重庆喀斯特生境中桢楠种群结构与动态特征
2022-08-03何雪娜王佳敏刘俊婷谭兴旺刘锦春
雷 颖,何雪娜,王佳敏,刘俊婷,谭兴旺,赵 彬,刘锦春,*
1 西南大学生命科学学院,三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室,重庆 400715 2 重庆市永川区国有林场,重庆 402100
种群结构与动态特征是种群生态学研究的两大核心[1—2],是植物种群的生物学特性对环境长期适应和选择的结果[3]。珍稀渐危树种在植物的系统演化和遗传育种中占据着重要地位,且在经济、科研和文化等方面有着较高的价值,其数量稀少,生境特殊[4]。珍稀保护树种在特定生境下的种群结构和动态特征以及繁殖更新情况,可以反映珍稀树种与环境之间的相互关系,对分析种群的现实情况、预测种群未来的走向以及实施合理的保护措施有重要意义。
桢楠(PhoebezhennanS. Lee)为樟科(Lauraceae)楠属(PhoebeNees)的高大乔木,是珍稀渐危树种[5—6]。由于历年来对桢楠的滥砍滥伐,其原生林几乎被毁,加上桢楠后期生长缓慢[7]和遗传多样性较低[8]等原因,导致现存资源有限且分布零散[9]。研究表明,桢楠喜湿耐荫,适宜在酸性土壤的山腰山谷中生存[10],野生多散生于海拔1500 m以下的亚热带常绿阔叶林中[5,11]。然而,在野外调查中发现重庆市大足区及永川区的喀斯特生境中也存在长势良好的桢楠林。
喀斯特生境是一种整体资源量低的脆弱生境,其岩石裸露率高、土壤浅薄且呈斑块化分布、土壤营养贫瘠且持水性能较差[12]。由于生境特点和人类活动的影响,喀斯特地区的植被常呈退化之势,乔木树种生存困难。受喀斯特生境独特地貌形成的地理屏障限制,植物种子很难通过水流、风力和重力等非生物因子进行长距离传播而到达到适宜其萌发的微生境中;且喀斯特的特殊生境也不利于后期种子的萌发和幼苗的生长,从而对后期植物种群的空间分布产生重要影响[13]。因此我们推测喀斯特地区的桢楠种群可能存在结构不稳定、更新困难等问题。
目前对于桢楠的研究大多集中在抗性生理[14—15]、营养生理[16—17]和群落特征[18—20]等方面,也有少数研究关注了部分楠木种群的结构和动态特征。如吴大荣等[21]研究了福建罗卜岩闽楠种群,发现该种群空间分布格局总体上具有由集群向随机分布的趋势,具有较强的自然更新的能力。徐振东[18]和李贤碧等[22]研究表明,贵州思南县楠木种群和湖北利川楠木种群所在群落物种组成均较丰富,更新能力强。然而,曾凡勇和冯邦贤[23]的研究却发现,德江沙溪楠木群落物种多样性整体偏低,更新能力较弱。可见,楠木种群结构和动态特征与其所在的群落环境及群落物种组成有一定的关系。
因此,本研究通过对重庆喀斯特生境中的两个桢楠种群的年龄结构、数量动态特征与空间分布格局的定量研究,明确其在喀斯特生境中的生存现状,揭示其动态规律和未来发展趋势,以期为桢楠种群的保护、群落更新和植被恢复提供理论依据。
1 研究区概况
研究区域选择重庆市大足区高坪镇纸槽沟和永川区太岚坳,是桢楠的天然群落分布区。该区四季分明,春季回温早,夏季气温高,降雨分布不均匀,光照充足,雨热同季,属于亚热带季风气候。地貌以丘陵、山地为主,坡地面积较大。土壤为黄壤土,中性至微酸性[7]。
重庆市大足区高坪镇纸槽沟(29°50′N,105°39′E),海拔480 m,境内分布的野生楠木占地约200 hm2,集中成片达107 hm2,林内百年树龄以上2万余棵,主要以黄绒润楠(Machilusgrijsii)为主。该楠木林已有两三百年历史,一直由该地村民守护。该区年均无霜期323 d,全年均降水量727 mm,集中在5—9月。该区域内有高等植物24科28属31种。
重庆市永川区太岚坳(29°33′N,105°53′E)大碑寺公路附近,海拔628 m,境内分布的天然桢楠占地约64 hm2,林内最高树龄有上百年。当地村民从1957年起对此林进行严格管理,现林场为桢楠建立了一套完整的保护体系,实现全方位立体化的保护。该区年均无霜期317 d,年均降水量1015 mm,集中在5—8月。该区域内有高等植物26科36属38种。
2 研究方法
2.1 样地设置与调查
由于两个区域的桢楠林林分均比较简单,因此设计的样方面积均为10 m×10 m。根据地形特点,大足区的桢楠林中共设置7个样方,呈“一”字形排列,永川区共设置10个样方,呈环状分布。调查并记录所有样方内桢楠的个体数、树高、胸径、高度和冠幅等指标。同时测定和记录各样地的海拔、坡度、坡向、郁闭度、草本覆盖度和凋落物厚度(表1)。
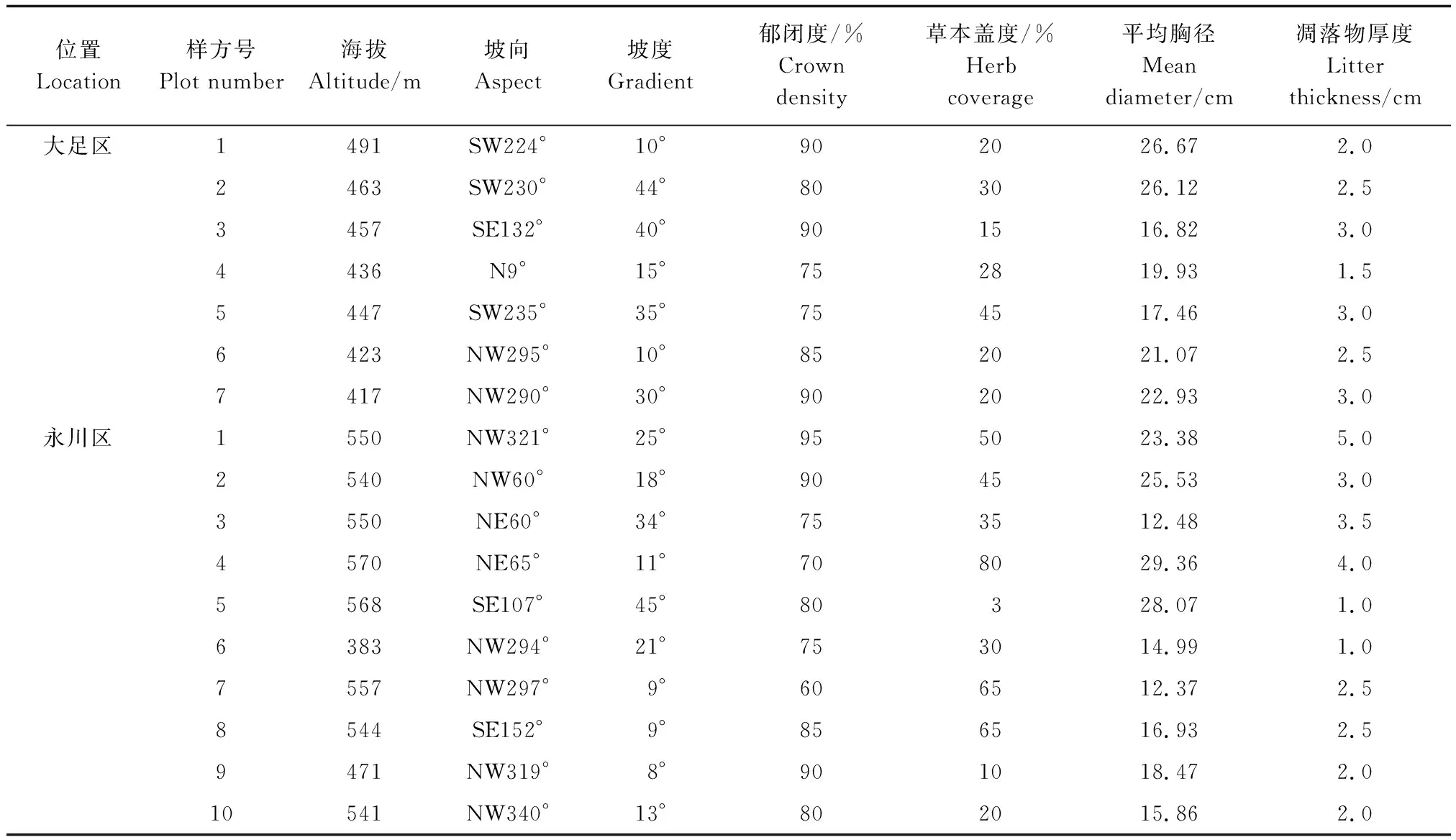
表1 桢楠种群环境条件Table 1 The environmental conditions of P. zhennan population
2.2 种群径级划分
由于同一树种的年龄结构(龄级)和径级对相同的环境所呈现出的规律具有一致性[24],再加上树木年龄获取比较困难,因此采用径级结构代替年龄结构的方法。根据所得调查数据和已有研究[7,25—26],将桢楠划分为8个龄级:第Ⅰ龄级,幼苗,H<2.5 m;第Ⅱ龄级,幼树,H≥2.5 m,胸径<10 cm;第Ⅲ龄级,中树,10 cm≤胸径<15 cm;第Ⅳ龄级,中树,15 cm≤胸径<20 cm;第Ⅴ龄级,成树,20 cm≤胸径<25 cm;第Ⅵ龄级,成树,25 cm≤胸径<30 cm;第Ⅶ龄级,成树,30 cm≤胸径<35 cm;第Ⅷ龄级,老树,胸径≥35cm。
2.3 静态生命表及存活曲线
静态生命表是用特定时间点上调查种群内各个年龄组上存活状况的数据编制而成的[27]。
在调查中系统误差不可避免,有可能会出现生命表中的死亡率为负值的情况,这是由于静态生命表反映的是世代重叠的年龄动态历程中的一个特定时间,而不是该种群的全部生活史[28]。因此在编制生命表之前对实测数据进行匀滑处理[29]得到ax(实际存活个体数经匀滑处理后所得存活数),编制出桢楠种群的静态生命表。
此外,采用Hett和Loucks[30]的方法检验桢楠种群的存活曲线。指数方程Nx=N0e-bx描述Deevey-Ⅱ型存活曲线,幂函数Nx=N0x-b描述Deevey-Ⅲ型存活曲线。
2.4 种群年龄结构及动态分析
运用陈晓德[31]的种群结构动态量化分析方法,得到桢楠种群年龄结构的动态指数,从而对种群动态(V)进行定量的描述。在方法中,Vn取正值、负值、零值时分别反映种群两相邻龄级个体数量的增长、衰退、稳定的结构动态关系;Vpi、V′pi取正值、负值、零值分别反映种群个体数量的增长、衰退、稳定。计算公式如下:
式中,Sn与Sn+1分别为第n和第n+1龄级的个体数;k为种群大小数量级。
2.5 种群空间结构分析
采用扩散系数(C),并进行t检验,从而判断桢楠的空间分布格局,同时使用负二项指数(K)、丛生指数(I)、平均拥挤度(M*)和聚块性指数(PAI)、以及聚集度指数(Ca)等多个指标拟合桢楠种群的分布格局[32]。具体计算公式如下:
(1)扩散系数C
C=V/m
当C>1、C=1 和C<1时,种群表现为聚集、随机和均匀分布。
(2)负二项指数K
当K<0时为均匀分布;K>0为聚集分布,低K值表示丛生比较显著,即聚集强度大,高K值表示丛生程度低,聚集强度弱;当K值趋于无穷大时(一般为K<8),种群逼近泊松分布。
(3)丛生指数I
丛生指数I>0、I=0和I<0时,种群分别表现为为聚集、随机和均匀分布。
(4)平均拥挤度和聚块性指标
PAI=M*/m
M*为平均拥挤度,代表的是样方内生物个体的拥挤程度;PAI为聚块性指标,当PAI>1时,为聚集分布;PAI=1时,为随机分布;PAI<1时,为均匀分布。
(5)聚集指数
当Ca<0、Ca=0、Ca>0时,种群分别表现为均匀分布、随机分布、聚集分布。
以上所有公式中V和m均分别表示种群方差和均值。
3 结果与分析
3.1 种群年龄结构
大足区与永川区两个桢楠种群年龄结构均为幼苗、幼树株数多,中、成、老树个体比例大致相同,整体为增长型(图1)。在桢楠种群实际调查中,样方内的幼苗由于生存空间的限制不可能全部存活,且桢楠幼苗20 cm以下的个体成活率较低。大足区的第Ⅰ龄级幼苗平均高度0.42 m,0.2 m以上的幼苗占总体幼苗的60%(16棵),由此来看,大足区桢楠种群各龄级数目相当,幼苗、幼树、中树、老树比例基本相同,大足桢楠种群的实际年龄结构应该为稳定型。永川区的0.2 m以上的幼苗占总体幼苗的74%(248棵),第Ⅰ龄级幼苗远大于其他龄级,但永川区桢楠种群依然呈幼树多的增长型。
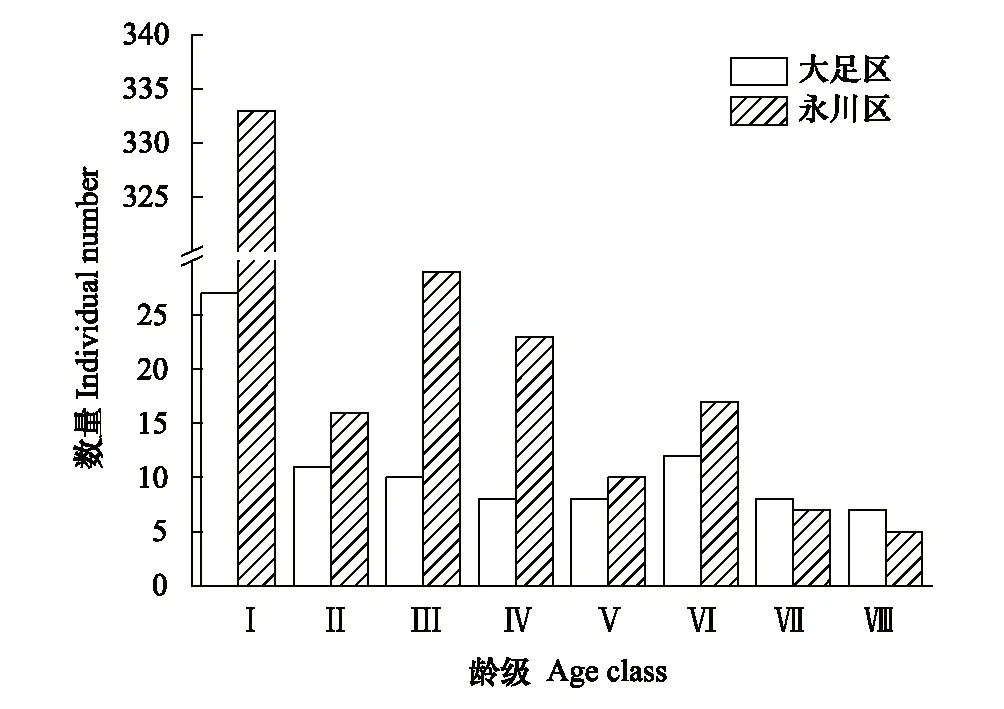
图1 桢楠种群龄级结构图Fig.1 Age class structure diagram of P. zhennan populationⅠ—Ⅷ表示桢楠的年龄结构划分为第1—8龄级
3.2 静态生命表与存活曲线
随着径级的增加,两个种群的标准化存活个体数(lx)、消失率(Kx)和死亡率(qx)逐渐减小,说明桢楠幼苗、幼树的死亡率较高,种群易受外界环境的干扰(表2)。大足区桢楠种群的生命期望(ex)在幼苗、幼树和中树(Ⅰ—Ⅳ龄级)阶段较高,并且随着个体的生长而下降,符合种群生态学特征。永川区种群中树的生命期望(ex)达到顶峰,说明该种群正处于活力旺盛的时期,此后开始逐渐降低。
大足区桢楠种群前期死亡率较高,后期较低(表2、图2);永川区桢楠种群幼苗幼树死亡率较高(表2、图2)。两个桢楠种群存活曲线均与Deevey-Ⅲ型拟合程度较高(表3)。
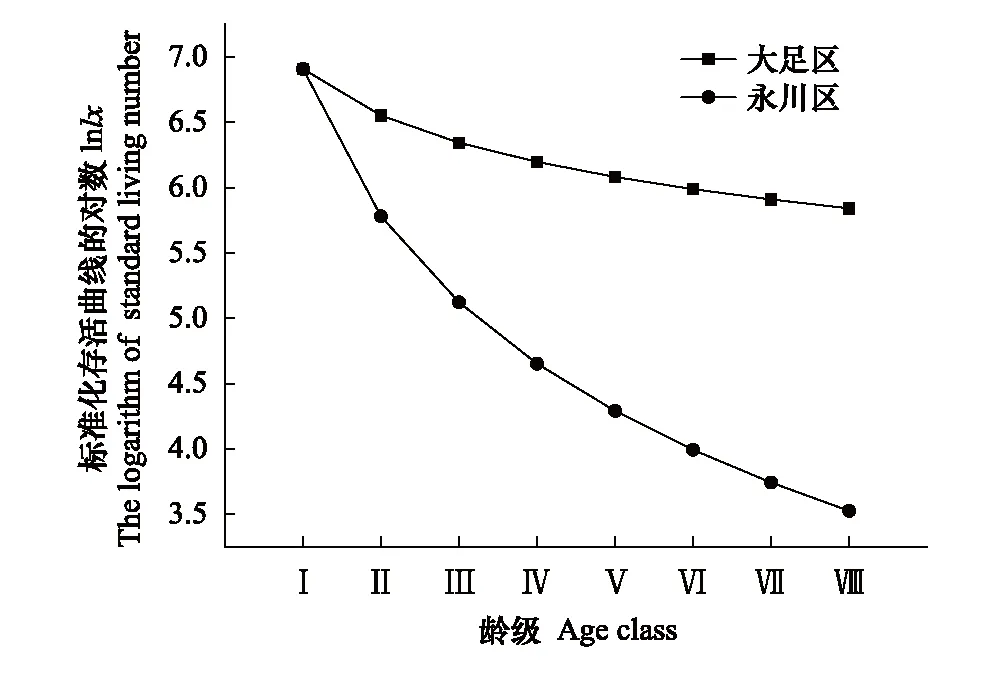
图2 桢楠种群存活曲线Fig.2 Survival curve of P. zhennan population
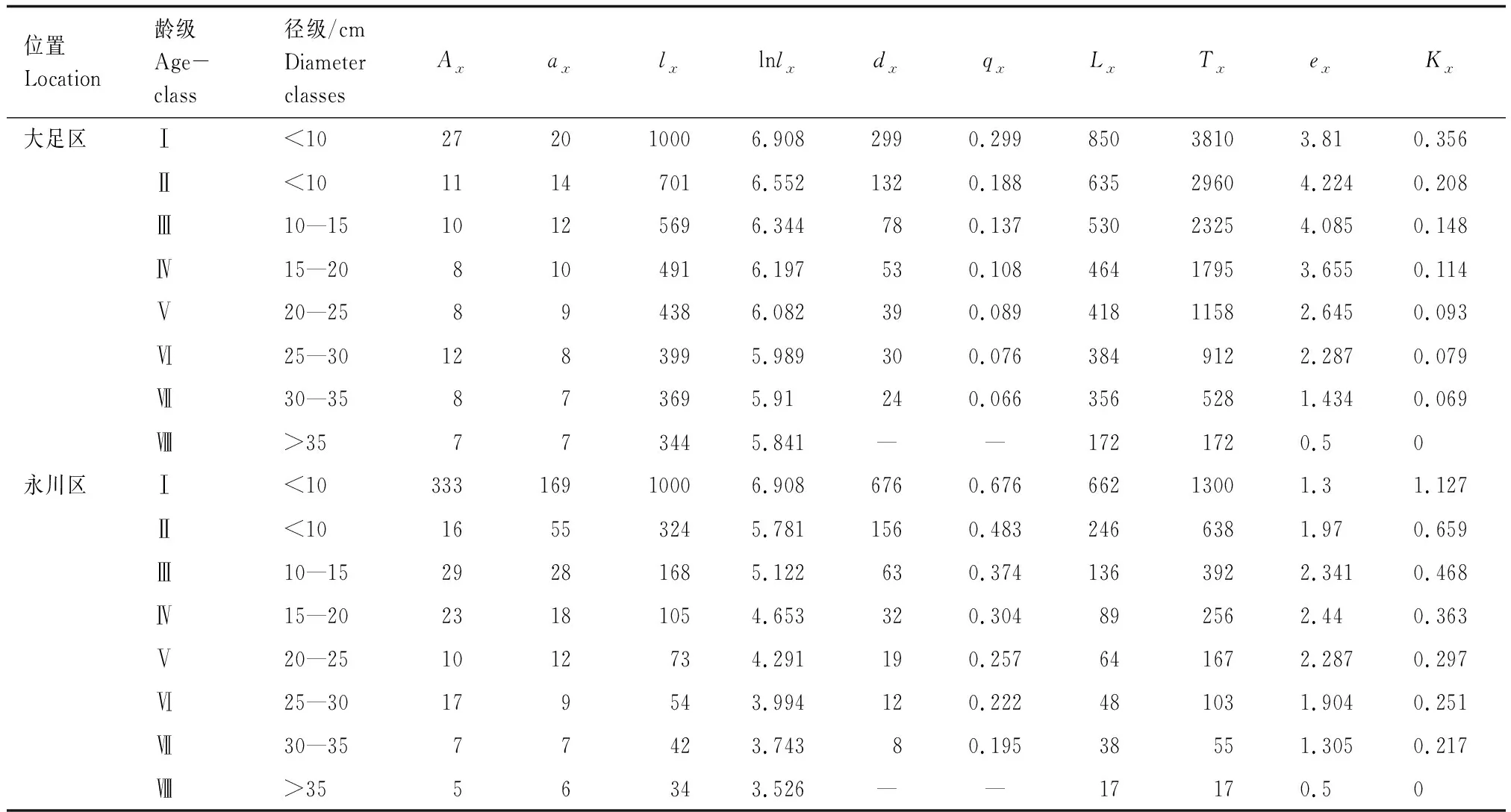
表2 桢楠种群静态生命表Table 2 Static life table of P. zhennan population
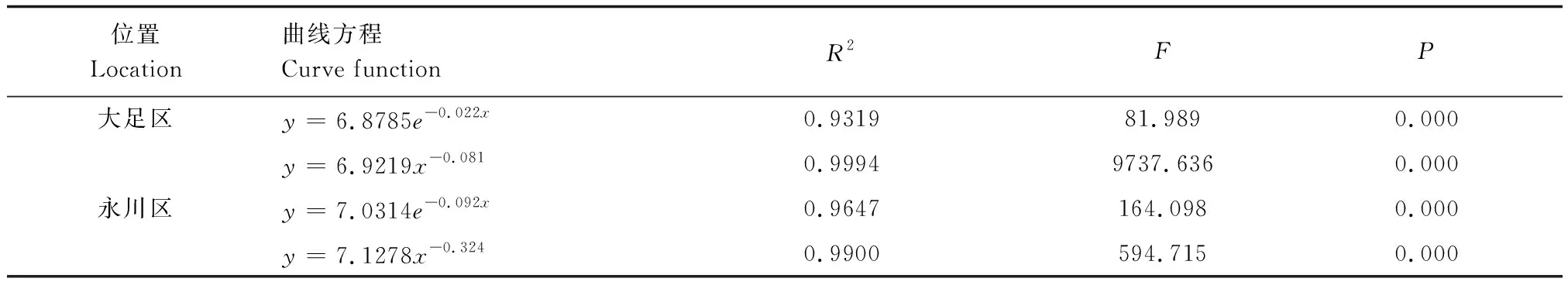
表3 桢楠种群存活曲线的检验模型Table 3 Test models of survival curves of P. zhennan population
3.3 种群年龄结构的动态变化分析
大足区桢楠种群在V1、V2、V3、V6和V7均大于0,种群呈现增长的结构动态,V4等于0,种群呈稳定状态,V5小于0,表明这个龄级的桢楠种群呈衰退状态;永川区的V2和V5均小于0,种群呈衰退状态(表4)。

表4 桢楠种群年龄结构动态指数/%Table 4 Dvnamic indices of P. zhennan population age structure
种群总体动态变化指数分析结果显示:大足区(Vpi=25.40%)和永川区(Vpi=77.40%)种群动态指数均大于0,种群整体结构属于增长型(表4)。当不忽略外部干扰时,种群龄级结构动态指数V′pi=0.45%和V′pi=1.94%均趋于0,且种群结构对随机干扰的敏感指数,即随机干扰风险的极大值Pmax=1.79%(大足)、Pmax=2.50%(永川)(表4),说明永川的桢楠种群结构对随机干扰的敏感性更大。因此,当两个种群在受到外部干扰的情况下,大体上仍然是增长型的种群,只不过增长速度较慢,并且对外界干扰敏感度高。
3.4 桢楠种群空间分布格局
大足区桢楠种群呈随机分布,聚集指数为0.07;永川区桢楠种群呈聚集分布,聚集指数为5.35(表5)。两个种群在不同龄级上均呈聚集分布,从聚集强度来看,大足区和永川区桢楠种群均为幼苗聚集程度最大(表6)。
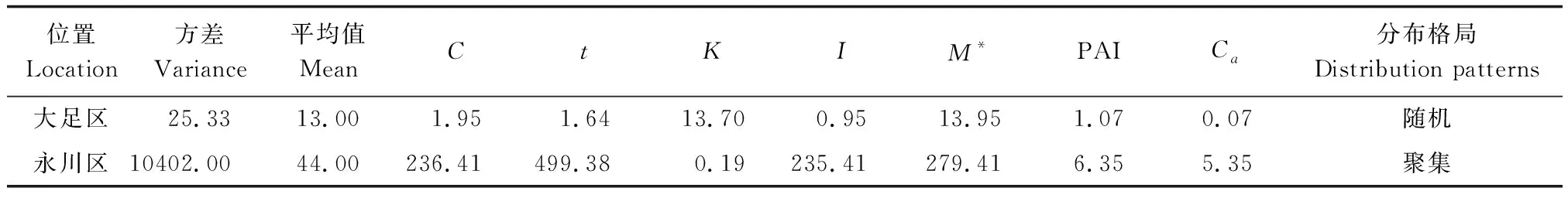
表5 桢楠种群分布格局Table 5 Distribution pattern of P. zhennan population
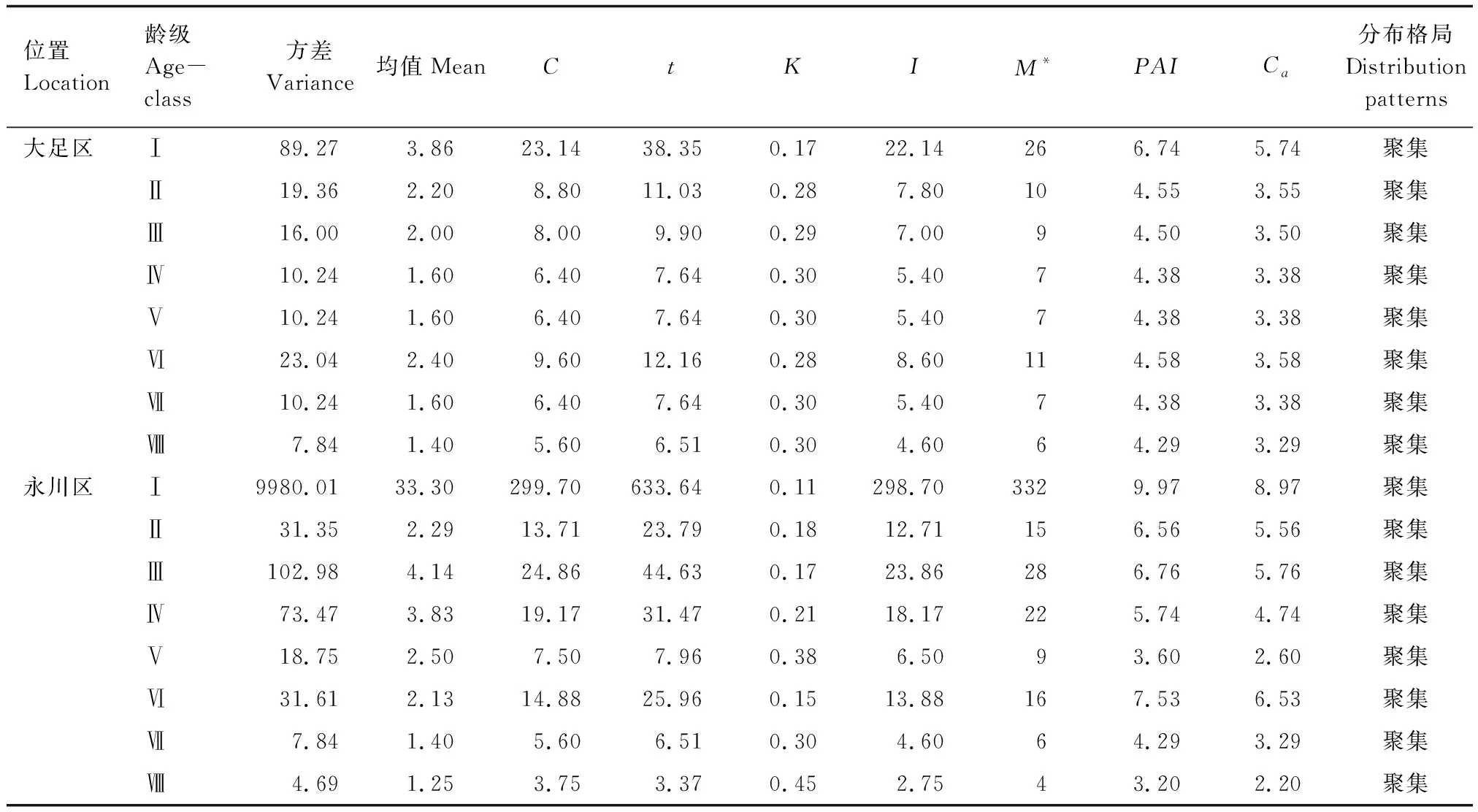
表6 桢楠不同径级分布格局Table 6 Distribution pattern of different diameter classes in P. zhennan
4 讨论
多数研究者认为桢楠适合生长在土壤深厚、温润潮湿的地方,而喀斯特地区土层浅薄,且容易出现干湿交替的现象,不宜桢楠生长。然而,大足区桢楠种群年龄结构属于稳定型,永川区种群年龄结构属于增长型,这与种群动态分析的结果一致,说明桢楠也能在喀斯特地区生长,但是两个区域的种群动态量化分析中仍存在局部龄级间衰退的现象。
大足区种群成树阶段呈衰退状态,永川区的幼树和成树也出现类似的情况,即在幼树、成树阶段呈衰退型。由于两地区的桢楠一直受到当地村民的保护,因此推测该阶段的生存压力主要是环境和种群自我调节导致的。桢楠幼树有一定的耐阴性[33],而成树对光的需求逐渐增强[34],且桢楠林郁闭度均在70%以上。随着龄级的增长,所需养分和光照逐渐增加,导致部分竞争力弱的成树无法进入林冠层。Janzen[35]提出动物取食、幼苗竞争的强度会受种子/幼苗与母株的距离有关,越近发生的频率越高。由于永川区20 cm以下的幼苗较多且呈聚集型分布(5号、6号样地凋落物厚度较小,种子萌发率远高于其他样地),推测其密度效应较强,种内竞争过于激烈且幼苗数量很可能远远超过了当地的环境容纳量,大部分幼苗无法成功转向幼树阶段,所以在一定范围内出现大量幼苗,且幼苗都在母株周围,大大提高了桢楠幼苗的死亡率。这与Comita和Hubbell等[36]研究的幼苗密度与耐阴性对幼苗存活影响的结果一致。再加上喀斯特地区土壤资源匮乏,水土易从岩石裂隙中流失,雨后即处于干旱缺水的状态[37],这对抵抗能力差的幼苗、幼树来说是致命,因此呈衰退状态。
从生命表、存活曲线和种群动态分析来看,大足区和永川区桢楠种群前期幼苗和幼树死亡率较高,此后死亡率逐渐下降。从种群年龄结构动态变化指数来看,在不忽略外部干扰时,两个桢楠种群整体结构呈增长型,这与湖北来凤县桢楠种群[9]的调查结果相反。其主要原因是大足区和永川区的桢楠种群长期以来受到人为干扰的强度较小,而来凤县的桢楠受人为砍伐严重。但这两个区域的桢楠种群也存在动态指数小于0的阶段,说明两个种群增长缓慢且对外界干扰敏感度高。
桢楠种群的空间分布格局受自身繁殖方式和环境的影响较大。整体而言,大足区桢楠种群的空间分布格局为随机分布,永川区桢楠种群为集群分布。研究表明,集群分布是楠木种群常见的分布方式[21]。原因是桢楠的种子传播能力不强,主要是靠自由掉落和鸟类传播,因此常聚集在母树附近,呈现聚集分布。永川区桢楠种群集群分布,一是与桢楠的种子传播力有关,另一方面也与其地形原因不易扩散有关。这也可能是对喀斯特环境的一种积极响应策略,因为聚集分布可以增强种群抵抗外界干扰的能力。而大足区的桢楠种群呈现随机分布的状况,其原因可能是种群内部之间的竞争导致的,幼苗充足,且幼苗到成树之间的竞争较为激烈,因而对资源(光照、水分和养分等)的需求加大,竞争加剧,从而形成均匀分布的格局。同时,大足区与永川区的森林小气候的不同,可能也是导致桢楠种群空间分布格局差异的另一原因,永川区的年平均降雨量比大足区多288 mm,土壤微酸性,海拔较高,因此永川区更适合桢楠的生存,且在一定范围内可利用的资源相对丰富,能容纳的桢楠就更多。此外,大足区楠木林的范围是永川区桢楠林的3倍左右,可能是由于两者的尺度不同导致调查结果有差异,这也能看出桢楠种群发育过程中,分布格局由集群分布向随机分布发展。
可见,重庆大足区与永川区两个桢楠种群生存现状良好,整体呈现稳定趋势,但局部龄级间仍存衰退现象。与我们的推测相反,喀斯特生境中桢楠种群仍有较强的更新能力,但因种群生长周期长[7],遗传多样性较低[8],对外部干扰敏感,适应能力较差。因此,对桢楠种群的保护仍不能掉以轻心,需要制定适宜的保护措施,进行科学合理的管理。同时建议在幼苗幼树死亡率高的情况下可以采用人工辅助措施,将幼苗从聚集的地方移植一部分出去,减少内部的竞争压力,待幼苗长成小树时再将其移植到合适的野外分布区,在现有条件下不断扩大桢楠资源,创造更多的生态价值和经济价值。
