云南白族民家支系18 个常染色体STR 位点的遗传多态性
2022-07-30张寿勋徐冲冲姜焰凌韩建利杨晓佩卢晓筱钟树荣
魏 巍 ,张寿勋 ,徐冲冲 ,姜焰凌 ,李 屹 ,杨 朔 ,韩建利 ,黎 宽 ,赵 斐 ,杨晓佩 ,张 柠 ,卢晓筱 ,钟树荣
(1)昆明医科大学法医学院,云南 昆明 650500;2)昆明医科大学公共卫生学院,云南 昆明 650500;3)大理白族自治州公安司法鉴定中心,云南 大理 671000;4)江西神州司法鉴定中心,江西 南昌 330000;5)楚雄医药高等专科学校,云南 楚雄 675005;6)北京通达首诚司法鉴定所,北京 100089;7)昆明医科大学司法鉴定中心,云南 昆明 650500)
白族是中国人口占比位列第13 的少数民族,主要分布在云南中西部、贵州毕节、湖南张家界、湘西、四川凉山等地,其中云南是白族人口最多的省份。在云南,白族主要分布于大理州,该州是白族的发祥地、祖籍地和主要聚集地。根据2020 年第七次人口普查,白族人口约为209.154 3万人,其中中国80%以上的白族人口居住在云南省大理洱海地区(http://www.stats.gov.cn/tjsj/ndsj/2021/indexch.htm)。
受地域和风俗习惯的影响,白族可以分为3个分支:民家、勒墨和那马。白族勒墨和那马支系主要分布在怒江流域的兰坪县、维西县、福贡县等地,经济文化水平与怒族和傈僳族相近。白族民家支系则是白族人口最多的一支,主要居住在云南大理的洱海地区,深受中国汉文化的影响,具有较高的经济文化水平[1]。
由于同一STR(short tandem repeat,STR)基因座中相同的等位基因在不同群体间基因频率分布存在差异,导致亲权鉴定和个体识别中关键法医学参数的计算值亦有不同,如亲权指数(paternity index,PI)和随机匹配概率(matching probability,Pm)等。因此,对不同种族和民族的人群进行基础遗传学数据的调查是非常有必要的。本研究选取2 323 名白族民家支系健康无关个体,对云南白族民家支系进行群体遗传调查,以期建立白族民家支系的法医遗传学参数,为司法鉴定工作中的亲子鉴定和个体识别提供基础数据。
1 材料与方法
1.1 DNA 提取、PCR 扩增和STR 分型
在签署知情同意书后,在云南省大理白族自治州洱源县和剑川县采集了2 323 名无亲属关系健康白族个体的血样或口腔拭子。采用DNATyperTM19(公安部物证鉴定中心,中国)试剂盒,使用GeneAmpPCR System 9700 PCR 仪(AB公司,美国)对18 个常染色体STR 基因座(D3S1358、D13S317、D16S539、Penta E、D18S51、CSF1PO、TH01、D2S1338、vWA、D21S11、D7S820、D5S818、D12S391、TPOX、D19S433、D8S1179、FGA和D6S1043)进行直接PCR 复合扩增,同时设立灭菌纯水为阴性对照样本,9947A为阳性对照样本。PCR 复合扩增所得到的产物使用AB 3130XL 自动遗传分析仪(AB 公司,美国)进行毛细管电泳分离,用GeneMapper ID-X 1.5软件(AB 公司,美国)进行STR 分型。根据国际法庭科学遗传学会(international society for forensic genetics,ISFG)的规定对扩增产物进行分型和命名[2-3]。
1.2 统计学处理
使用Modified-Powerstats 软件[4]统计等位基因频率(allele frequency,AF),并进行Hardy-Weinberg 平衡(HWE)检验,采用Bonferroni 多重校正,以P<0.002 8 为差异有统计学意义。计算随机匹配概率(matching probability,Pm)、观测值杂合度(observed heterozygosity,Ho)、多态性信息含量(polymorphism information content,PIC)、个体识别能力(discrimination power,DP)、非父排除概率(probability of exclusion,PE)、亲权指 数(paternity index,PI)等法医学参数。使用Arlequin v3.5 软件计算遗传分化指数(Fst)和P值[5],采用Bonferroni 多重校正,以P<0.003 3 为差异有统计学意义。采用Phylip 3.698 软件包计算白族民家支系与21 个已发表人群的群体遗传距离(Nei’ s)(http://evolution.genetics.washington.edu/phylip.html)。基于Nei’ s 遗传距离,应用Mega 7.0 软件[6]构建相邻连接(neighbour-joining,NJ)系统发育树。使用SPSS v24.0 统计软件(IBM 公司,美国),进行多维尺度(multidimensional scaling,MDS)分析。
2 结果
2.1 18 个STR 基因座的等位基因频率和法医学相关参数
白族民家支系2 323 名无亲属关系个体18 个STR 基因座的等位基因频率和法医学相关参数,见表1。在上述18 个STR 基因座中共检出230 个等位基因和1 073 个基因型。等位基因频率分布在0.000 2~0.509 3 之间。等位基因数量最少为7 个(TPOX),最多为22 个(Penta E)。经Bonferroni校正后,有17 个基因座等位基因频率和基因型频率符合Hardy-Weinberg 平衡(P>0.05/ 18=0.002 8),D13S317 基因座不符合Hardy-Weinberg 平衡(P=0.000 8)。随机匹配概率(Pm)的范围为0.016 6~0.202 3。观测值杂合度(Ho)的范围为 0.622 0~0.899 3。多态性信息含量(PIC)在0.564 1~0.897 7之间。个体识别能力(DP)在0.797 7~0.983 4 之间,累积个人识别能力(cumulative discrimination power,CDP)达到0.999 999 999。非父排除概率(probability of exclusion,PE)在0.318 2~0.793 9 之间,累积非父排除率(cumulative probability of exclusion,CPE)达到0.999 999 940 55。
2.2 遗传分化指数(Fst)和P 值
白族民家支系与21 个已发表的人群[7-26]在共有的15 个STR 基因座之间的遗传分化指数(Fst)和P值,见 表2 。经Bonferroni 校正后(P<0.05/15≈0.003 3),白族民家支系与喀什地区维吾尔族、云南佤族和云南地区越南人群在14 个STR 基因座上存在差异(P<0.003 3),与广西侗族和印度人群在11 个STR 基因座上存在差异(P<0.003 3),与云南哈尼族在9 个STR 基因座上存在差异(P<0.003 3),与海南黎族和昌都藏族在8个STR 基因座上存在差异(P<0.003 3),与伊犁地区维吾尔族在7 个STR 基因座上存在差异(P<0.003 3),与日本人群在6 个STR 基因座上存在差异(P<0.003 3),与云南汉族在3 个STR 基因座上存在差异(P<0.003 3),与云南布朗族和长沙汉族在2 个STR 基因座上存在差异(P<0.003 3),与四川汉族和纳西族在1 个STR 基因座上存在差异(P<0.003 3),与贵州仡佬族、彝族、宁夏回族、拉萨藏族、怒江白族和大理白族在各基因座上均无统计学差异(P>0.003 3)。

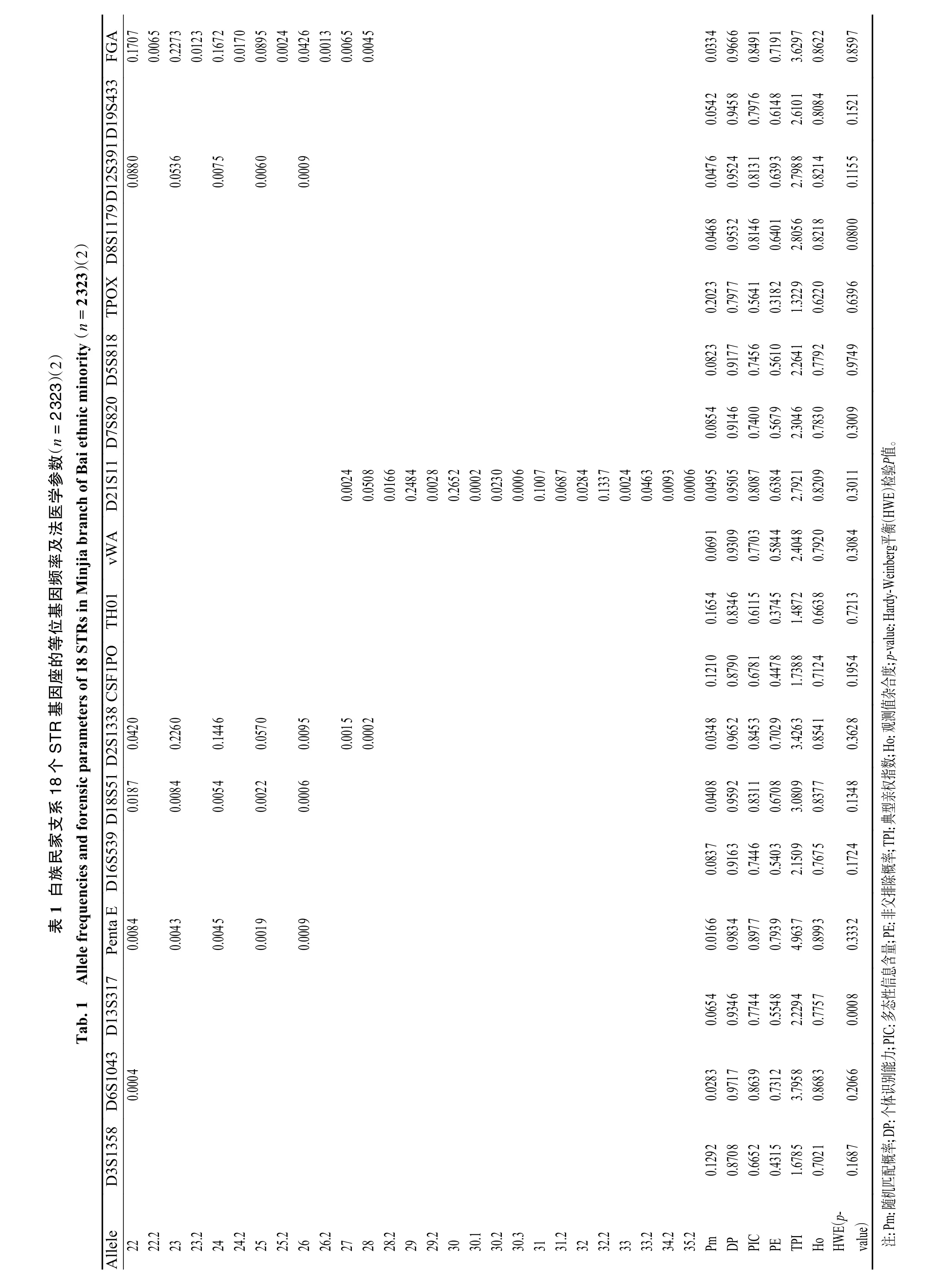

表2 白族民家支系与21 个已发表人群的Fst 和P 值(1)Tab.2 Pairwise Fst and P values between Minjia branch of Baiethnic minority and 21 published populations(1)

表2 白族民家支系与21 个已发表人群的Fst 和P 值(2)Tab.2 Pairwise Fst and P values between Minjia branch of Bai ethnic minority and 21 published populations(2)

表2 白族民家支系与21 个已发表人群的Fst 和P 值(3)Tab.2 Pairwise Fst and P values between Minjia branch of Bai ethnic minority and 21 published populations(3)
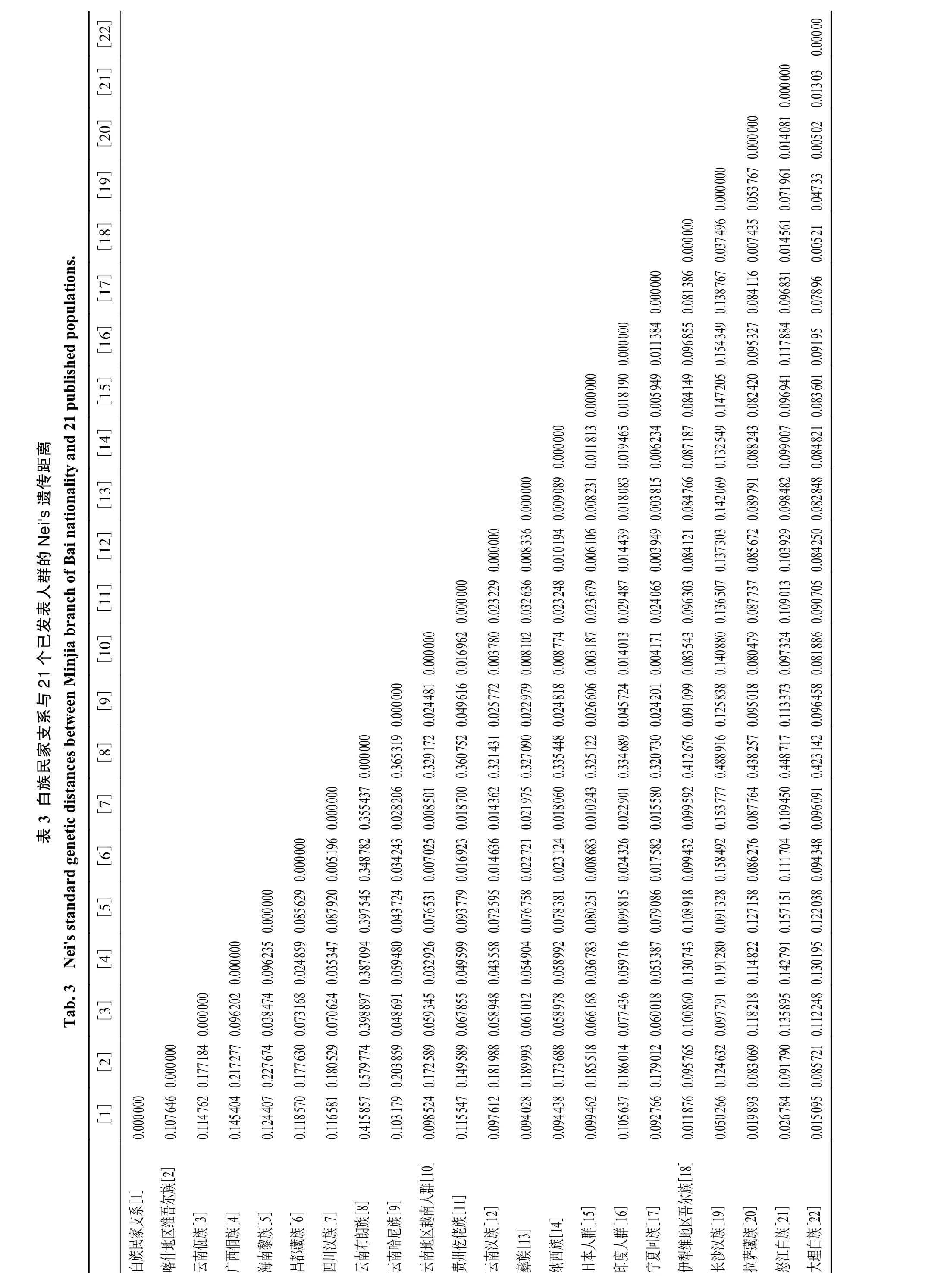
2.3 Nei’s 遗传距离
根据个体之间共享等位基因片段的数量计算Nei’s 遗传距离。白族民家支系与云南布朗族和云南佤族相距较远,而与大理白族和怒江白族相距较近,见表3。
2.4 NJ 系统发育树和多维尺度分析(MDS)
为进一步探索白族民家支系与其他人群的亲缘关系,基于Nei’s 遗传距离矩阵笔者构建了NJ 系统发育树,见图1。从NJ 系统发育树可以看出,白族民家支系与大理白族和怒江白族相距较近,而与云南布朗族和云南佤族相距较远。此外,基于Nei’s 遗传距离矩阵笔者还进行了多维尺度分析(MDS),见图2。从MDS 分析同样可以看出,白族民家支系与大理白族和怒江白族相距较近,而与云南布朗族和云南佤族相距较远。

图1 白族民家支系与21 个已发表人群的系统发育树Fig.1 The phylogenetic tree showing the relationship of the Minjia branch of Bai nationality and 21 published populations
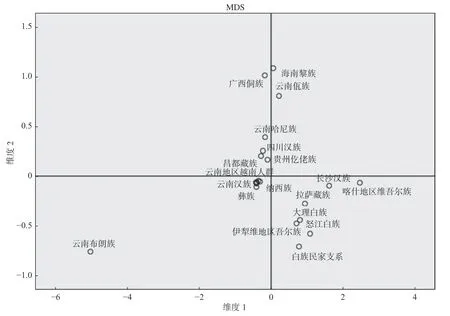
图2 白族民家支系与21 个已发表人群的MDS 分析(基于Nei’s 遗传距离矩阵)Fig.2 Multidimensional scaling(MDS)plot based on Nei’s standard genetic distances matrix among Minjia branch of Bai nationality and 21 published populations
3 讨论
Ho 和PIC 等法医学参数常被用来衡量STR基因座的多态性程度,并推测其法医学应用价值。在本次针对白族民家支系的调查研究中发现,所选的18 个STR 基因座的PIC 在0.564 1~0.897 7之间,PIC>0.5,遗传标记具有高度的多态信息[27]。在个体识别和亲子鉴定中,CDP 和CPE参数的计算是极其重要的。在针对白族民家支系的调查中发现,本研究所选的18 个常染色体STR 基因座的CDP 达0.999 999 999,具有较强的个体识别能力;CPE达0.999 999 940 55,大于0.999 9,在亲子鉴定中具有极强的系统效能[28-29]。本研究所获得的白族民家支系18 个常染色体STR 基因座的法医学参数和群体遗传学数据具有极高的应用价值。
云南佤族和云南布朗族属于南亚语系中的孟高棉语族[30]。对Y 染色体的研究发现,云南孟高棉语族起源于东亚南部[31]。而白族民家支系则属于汉藏语系的藏缅语族[32]。戴庆厦等[32]从语言学的角度发现,在中国南方分布的藏缅语族人群起源于中国北方的黄河流域。Wang 等[33]对Y 染色体的研究表明,中国南方分布的藏缅语族人群是由北方高原黄河流域迁徙而来。上述研究提示,云南佤族、云南布朗族与白族民家支系在起源上存在着显著的地理差异。与本研究结果,白族民家支系与云南佤族、云南布朗族在遗传上相距较远相一致。
白族的3 个主要分支中,勒墨支系于15 世纪之前就已经从大理白族的主体中迁徙出来,最终聚居于怒江流域的兰坪县[34]。而民家支系主要聚居于大理州[1]。上述史料与研究提示,白族民家支系与属于勒墨支系的怒江白族存在一定的差异。
本研究报道了白族民家支系18 个常染色体STR 基因座的等位基因频率和法医学参数等。研究结果提示,上述18 个常染色体STR 基因座在白族民家支系群体中具有较高的遗传多态性,可用于个体识别、亲子鉴定以及群体遗传结构的研究。