分子标记在绣球属育种中的应用研究进展
2022-08-02陆泓锦吴宜静张一鸣赵天祎杨玉勇张启翔
陆泓锦,吴宜静,张一鸣,续 言,王 露,赵天祎,章 寒,蔡 明*,杨玉勇,张启翔
(1.花卉种质创新与分子育种北京市重点实验室,国家花卉工程技术研究中心,城乡生态环境北京实验室,园林环境教育部工程研究中心,林木花卉遗传育种教育部重点实验室,北京林业大学 园林学院,北京 100083;2昆明杨月季园艺有限责任公司,云南 昆明 650500)
绣球(Hydrangeamacrophylla)又名八仙花、紫阳花,是虎耳草科绣球属植物[1]。绣球属植物花序硕大,花色丰富,观赏价值极高,可作为切花、盆花、园林绿化材料应用,深受世界各国人们喜爱[2]。我国是绣球属种质资源分布中心,但资源利用率低。目前国内生产与应用的品种多引自国外,缺少具备市场竞争力的自育品种,且育种技术落后,因此需要对绣球种质资源进行系统的收集与研究,改善育种技术,提高利用率。
分子标记是根据基因组DNA存在丰富的多态性而发展起来的可直接反映生物个体DNA水平上的差异的一类新型的遗传标记,它是继形态学标记、细胞学标记、生化标记之后最为可靠的遗传标记技术。通常所说的分子标记是指以DNA多态性为基础的遗传标记。可以分成以下3类:第1类是以分子杂交为核心的DNA分子标记 RFLP;第2类是以聚合酶链式反应(PCR)为核心的DNA分子标记 RAPD、SSR 、ISSR、AFLP、SCAR、STS等。第3类是一些新型分子标记如SNP、InDel、CNV、SV等。常见的分子标记名称以及定义如表1所示。DNA分子标记在种质资源鉴定与分子育种等方面具有广阔的应用前景。自20世纪80年代引入分子标记以来,育种研究得到了深度上的扩展和步骤上的简化[4],分子标记数量丰富,遗传稳定,多态性高,多为共显性,不仅广泛应用于植物种质鉴定与评价[5]、植物遗传多样性[6]、亲缘关系鉴定[7]、杂交后代鉴定[8]、重要性状基因定位以及分子标记连锁图的构建[9]等方面的研究,已成为观赏植物种质资源鉴定与评价、重要性状遗传规律分析、缩短育种周期、定向改良观赏性状和杂交后代早期鉴定与选择的重要手段。本研究整理了不同分子标记技术应用于绣球属植物研究的情况,为绣球属植物资源评价与利用奠定基础。

表1 常见分子标记名称及定义Table 1 Commonly used molecular marker names and definitions
1 分子标记在种质资源鉴定与评价研究中的应用
1.1 资源鉴定
分子标记能够较为准确地区分品种,便于后期管理与应用。T.A.Rinehart等[11]利用38个SSR位点构建系统发生树,对部分绣球品种名称提出了修改建议,绣球栽培品种Hydrangeamacrophylla‘Libelle’与Hydrangeamacrophylla‘Libelle White’为同一品种,品种名中‘White’用来描述颜色,是为了便于市场流通而后期添加;Hydrangeamacrophylla‘Pink Beauty’与Hydrangeamacrophylla‘Preziosa’的DNA指纹图谱相同,可能为同一品种;部分圆锥绣球(Hydrangeapaniculata)品种基因型一致,应进行统一命名;Hydrangeamacrophylla‘Reewee’的绣球品种实际上包含2个不同品种[12-13],应区分命名。P.Hempel等[14]利用SSR分子标记技术构建了Wädenswil家族图谱,纠正了Wädenswil家族中部分品种的命名。J.T.Lindstorm等[15]使用11个RAPD引物区别鉴定出7个绣球品种,包含5个多次开花品种,2个抗寒品种,其中5个多次开花品种得到了很好的区分,但上述引物未能区分开Hydrangeamacrophylla‘Endless Summer’与Hydrangeamacrophylla‘David Ramsay’,推测其为相同品种,或由于引物多态性较差,不足够将其区分。D.Zlesak等[16]发现绣球Hydrangeamacrophylla‘Bailday’与名称较为相似的Hydrangeamacrophylla‘Bailmer’可被156个AFLP条带区分,推测二者不是同一品种;而H.macrophylla‘Bailday’与表型相似的Hydrangeamacrophylla‘Varigata’仅在1个位点上有差别,推测‘Varigata’可能是‘Bailday’的衍生品种,或是同一品种。Y.H.Joung等[17]结合叶片等表型信息,将29种绣球资源利用RAPD进行聚类分析,此外还进行了基于rbcL基因获得的SNP标记的单体型分析。BS-UPGMA和IB-NJ聚类分析都将29个绣球资源分成了2个组,并将不知名品种鉴定为一种冠盖绣球(Hydrangeaanomalavar.petiolaris)。李艳香[18]采用SRAP分子标记技术对绣球属11个野生种和8个品种进行了亲缘关系研究,从110对引物中筛选出14条扩增条带清晰、多态性丰富的引物组合,其中一个引物组合可以把19种绣球品种区别开。E.Mortreau等[19]利用25个ISSR引物,测试了7个绣球属植物,UPGMA聚类分析将7种植物分成了总苞绣球(Hydrangeainvolucrata)和马桑绣球(Hydrangeaaspera)2组,总苞绣球的个体之间区分明显,马桑绣球被细分成了5个小组,鉴定结果与地理学数据、表型分析数据、生理学、染色体学数据吻合。地理分布较广泛的种类,其遗传多样性通常较高。J.H.Lee等[20]从64个AFLP引物组合筛选出2个组合对22种绣球属植物进行遗传多样性分析,共检测到120条DNA片段,通过UPGMA聚类分析发现,17个绣球品种和山绣球(Hydrangeamacrophyllavar.normalis)聚类到一组,其余4种韩国栽培品种聚类到一组,韩国栽培品种与绣球品种遗传距离相对较大,得到很好区分,并推测山绣球是野生原始种。S.Yamamoto等[21]利用19个来自日本的粗齿绣球品种和14个来自韩国济州岛的粗齿绣球品种共188株的遗传结构进行SSR分子标记分析。结果表明,济州岛种群的遗传结构与日本西部地区种群的遗传结构密切相关,日本西部和西北部地区采样的粗齿绣球与在日本中部和东部地区采样的粗齿绣球的遗传结构存在差异。此外,韩国发现本土的黄脉绣球(Hydrangealuteovenosa),其在日本西部分布广泛,但在韩国是濒危种[22]。T.Ito等[23]在4个来自韩国和日本的黄脉绣球中利用SSR分子标记技术研究分析,发现在23对引物中有5个SSR位点在基因上都有清晰条带。H.J.Choi等[24]利用上述5个SSR标记对韩国黄脉绣球和3种日本黄脉绣球进行分析,发现存在于韩国285个个体中的2个多座位基因型在日本个体中并未被检测到,推测韩国黄脉绣球可能引自日本。
1.2 遗传多样性分析
彭继庆等[25]利用ISSR-PCR 分子标记技术对23个绣球品种进行遗传多样性研究,从100条引物中筛选出10条引物对供试材料基因组总DNA进行PCR扩增,共扩增出条带123条,其中多态性条带102条,经PopGen32软件包分析,23个绣球花品种的平均有效等位基因数为1.493 7,平均 Nei’s基因多样性指数为0.290 7,平均Shannon's信息指数为0.435 7,遗传距离介于0~0.769 1,遗传一致度介于0.463 4~1.000 0,具有广泛的遗传多样性。此外彭继庆等[26]还利用上述ISSR分子标记的10条引物以绣球和圆锥绣球的38个品种为材料进行遗传多样性分析,绣球、圆锥绣球2个种的多态条带比例分别79.55%、 67.2%;38个绣球花品种观测等位基因数为1.924 2,有效等位基因数为1.676 8,Nei's 基因多样性指数为0.378 2,Shannon's 信息指数为0.549 7,具有较高的遗传多样性和较强的环境适应能力;绣球、圆锥绣球2个种的基因多样性为0.375 8,种内基因多样性为0.282 6,种内遗传变异占75.20%。T.Uemachi等[27]从426个RAPD标记中筛选出413有多态性的标记,对山绣球和2种粗齿绣球(Hydrangeaserratavar.serrata、Hydrangeaserratavar.yesoensis)进行UPGMA分析,发现RAPD和叶绿体DNA分析均表明山绣球遗传多样性高于另外2个,推测是因为其有广泛的地理分布。
1.3 亲缘关系分析
绣球属早期亲缘关系分析常用表型分析、核型分析、同工酶分析等[28-29],而分子标记技术具有较高的准确性与可信度,是近年来亲缘关系研究的主要方法。T.A.Rinehart等[13]使用14个SSR标记对85个绣球属和近缘属资源进行分析,聚类结果印证了大部分绣球属野生种原有的分类观点,各亚组内物种间遗传距离差异较大,物种间和亚组间缺乏基因流使得种间自然杂交困难。S.M.Reed[12]等利用上述SSR分子标记引物鉴定了36个绣球品种及杂交种和野生圆锥绣球亲缘关系,发现虽然很多品种源自同一亲本,但是各个品种之间相似性较低;早花与夏花品种聚成2类,早花品种间遗传关系较远,推测是由于这些早花品种的来源地不同而造成的。N.Sukhikh等[30]从利用38个SSR分子标记对39个绣球品种进行亲缘关系分析,其中9个SSR引物具有显著的多态性,通过聚类分析将39份材料分为3大类,其中2类全部为圆球形(Mophead)花序品种,另一类全部为蕾丝帽(Lacecap)花序品种,且蕾丝帽品种间亲缘关系更近。S.M.Reed等[31]用39个SSR标记鉴定114个绣球品种,聚类分析发现5个多次开花品种与4个单次开花品种具有较高遗传相似性,多次开花品种Hydrangeamacrophylla‘Early Sensation’与其他多次开花品种亲缘关系较远,推测可能是杂交中的亲本。X.B.Wu等[32]通过GBS(Genotyping by Sequencing)技术,在82个绣球品种中找到5 803个SNP位点,利用种群结构分析(population structure analysis)将82个品种聚类成了3组,一组全部为粗齿绣球(Hydrangeamacrophyllassp.serrata)栽培品种,另外2组为大花绣球(Hydrangeamacrophyllassp.macrophylla)栽培品种。早期研究认为粗齿绣球(Hydrangeamacrophyllassp.serrata)与大花绣球(Hydrangeamacrophyllassp.macrophylla)在系统分类学上属于并列关系[33],但是X.B.Wu等[32]根据5 803个SNP位点进行系统发生分析认为应将粗齿绣球归类为绣球的亚种,与部分学者[12-13]观点一致。J.T.Lindstorm等[15]使用11个RAPD引物将7个绣球品种分成了2个聚类,聚类分析发现7个品种遗传距离都很相近,最大遗传距离的2物种相似度可达到88%。Q.Song等[34]筛选了10条ISSR引物对9组近缘无性系绣球进行分子标记研究,10条引物共产生207条条带,其中多态性条带171条,占总条带的82.61%。绣球品种Hydrangeamacrophylla‘Twist N Shout’是最具代表性的蕾丝帽品种之一,但其中1个芽突变体缺失了蕾丝帽这一典型特征。然而ISSR标记显示,该无蕾丝帽花序的芽突变体与‘Twist N Shout’只有3条条带不同。此外,单瓣花品种与重瓣花品种的ISSR分子标记条带有显著差异。
2 分子标记辅助育种
2.1 杂交子代鉴定
S.M.Reed等[35]使用常山与绣球作为亲本进行杂交,通过SSR分子鉴定获得了真实杂种,杂交子代表型介于双亲之间并可产生大量花粉。J.H.Kardos等[36]使用绣球与伞花绣球(Hydrangeaangustipelata)杂交,利用12个SSR引物进行鉴定,结合表型对比分析确认杂交后代真实性。N.Kudo等[37]用绣球与耐寒绣球杂交,通过子叶培养获得杂交后代,采用RAPD鉴定了后代[38]。此外还采用中国绣球Hydrangeascandensssp.chinensis(现名称修订为Hydrangeachinensis)与绣球杂交,用2对RAPD引物鉴定出其为真实杂种[39]。S.M.Reed等[40]以绣球Hydrangeamacrophylla‘Kardinal’为母本,以圆锥绣球Hydrangeapaniculata‘Brussels Lace’为父本进行杂交,子代经RAPD验证为真实杂种后代。后代在叶片和被毛方面与父本圆锥绣球相似,比母本对白粉病抗性更强。T.A.Rinehart等[41]用SSR验证了上述杂交后代的真实性,并发现除了性状介于双亲之间,染色体数量有时也会介于双亲之间。K.D.Jones等[42]采用人工相互授粉进行绣球属杂交试验,使用RAPD验证杂种后代真实性,鉴定了以耐寒绣球Hydrangeaarborescens‘Dardom’为母本,总苞绣球(Hydrangeainvolucrata)为父本的杂交组合得到的杂交后代,杂交后代染色体数量(2n=34)介于双亲之间。M.Cai等[43]使用绣球Hydrangeamacrophylla‘Diamond’为母本与耐寒绣球Hydrangeaarborescens‘Annabelle’为父本进行杂交,通过胚挽救获得F1代,对后代进行SSR分子标记鉴定证明为真实杂交后代。
分子标记技术在鉴定子代真实性的同时也可以通过观察引物在杂交后代中产生片段的来源,可以判断双亲之间多样性的高低。M.Wiedemann等[44]使用粗齿绣球、山绣球与2个常山品种进行属间杂交,使用RAPD测序验证后代真实性,引物共产生30个片段,11个片段来自母本,13个片段来自父本,说明双亲多样性较高。L.Crespel等[45]使用马桑绣球种内亲缘关系最远的2品种进行杂交,对获得的10个后代进行ISSR分析标记证明后代为真实后代,亲本之间多态性为96.9%,后代之间多态性标记为60%,表明亲本杂合度很高。
2.2 遗传连锁图谱和观赏性状关联标记
绣球的遗传连锁图谱研究起步相对较晚。2018年,T.A.Rinehart等[46]采用转录组测序获得1 535个SNP,其中779个位点具有多态性,但是由于部分位点在图谱中位置相同和预期分离极为不同等原因,标记密度不够充分,不足以构建连锁图谱。因此,T.A.Rinehart等[41]对绣球Hydrangeamacrophylla‘Bailmer’和Hydrangeamacrophylla‘Vetichii’杂交获得的F1代进行基因分型测序(GBS),以期待找到更多SNP标记来完成图谱。T.Waki等[47]以‘Kirakiraboshi’和‘Frau yoshimi’杂交F2代的93株材料为作图群体,通过NGS(Next-generation DNA Sequencing)技术获得672个SSR标记,构建了第1张绣球遗传连锁图谱,包含147个标记和18个连锁群,总图距980 cm,并将花序类型INF位点定位到连锁群4,并获得2个标记,分别可解释93.5%和96.3%的花序表型变异。C.Tränkner等[48]使用RAD(Restriction-site-associated DNA Sequencing)结合BSA(Bulk Sequence Analysis)混池测序方法对蕾丝帽花序和圆球型花序F1杂交后代进行测序,获得2个与INF位点紧密关联的标记,每个标记可解释99.7%的花序表型变异。X.Wu等[49]以82个绣球品种为材料,使用了5 803个SNP标记对绣球花序类型和多次开花性状开展了全基因组关联分析(Genome-wide Association Study,GWAS),筛选到1个与花序类型相关标记,最多解释65.5%表型变异,经转换为CAPS标记,表型预测准确率达100%;同时获得了23个与多次开花连锁的分子标记。K.Nashima等[50]在绣球全基因组数据的基础上,通过ddRAD简化基因组测序技术构建了高密度遗传连锁图谱,最长为2 944.5 cm,包含4 071个SNP标记和18个连锁群;将重瓣性状位点djo定位到CHR17连锁群33.7~43.8 Mb,将另一位点dsu定位到CHR04连锁群,开发了J01和S01标记用于快速鉴别重瓣表型。X.Wu等[51]以绣球Hydrangeamacrophylla‘Veitichii’和Hydrangeamacrophylla‘Endless Summer’为亲本构建F1群体,利用267个由转录组获得的SSR多态性分子标记和3 923个由GBS获得的SNP多态性标记共4 190个标记进行父本、母本以及共同遗传图谱的构建,共同遗传图谱包含1 767个定位标记(146 SSRs and 1 621 SNPs),长度为1 383.4 cm,由18个连锁群组成,平均映射区间为0.8 cm。
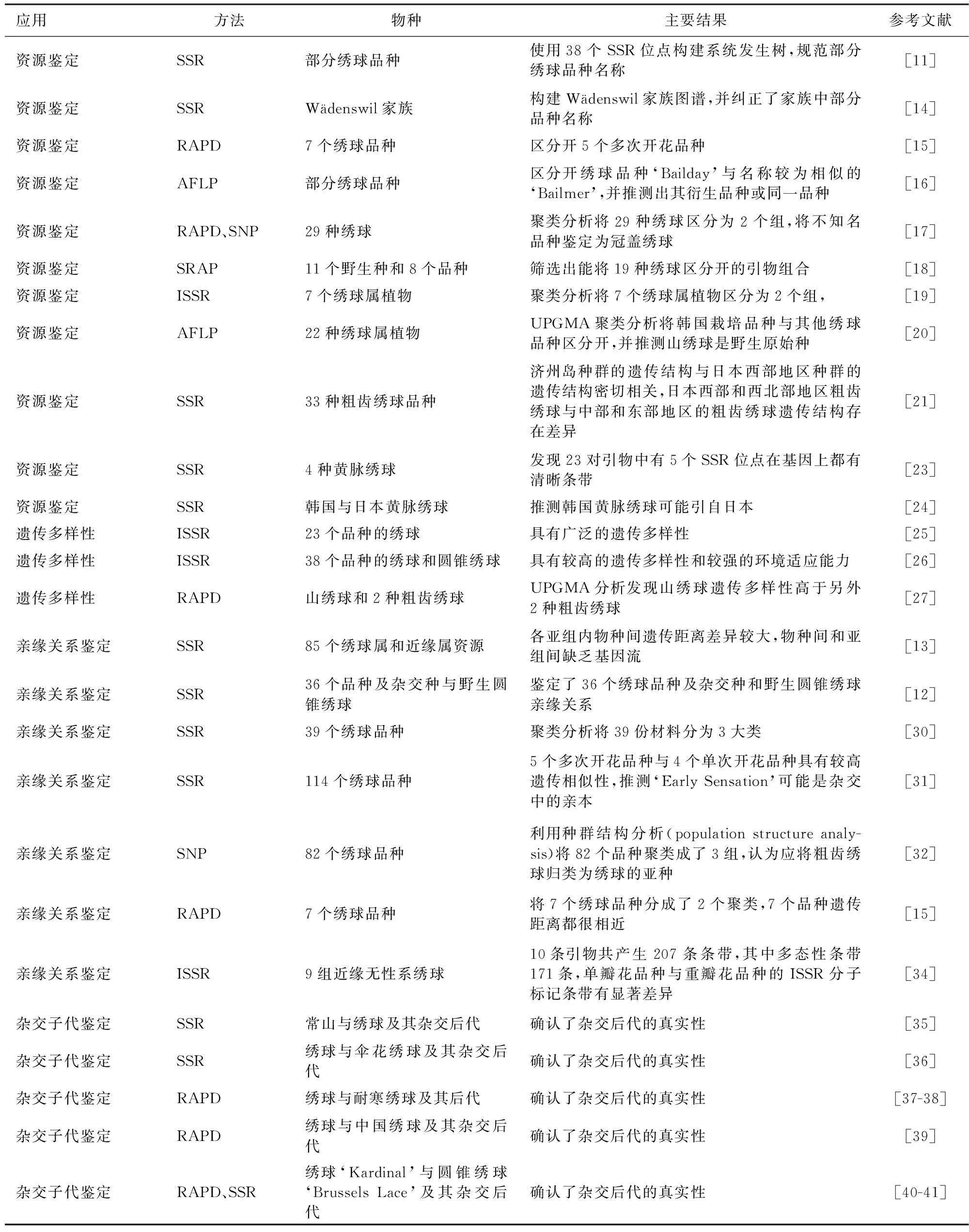
表2 分子标记技术在绣球属研究应用情况Table 2 Application of molecular markers in research and use of Hydrangea

续表2
3 结论与讨论
分子标记已大量应用于绣球属植物研究中,其中资源评价使用了RAPD、AFLP、ISSR、SSR、SRAP、SNP等分子标记技术,辅助育种研究常用RAPD、AFLP、ISSR、SSR等分子标记,这些标记均有其各自的优点和局限性。随着新一代测序技术的快速发展和测序成本的降低,基于转录组、简化基因组和全基因组数据的SSR、SNP、InDel等分子标记将大规模应用于绣球研究中,在提高资源评价与鉴定的准确性、发掘与观赏性状紧密连锁分子标记、提高育种后代选择效率、缩短育种周期等方面发挥重要作用。基于组学数据开发的分子标记,已在绣球遗传连锁图谱构建,花色、花序类型、重瓣性等重要观赏性状遗传规律研究中获得了初步成果[46-55]。未来,随着绣球全基因组序列的公布[47-51],全基因组关联分析、全基因组选择(Genome Selection)等高效分析策略将广泛应用于绣球复杂性状解析、紧密连锁分子标记开发和基因精细定位中,结合多组学数据,研究者将深入了解绣球花朵变色、花序类型、花朵重瓣、攀援和花香等性状形成的分子机理和遗传规律,为实现绣球分子设计育种奠定基础。
