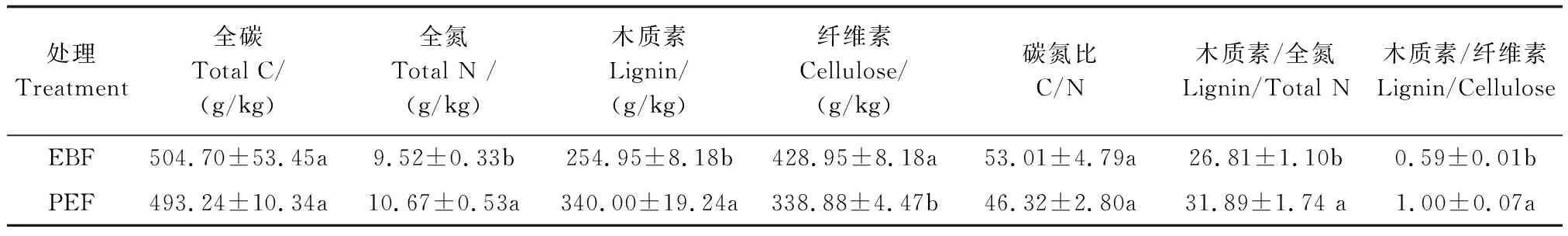
表2 两种林分凋落物化学性质Table 2 Chemical properties of litters in two forests
由表3可知,两种林分在未进行凋落物处理前,毛竹林土壤pH和全氮含量均显著高于阔叶林土壤,而C/N、铵态氮、硝态氮含量均显著低于阔叶林土壤。两种林分去除凋落物后,与对照相比,毛竹林土壤全氮和硝态氮含量分别增加了28.6%、77.2%,而有机碳、C/N分别下降27.7%、60.2%。两种林分置换凋落物后,与对照相比,阔叶林土壤C/N增加17.9%,全氮、铵态氮含量分别下降18.2%和9.1%;毛竹林土壤C/N增加27.6%,铵态氮下降19.6%。
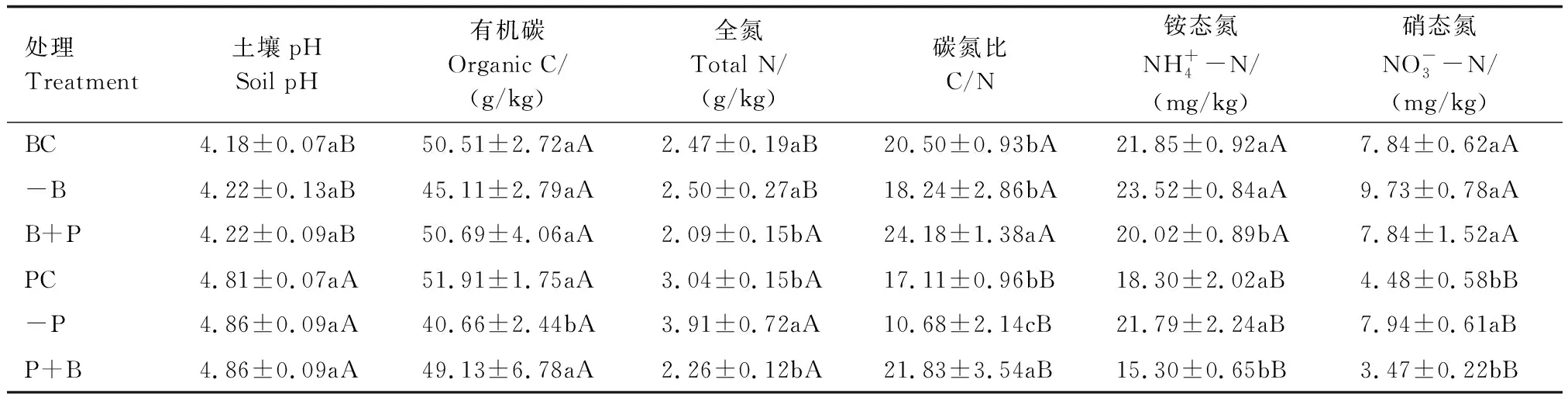
表3 两种林分凋落物处理对土壤理化性质的影响Table 3 Effects of litter treatment of two forest stands on soil physicochemical properties
2.2 两种林分凋落物处理和抑制剂添加对土壤净氮矿化速率的影响
由图1可知,阔叶林凋落物去除和毛竹林凋落物去除处理后,与各自的对照相比,土壤净氨化速率分别增加27.0%和23.4%。凋落物置换处理并未对毛竹林土壤的氨化速率造成影响,但毛竹林凋落物置换到阔叶林后土壤净氨化速率增加43.8%。为了进一步阐明置换凋落物前后真菌和细菌对土壤净氨化的贡献,分别选择两种林分各自的对照和置换处理进行抑制剂添加试验。在阔叶林对照土壤中添加抑制剂与未添加抑制剂相比,添加链霉素使土壤净氨化速率下降117.5%;添加放线菌酮使土壤净氨化速率显著升高72.0%;同时添加放线菌酮和链霉素则无显著差异。在阔叶林凋落物置换土壤中添加抑制剂与未添加抑制剂相比,添加链霉素和放线菌酮后土壤净氨化速率分别下降70.0%、51.1%,同时添加放线菌酮和链霉素之后,土壤净氨化速率无明显差异。在毛竹林对照土壤中添加放线菌酮及同时添加放线菌酮和链霉素后,与未添加抑制剂相比土壤净氨化速率分别升高67.2%、43.8%;但添加链霉素处理对土壤净氨化速率无明显影响。在毛竹林凋落物置换土壤中添加放线菌酮及同时添加放线菌酮和链霉素后,与未添加抑制剂相比土壤净氨化速率分别升高60.6%、45.8%;但添加链霉素对土壤净氨化速率无明显影响(图1)。
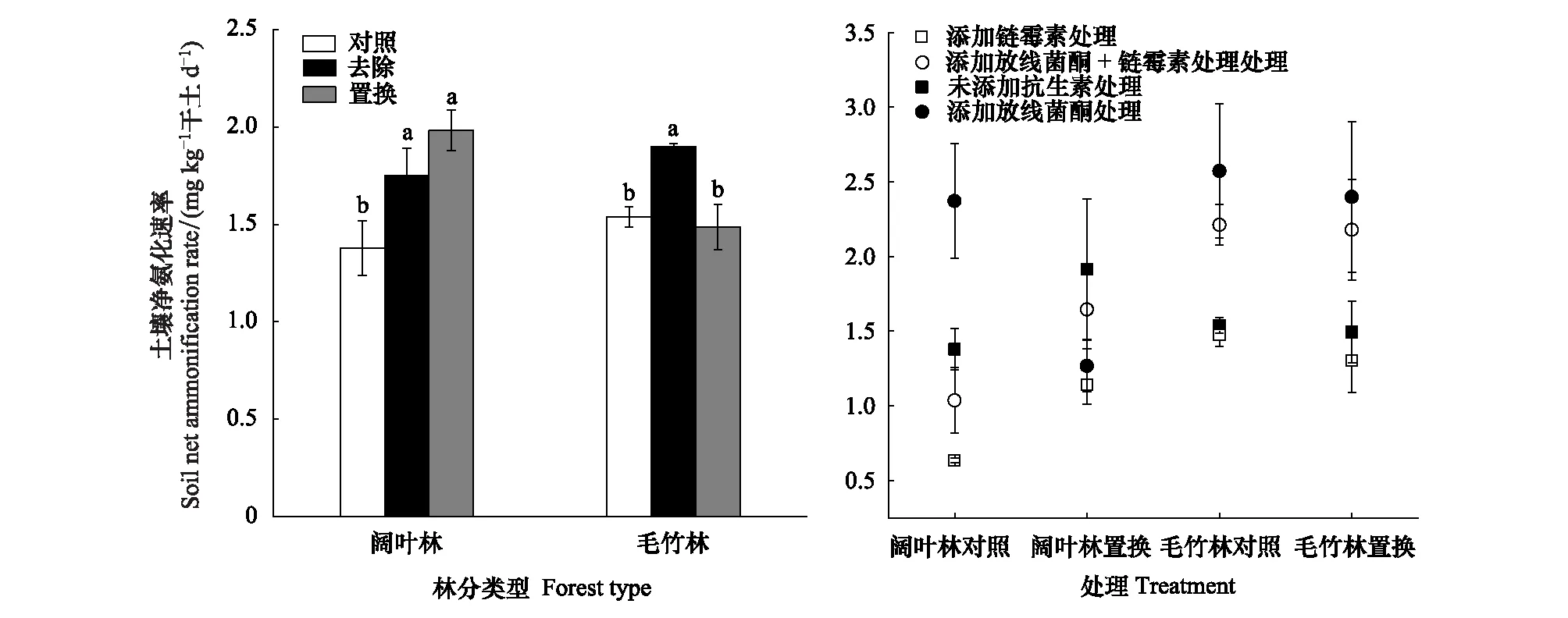
图1 毛竹林和阔叶林凋落物处理及抗生素添加对土壤净氨化速率的影响Fig.1 Effects of litter treatment and antibiotic addition on soil net ammonification rate in Moso bamboo and broadleaved forests
由图2可知,凋落物去除使阔叶林土壤净硝化速率下降11.4%,但对毛竹林土壤净硝化速率无明显影响。毛竹林凋落物置换处理与其对照相比,土壤净硝化速率增加73.1%;但阔叶林凋落物置换处理与其对照相比,土壤净硝化速率降低33.5%。在阔叶林对照土壤中添加抑制剂与未添加抑制剂相比,添加链霉素和放线菌酮使土壤净硝化速率分别下降171.5%、139.2%;同时添加链霉素和放线菌酮后土壤净硝化速率则无明显差异。阔叶林置换土壤中添加抑制剂与未添加抑制剂相比,添加放线菌酮使土壤净硝化速率下降104%,但添加链霉素、同时添加放线菌酮和链霉素使土壤净硝化速率分别升高23.5%、31.1%。毛竹林对照土壤中添加抑制剂与未添加抑制剂相比,添加放线菌酮使土壤净硝化速率下降27%;添加其它抑制剂处理均对土壤净硝化速率无显著影响。毛竹林凋落物置换土壤中添加抑制剂与未添加抑制剂相比,添加放线菌酮、链霉素使土壤净硝化速率分别下降82.8%、102.1%;但同时添加放线菌酮和链霉素对土壤净硝化速率无显著影响。
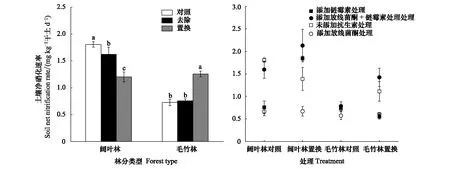
图2 毛竹林和阔叶林凋落物处理及抗生素添加对土壤净硝化速率的影响Fig.2 Effects of litter treatment and antibiotic addition on soil net nitrification rate in Moso bamboo and broadleaved forests
总体而言,凋落物去除和置换对阔叶林土壤中净氮矿化速率无明显影响;但毛竹林土壤中与对照相比,凋落物去除、置换后净氮矿化速率分别增加了17.4%、24.4%(图3)。
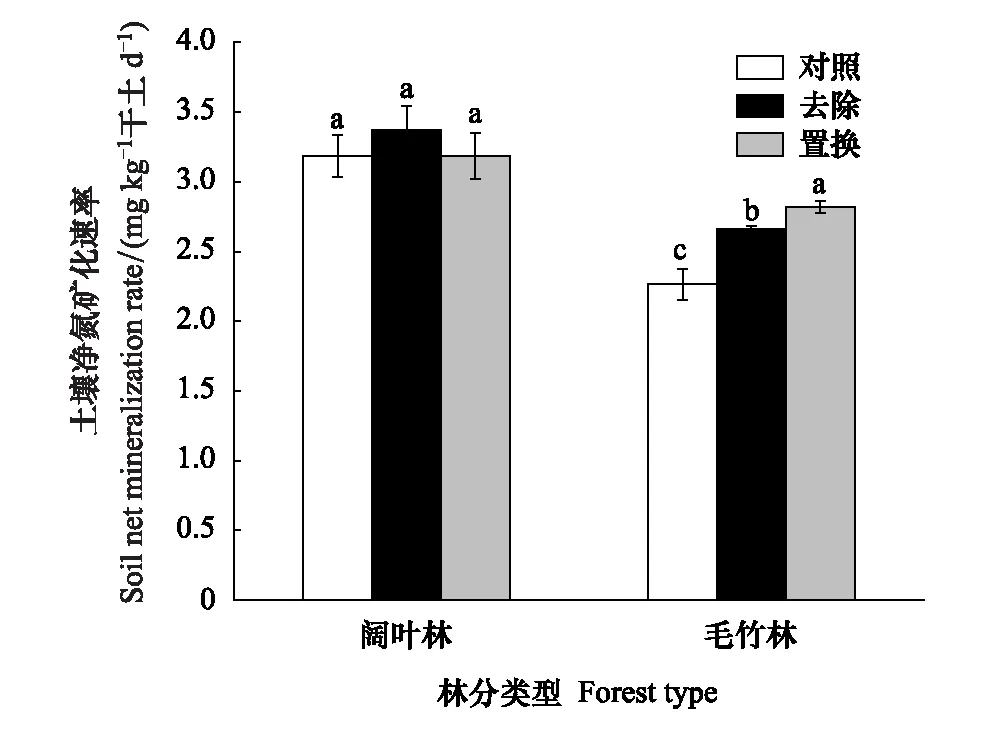
图3 毛竹林和阔叶林凋落物处理对土壤净氮矿化速率的影响Fig.3 Effects of litter treatment on soil net mineralization rate in Moso bamboo and broadleaved forests
2.3 两种林分土壤氮素矿化速率的影响因素分析
由表4可知,阔叶林土壤中净氨化速率与凋落物纤维素含量呈负相关,与凋落物木质素/纤维素呈正相关;土壤净硝化速率与凋落物纤维素含量呈正相关,与凋落物木质素/纤维素呈负相关。在毛竹林土壤中,净硝化速率仅与凋落物木质素/纤维素呈负相关。
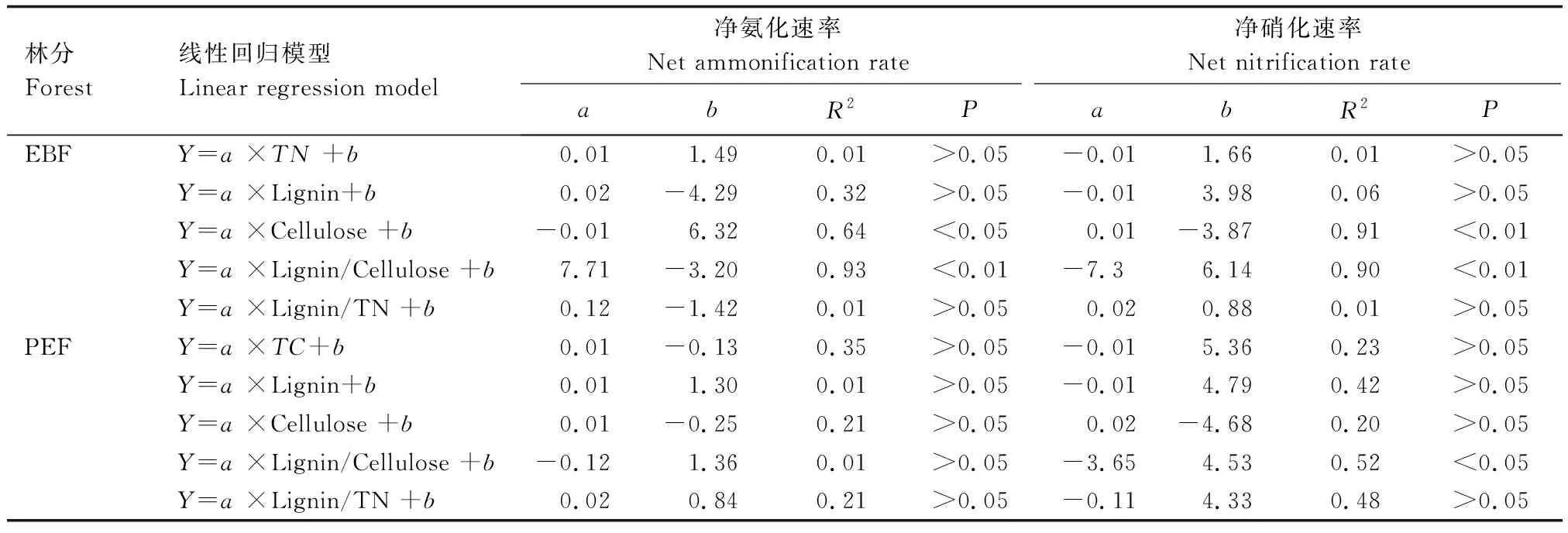
表4 两种林分凋落物化学性质与土壤氮素矿化速率之间的线性回归关系Table 4 Linear regression relationship between litter chemical properties and soil nitrogen mineralization rate in two forest stands
由表5可知,在阔叶林土壤中,净氨化速率与土壤理化性质无相关性,但净硝化速率与全氮、C/N呈正相关。在毛竹林土壤中,净氨化速率与全氮呈正相关,与有机碳和C/N呈负相关;净硝化速率与C/N呈正相关,与全氮呈负相关。
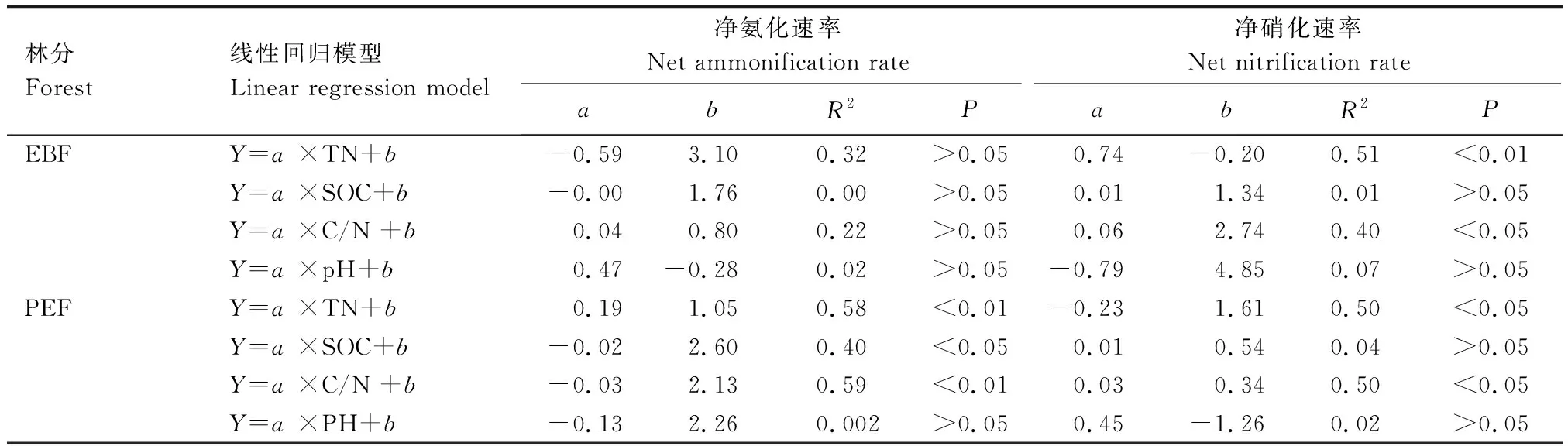
表5 两种林分土壤化学性质与氮素矿化速率之间的线性回归关系Table 5 Linear regression relationship between soil chemical properties and nitrogen mineralization rate in two forest stands
3 讨论
凋落物是土壤有机质的重要来源[28],去除凋落物使土壤碳输入方式发生改变,且通过改变土壤理化性质来影响土壤氮素矿化[29]。在本研究中,凋落物去除和置换对两种林分土壤净氮矿化速率产生的影响截然不同;去除凋落物后发现两种林分土壤净氨化速率显著升高,即铵态氮净生产增加,这与以往研究中去除凋落物使土壤铵态氮含量下降的结果[30]不一致。其原因可能与凋落物中木质素含量有关,木质素能够降解为酚类化合物,这些化合物与植物蛋白质和氨基酸结合,形成抗腐烂的聚合物,因此木质素含量过高则凋落物降解速率减慢,最终会降低土壤净氨化速率[31—32]。本研究中毛竹林和阔叶林凋落物木质素占比分别达到34%和25%,相对较高的木质素含量不利于土壤净氨化过程,因此两种林分土壤在无凋落物输入的情况下促进了净氨化过程。纤维素作为凋落物中主要的聚合物组分,是影响森林凋落物分解程度的关键调节因子[33],纤维素含量高低也会影响土壤净氨化[34]。本研究中置换凋落物后阔叶林土壤净氨化速率升高,且净氨化速率与纤维素含量呈负相关。可见阔叶林土壤中净氨化速率升高是由于毛竹林凋落物纤维素含量低于阔叶林凋落物,因此毛竹林凋落物置换到阔叶林中促使土壤净氨化速率升高。
凋落物质量是影响土壤净硝化速率的重要因素之一[35],不同类型凋落物的质量对土壤净硝化速率的影响均存在差异[20]。本研究中置换凋落物处理使毛竹林土壤净硝化速率显著升高,但在阔叶林土壤中净硝化速率则显著下降。此结果与以往研究中置换凋落物处理对土壤净硝化速率无显著影响的报道[35]不一致。存在不一致的原因可能与凋落物化学性质和分解的主客场效应有关,本研究发现毛竹林凋落物的木质素含量、木质素/纤维素较高,土壤中能够分解这些不易被降解的化学组分的微生物相对较少,阔叶林土壤中多样性更高的真菌等降解群落更加有利于难降解组分分解[22]。本研究中线性回归分析也表明,不论阔叶林还是毛竹林土壤中净硝化速率均与凋落物木质素/纤维素呈负相关。可见凋落物置换后毛竹林土壤净硝化速率增加,是由于阔叶林凋落物木质素/纤维素低于毛竹林凋落物,从而增加了毛竹林土壤中净硝化速率,阔叶林土壤则反之。本研究中去除凋落物使阔叶林土壤净硝化速率显著下降,其原因可能与有机质中难分解和易分解组分的相对数量影响了土壤氮素矿化有关[36]。Wen等[37]研究表明土壤硝化速率与土壤C/N呈正相关,即土壤C/N越低土壤净硝化速率也越低。线性回归分析表明两种林分土壤C/N与净硝化速率呈正相关,暗示去除凋落物处理导致阔叶林土壤净硝化速率降低的主要因素可能是土壤C/N。
土壤微生物控制着土壤氮的矿化过程,真菌和细菌是土壤中重要的微生物类群,但它们对土壤净氨化速率的贡献并不相同[38]。使用放线菌酮和链霉素等抗生素是研究细菌和真菌功能差异的成熟方法,在短期实验中能够对目标群体进行选择性抑制[39]。本研究在阔叶林对照土壤中抑制细菌群落发现土壤净氨化速率显著降低,抑制真菌群落后则土壤净氨化速率无显著变化,表明在阔叶林对照土壤中细菌对净氨化速率的相对贡献大于真菌。但凋落物置换后改变了阔叶林中真菌和细菌群落对土壤净氨化速率的相对贡献,除细菌外真菌群落同样对阔叶林土壤净氨化起主导作用。其原因可以从以下两个方面解释:首先,与置换处理改变凋落物质量有关,真菌在分解凋落物木质素和纤维素组分中起重要作用[40],尤其在分解木质素中占有优势[41]。本研究中毛竹林凋落物木质素含量和木质素/纤维素均显著高于阔叶林。其次,以往的研究表明,凋落物木质素含量高的土壤中真菌比较丰富,凋落物分解过程中存在较强的主场效应[10]。而在本研究中,阔叶林凋落物中的木质素含量较低,因此阔叶林中凋落物置换处理后,真菌群落对土壤净氨化速率的相对贡献增加。本研究中添加放线菌酮和同时添加放线菌酮和链霉素促进了毛竹林对照和置换处理中的土壤净氨化速率。其原因可能是因为抗生素添加后使真菌和细菌以外的群体受到抑制,而主要菌群受到抗生素和死亡微生物额外碳和氮的刺激[16],从而导致添加抑制剂后土壤净氨化速率升高。
放线菌酮和链霉素也能选择性抑制土壤净硝化速率[16]。本研究在毛竹林对照土壤中添加放线菌酮使土壤净硝化速率显著降低,添加链霉素则无显著变化;但凋落物置换后,抑制真菌和细菌都会使土壤净硝化速率降低,表明置换凋落物后除真菌外细菌群落也在土壤净硝化速率中发挥了作用。其原因可能与凋落物中的纤维素含量有关,有研究表明真菌和细菌都能够分解纤维素,并且细菌的变化主要与纤维素的分解有关[40, 42]。本研究中阔叶林凋落物纤维素含量高于毛竹林,因此置换凋落物后细菌在土壤净硝化中同样发挥了主要作用。本文的抗生素抑制实验也表明,真菌和细菌群落在阔叶林对照土壤净硝化过程中发挥作用;然而凋落物置换后,只有真菌在土壤净硝化中发挥主要作用。其原因可能是由于真菌群落是分解木质素的主要微生物类群[41],有研究表明凋落物中的木质素含量与真菌生物量呈显著正相关[43]。本研究中,与阔叶林凋落物相比,毛竹林凋落物的纤维素含量低但木质素含量高,因此置换凋落物后真菌群落可能在阔叶林土壤净硝化中发挥了主导作用。
4 结论
本研究采用野外控制试验和室内培养相结合的方法探讨毛竹入侵阔叶林生境中,凋落物处理对土壤氮素矿化的影响以及真菌和细菌对氮素矿化的相对贡献。结果表明,去除凋落物改变了两种林分土壤的净氨化和净硝化速率,去除凋落物后两种林分的土壤净氨化速率升高,而阔叶林土壤净硝化速率降低,证实凋落物输入直接调控了两种林分土壤氮素矿化过程。另外,本文从凋落物质量的角度,探讨了两种林分凋落物处理后土壤净氨化和净硝化速率的变化,结果表明置换凋落物后,凋落物纤维素含量与阔叶林土壤净氨化速率呈负相关;两种林分土壤中净硝化速率均与凋落物木质素/纤维素呈负相关。可见毛竹林凋落物输入使铵态氮净生产增加,阔叶林凋落物输入使硝态氮净生产增加。特别是两种林分凋落物木质素和纤维素含量的差异,使得真菌和细菌对土壤净氨化和净硝化的相对贡献发生改变。本文解析了凋落物质量在毛竹扩张改变土壤氮素矿化中起到的重要作用,未来的研究需要在更长时间尺度上探讨凋落物输入改变土壤氮素矿化的过程,为阐明毛竹入侵阔叶林的地下生态机制提供理论依据。
