Influence of biological sex and exercise on murine cardiac metabolism
2022-07-29KyleFulghumHelenCollinsStevenJonesBrdfordHill
Kyle Fulghum,Helen E.Collins,Steven P.Jones,Brdford G.Hill,*
a Diabetes and Obesity Center,Department of Medicine,Division of Environmental Medicine,Christina Lee Brown Envirome Institute,University of Louisville,Louisville,KY 40202,USA
b Department of Physiology,University of Louisville,Louisville,KY 40202,USA
Abstract Although the structural and functional effects of exercise on the heart are well established,the metabolic changes that occur in the heart during and after exercise remain unclear.In this study,we used metabolomics to assess time-dependent changes in the murine cardiac metabolome following 1 session of treadmill exercise.After the exercise bout,we also recorded blood lactate,glucose,and ketone body levels and measured cardiac mitochondrial respiration.In both male and female mice,moderate-and high-intensity exercise acutely increased blood lactate levels.In both sexes,low-and moderate-intensity exercise augmented circulating 3-hydroxybutryrate levels immediately after the exercise bout;however,only in female mice did high-intensity exercise increase 3-hydroxybutyrate levels,with significant increases occurring 1 h after the exercise session. Untargeted metabolomics analyses of sedentary female and male hearts suggest considerable sex-dependent differences in basal cardiac metabolite levels,with female hearts characterized by higher levels of pantothenate,pyridoxamine,homoarginine,tryptophan,and several glycerophospholipid and sphingomyelin species and lower levels of numerous metabolites, including acetyl coenzyme A, glucuronate, gulonate,hydroxyproline, prolyl-hydroxyproline, carnosine, anserine, and carnitinylated and glycinated species, as compared with male hearts. Immediately after a bout of treadmill exercise, both male and female hearts had higher levels of corticosterone; however, female mice showed more extensive exercise-induced changes in the cardiac metabolome,characterized by significant,time-dependent changes in amino acids(e.g.,serine,alanine,tyrosine,tryptophan,branched-chain amino acids)and the ketone body 3-hydroxybutyrate.Results from experiments using isolated cardiac mitochondria suggest that high-intensity treadmill exercise does not acutely affect respiration or mitochondrial coupling;however,female cardiac mitochondria demonstrate generally higher adenosine diphosphate sensitivity compared with male cardiac mitochondria. Collectively,these findings in mice reveal key sex-dependent differences in cardiac metabolism and suggest that the metabolic network in the female heart is more responsive to physiological stress caused by exercise.
Keywords: Ketone bodies;Metabolomics;Mitochondria;Physical activity;Sex differences
1. Introduction
Exercise presents a major challenge to systemic metabolic homeostasis.1Moderate- to high-intensity exercise is particularly demanding because maintenance of high levels of physical and cardiac work requires higher oxygen and substrate utilization in skeletal muscle and the heart.Although it is known that the metabolic requirements of tissues vary as a product of exercise intensity and duration,1-3it remains unclear how exercise affects the many different pathways of metabolism.This is important to understand because recent evidence suggests that transient changes in metabolism are critical for adaptive responses to exercise.2,4In particular,little is understood about how heart metabolism changes during and after exercise.
During exercise,cardiac contractile power and oxygen consumption can increase by up to 10-fold above resting rates.5,6This increase in myocardial workload is accompanied by increased catabolism of several circulating substrates, including glucose, lactate, and fatty acids.4Although moderateintensity exercise has been associated with elevations in myocardial glucose uptake and oxidation, elevations in circulating concentrations of competing substrates such as lactate and fatty acids may decrease glucose catabolism.4,7Regular exercise also promotes adaptive metabolic remodeling in the heart.Perfused heart studies indicate that acclimation to an exercise regimen is associated with increases in the rates of basal glycolysis,8glucose oxidation, and fat oxidation9in mice;however,cardiac glycolysis has been suggested to be lower in exercise-adapted rats,despite higher myocardial palmitate and glucose oxidation.10Similarly,the effects of exercise on mitochondrial respiration are not consistent:although some studies show that acute or long-term exercise training increases cardiac mitochondrial respiration,9,11-14other studies suggest that exercise either does not affect or may even decrease state 3 respiration in cardiac mitochondria.15-17The reasons for these discrepancies remain unclear, but they are likely due to technical differences, such as model-dependent differences or variations in study design.
Because adaptive changes are triggered early in an exercise training program,2we examined the changes in metabolism that occur with a single bout of exercise.Using metabolomics,we assessed how exercise influences the cardiac metabolome at different times following an exercise session in male and female mice. We also examined whether or not exercise acutely influences cardiac mitochondrial respiration. Our data suggest that the metabolic response to exercise is different in male versus female murine hearts and that, even basally,female hearts have remarkably different metabolite profiles than male hearts.Moreover,our data suggest that a single bout of high-intensity exercise has minimal effects on mitochondrial respiration but that female cardiac mitochondria have intrinsically higher adenosine diphosphate (ADP) sensitivity than those in male mice.
2. Methods
2.1. Experimental animals
All procedures were approved by the Institutional Animal Care and Use Committee at the University of Louisville.Adult male and female FVB/NJ mice were purchased from the Jackson Laboratory (Bar Harbor, ME, USA) and maintained on 12 h:12 h(light:dark)cycle,with both chow and water provided ad libitum unless fasted (6 h) for untargeted metabolomics experiments. All mice were 13 weeks of age at the time of exercise experimentation, and all mice were both exercised and euthanized at the same time of day. For metabolomics studies,all mice were fasted for 6 h prior to euthanasia. Upon completion of each experiment, mice were anesthetized with sodium pentobarbital (Sigma-Aldrich, St. Louis, MO, USA;150 mg/kg,i.p.),and mice were euthanized via excision of the heart, which was freeze-clamped in situ. Additional tissues were harvested following euthanasia. These procedures are consistent with the American Veterinary Medical Association Guidelines on Euthanasia.18
2.2. Exercise training protocol
Mice were acclimated to forced treadmill running and exercised as previously described.19Briefly, mice were exercised to exhaustion to determine initial exercise capacity. Training intensity was then determined from this initial capacity test:low-intensity exercise was carried out at 12 m/min for 40 min with 10˚incline (55%-60% of the initial exercise capacity);moderate-intensity exercise was carried out at 19.1 m/min for 40 min with 10˚incline(75%of the initial exercise capacity);and the exercise capacity test served as a high-intensity,exhaustive bout of exercise.In this exercise capacity test,mice run to exhaustion with increasing treadmill speed and incline,as described previously.8,19Blood glucose, lactate, and 3-hydroxybutyrate measurements were acquired from tail blood before and after the exercise bout using an Accu-Check Aviva meter(Roche,San Francisco,CA,USA),a Lactate Plus meter(Nova Biomedical, Waltham, MA, USA), and a Keto-Mojo meter (Keto-Mojo, Amsterdam Duivendrecht, Netherlands),respectively. Sedentary control mice were subjected to the same conditions as exercised mice but were sat on a treadmill with speed=0 m/s. For metabolomics studies, all mice were fasted for 6 h prior to euthanasia.
2.3. Metabolomics
Hearts were freeze-clamped in situ using liquid nitrogencooled Wollenberger Tongs and powdered under liquid nitrogen. The samples were then prepared by Metabolon using an automated MicroLab STAR system (Hamilton Company,Reno, NV, USA). First, tissue homogenates were made in water at a ratio of 5 μL per mg of tissue. For quality control,several recovery standards were added prior to the first step in the extraction process. To remove protein, dissociate small molecules bound to protein or trapped in the precipitated protein matrix, and to recover chemically diverse metabolites,proteins were precipitated with methanol (VWR, Radnor, PA,USA;final concentration 80%v/v)under vigorous shaking for 2 min (Glen Mills GenoGrinder 2000, Clifton, NJ, USA) followed by centrifugation. For quality assurance and control, a pooled matrix sample was generated by taking a small volume of each experimental sample to serve as a technical replicate throughout the data set.Extracted water samples served as process blanks. A cocktail of standards known not to interfere with the measurement of endogenous compounds was spiked into every analyzed sample, allowing instrument performance monitoring and aiding chromatographic alignment.
The extract was divided into fractions for analysis by reverse-phase/ultra-performance liquid chromatography-tandem mass spectrometry with positive ion mode electrospray ionization, by reverse-phase/ultra-performance liquid chromatography-tandem mass spectrometry with negative ion mode electrospray ionization, and by hydrophilic interaction chromatography/ultra-performance liquid chromatography-tandem mass spectrometry with negative ion mode electrospray ionization. Samples were placed briefly on a TurboVap (Zymark,Clackamas, OR, USA) to remove the organic solvent. All methods utilized a Waters ACQUITY UPLC (Milford, MA,USA) and a ThermoFisher Scientific Q-Exactive high resolution/accurate mass spectrometer(Bethlehem,KY,USA)interfaced with a heated electrospray ionization source and Orbitrap mass analyzer operated at 35,000 mass resolution.The sample extract was reconstituted in solvents compatible with each mass spectrometry/mass spectrometry method.Each reconstitution solvent contained a series of standards at fixed concentrations to ensure injection and chromatographic consistency. One aliquot was analyzed using acidic positive ion conditions chromatographically optimized for hydrophilic compounds. In this method, the extract was gradient eluted from a C18 column (Waters UPLC BEH C18-2.1×100 mm,1.7 μm)using water and methanol containing 0.05%perfluoropentanoic acid and 0.1% formic acid. For more hydrophobic compounds, the extract was gradient eluted from the aforementioned C18 column using methanol, acetonitrile, water,0.05% perfluoropentanoic acid, and 0.01% formic acid. Aliquots analyzed using basic negative ion optimized conditions were gradient eluted from a separate column using methanol and water containing 6.5 mM ammonium bicarbonate (pH 8).The last aliquot was analyzed via negative ionization following elution from a hydrophilic interaction chromatography column (Waters UPLC BEH Amide 2.1×150 mm, 1.7 μm)using a gradient consisting of water and acetonitrile with 10 mM ammonium formate(pH 10.8).The MS analysis alternated between MS and data-dependent MSnscans using dynamic exclusion. The scan range covered 70-1000 m/z(mass to charge ratio).
Raw data were extracted, peak-identified, and processed using Metabolon’s proprietary hardware and software(Metabolon Inc., Research Triangle Park,NC, USA).Compounds were identified by comparison to library entries of purified,authenticated standards or recurrent unknown entities, with known retention times/indices, m/z, and chromatographic signatures(including mass spectrometry/mass spectrometry spectral data).Biochemical identifications were based on 3 criteria: retention index within a narrow retention times/indices window of the proposed identification, accurate mass match to the library±10 ppm, and the mass spectrometry/mass spectrometry forward and reverse scores between experimental data and authentic standards. Proprietary visualization and interpretation software (Metabolon Inc., Research Triangle Park, NC, USA)was used to confirm the consistency of peak identification among the various samples. Library matches for each compound were checked for each sample and corrected if necessary.The area under the curve was used for peak quantification.
2.4. Mitochondrial isolation and respiration studies
Hearts from sedentary and exercised mice were isolated and homogenized in 1 mL of isolation buffer (Buffer A: 220 mM mannitol, 70 mM sucrose, 5 mM MOPS, 1 mM EGTA, 0.2%fatty acid-free bovine serum albumin, pH 7.2) using a Potter Elvehjem tube (VWR, Radnor, PA, USA) and a Teflon pestle(VWR, Radnor, PA, USA). The homogenate was centrifuged at 800 g for 10 min at 4˚C. The supernatant was then centrifuged at 10,000 g for 15 min at 4 ˚C to obtain the mitochondrial fraction. The mitochondrial pellets were washed once in 1 mL of isolation buffer and twice in 500 μL of isolation buffer, then resuspended in 400 μL of respiration buffer(120 mM KCl,25 mM sucrose,10 mM HEPES,1 mM MgCl2,5 mM KH2PO4,pH 7.2)for extracellular flux analysis and biochemical assays.Protein concentration was assessed using the Lowry DC Protein Assay kit(Biorad,Hercules,CA,USA).
Mitochondrial respiration was assessed using a Seahorse XF96e analyzer(Agilent,Santa Clara,CA,USA),as described previously.15,20,21For each group, 2.5 μg of mitochondrial protein was suspended in 20 μL of respiration buffer and loaded into 96-well extracellular flux culture microplates.The microplates were centrifuged at 500 g for 3 min at 4˚C, followed by the addition of 160 μL of warm (37˚C) respiration buffer. We used the following substrates to stimulate state 3 respiration (in the presence of 1 mM ADP): 5 mM pyruvate+2.5 mM malate; 5 mM glutamate+2.5 mM malate;10 mM succinate+1 μM rotenone+2.5 mM malate; or 100 μM octanoylcarnitine+2.5 mM malate. State 4 respiration was induced by addition of oligomycin(1 μM,final concentration). The respiratory control ratio was calculated by dividing State 3 respiration rates by State 4 respiration rates.At the end of the respiration assays, 10 μM antimycin A was added to ensure that all oxygen consumption was due to mitochondrial respiration. For mitochondrial ADP sensitivity studies, we measured State 2 respiration using 5 mM pyruvate+2.5 mM malate, then stimulated State 3 respiration with variable concentrations of ADP(0-5 mM).
2.5. Statistical analyses
Original scale data (raw area counts) were analyzed using Metaboanalyst 5.0 software (http://www.metaboanalyst.ca/).22Metabolites with missing values were omitted and the data were filtered by interquartile range, followed by log-transformation.For multiple comparison testing,q values were calculated using a method embedded within the Metaboanalyst software that controlled for the false discovery rate(FDR).23An FDR cutoff of p <0.10 was implemented to assume significance.We used two-way analysis of variance where appropriate and confirmed significance with Bonferroni’s post hoc test as indicated.Statistical significance was assumed where p <0.05.
3. Results
3.1. Acute exercise alters circulating substrate availability
Because exercise intensity could influence circulating substrate levels, we first examined the effects of low-, moderate-,and high-intensity exercise bouts on blood lactate, glucose, and ketone body levels.To accomplish this,we measured circulating substrate levels before(at rest)and after the session of exercise.For low- and moderate-intensity exercise, mice were run for 40 min at 12 m/min or 19.1 m/min(both at a 10˚incline),respectively;for high-intensity exercise,the mice were subjected to an exercise capacity test, where belt speed and incline were increased (up to 30 m/min and 15˚incline) until exhaustion(Fig.1A).As shown in Fig.1B and 1C,distance and work were higher in the moderate-intensity group than the low-intensity group. Due to their relatively early exhaustion during the exercise bout,the high-intensity exercise group ran a shorter distance and performed less work than the moderate-intensity group.
Circulating lactate levels immediately after the exercise bout were not influenced by low- or moderate-intensity exercise, but were significantly increased in both male and female mice by high-intensity exercise(Fig.1D).Although circulating glucose was relatively stable regardless of exercise intensity,we found modestly higher blood glucose levels in male mice subjected to moderate-intensity exercise (Fig. 1E). Interestingly, circulating ketone body levels changed in a manner dependent on exercise intensity: 3-hydroxybutrate was higher immediately after moderate-intensity exercise in male mice and both low-and moderate-intensity exercise in female mice,but it was not changed immediately following high-intensity exercise (Fig. 1F).Collectively, these data indicate that circulating lactate and 3-hydroxybutyrate levels respond in a dissimilar fashion to different exercise intensities and that biological sex could influence this response.
We next examined how circulating substrates change with time after a bout of high-intensity, exhaustive exercise. Blood lactate, glucose, and 3-hydroxybutyrate levels were measured immediately after exercise,1 h following exercise,or 24 h following exercise,with appropriate sedentary controls(Fig.2A).These timepoints were chosen because in our previous studies in male mice,8we found decreased activation of cardiac 6-phosphofructo-2-kinase and elevated levels of cardiac glycogen immediately after exercise, which suggests marked changes in cardiac glucose metabolism.During initial exercise capacity tests, all exercise groups showed similar distance to exhaustion and work performed (Fig. 2B, 2C). As expected,circulating levels of lactate increased in both male and female mice during exercise but returned to normal levels within 1 h following the exercise session (Fig. 2D). While male mice showed no changes in blood glucose concentration following exercise,we observed in female mice a slight reduction in circulating glucose 1 h following exercise(Fig.2E,p=0.05).At this same time(i.e.,1 h after exercise),female mice had significantly higher circulating 3-hydroxybutyrate levels compared with sedentary controls; this indicates a shift in circulating substrate availability, which could affect tissue metabolism.However,there were no significant changes in 3-hydroxybutyrate levels in male mice after a single,exhaustive bout of exercise(Fig.2F).
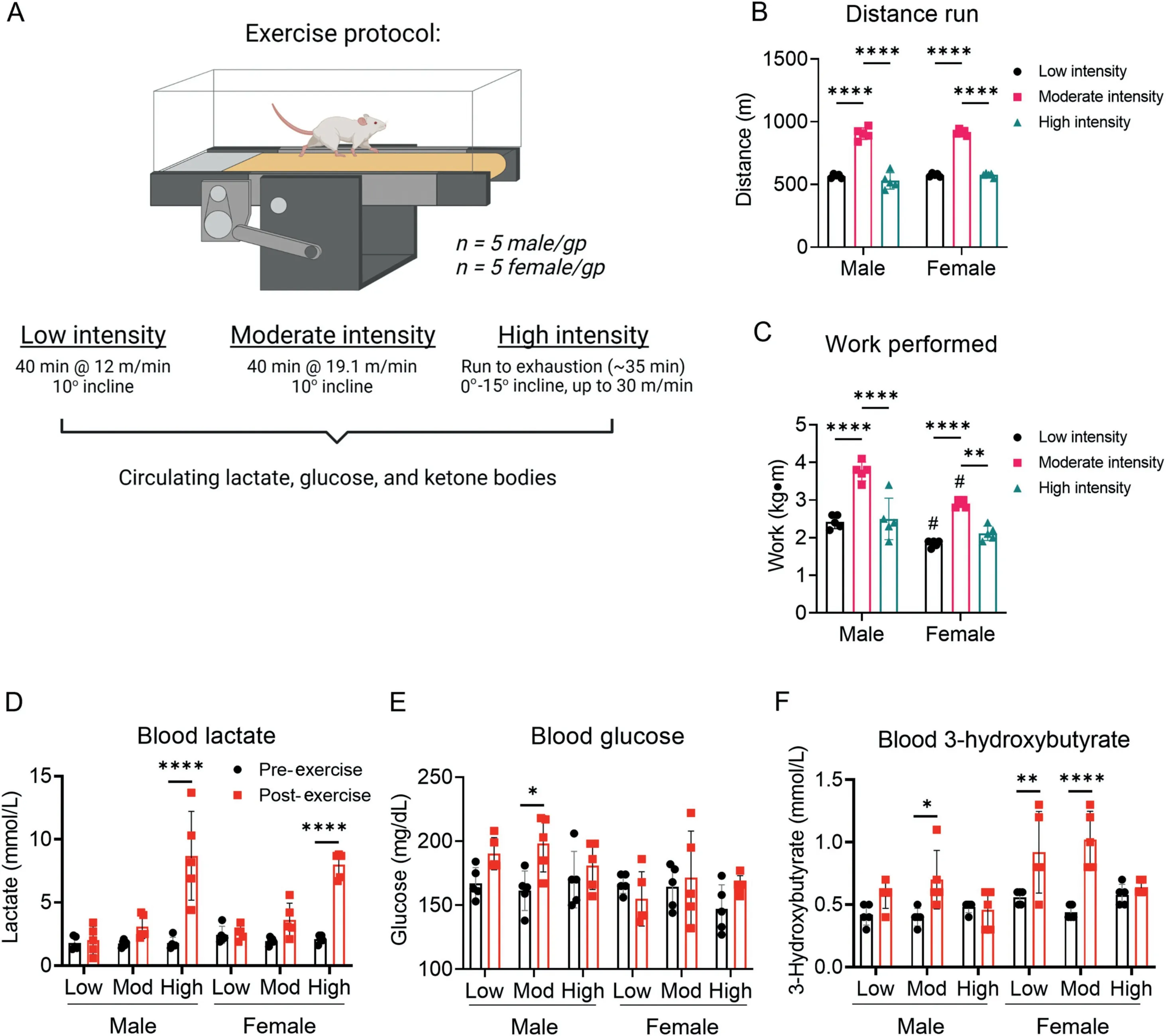
Fig.1. Exercise intensity affects circulating substrate levels.Mice were subjected to low-,moderate-,and high-intensity exercise followed by measurement of circulating substrates immediately after the exercise bout. (A) Schematic of study design; (B) Distance run to exhaustion; (C) Work performed during the exercise session;Measurements of (D) circulating lactate, (E) glucose, and (F) 3-hydroxybutyrate at the end of exercise bout. n=5 mice per group. * p <0.05, ** p <0.01, *** p <0.001,****p <0.0001, #p <0.05 female vs.male,two-way ANOVA with Bonferroni’s post hoc test(B-F).ANOVA=analysis of variance;gp=group.
3.2. Sex-dependent differences in cardiac metabolism
Because the influence of biological sex on metabolic phenotype is not well characterized,we first examined the metabolomic profiles of the sedentary male and female hearts. We found 69 cardiac metabolites that were significantly different(FDR <0.10) in abundance between the sexes (Fig. 3A,Table 1).Partial least-squares discriminant analysis(PLS-DA)showed distinct group clustering by biological sex (Fig. 3B),with several metabolites highlighted in a variable importance in projection (VIP) score plot as being important to the PLSDA model(Fig.3C).Compared with male hearts,these analyses in female hearts revealed higher levels of heme, pantothenate, triethanolamine, and phospholipid species but lower levels of α-hydroxyvalerate, xenobiotics, carnitinylated and glycinated species,carnosine,gulonate,maltotriose,and acetyl coenzyme A(CoA),all of which contributed strongly to group differences.
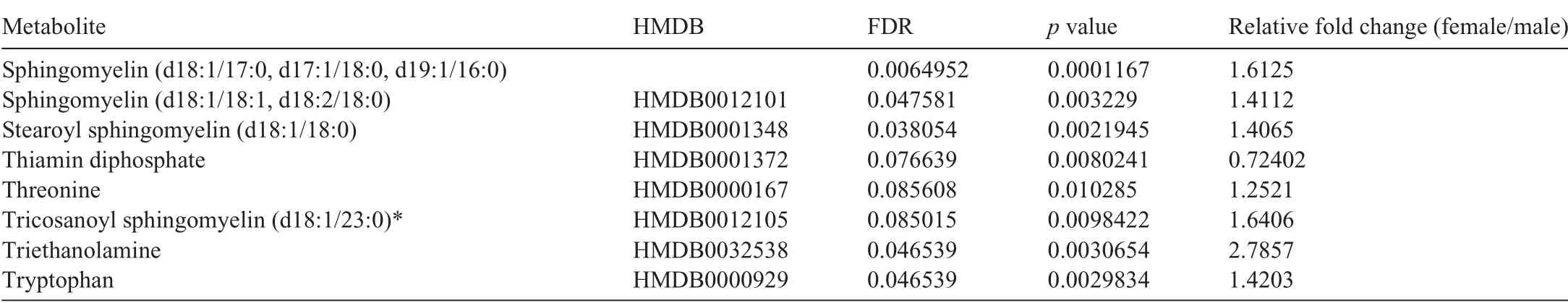
Table 1(Continued)
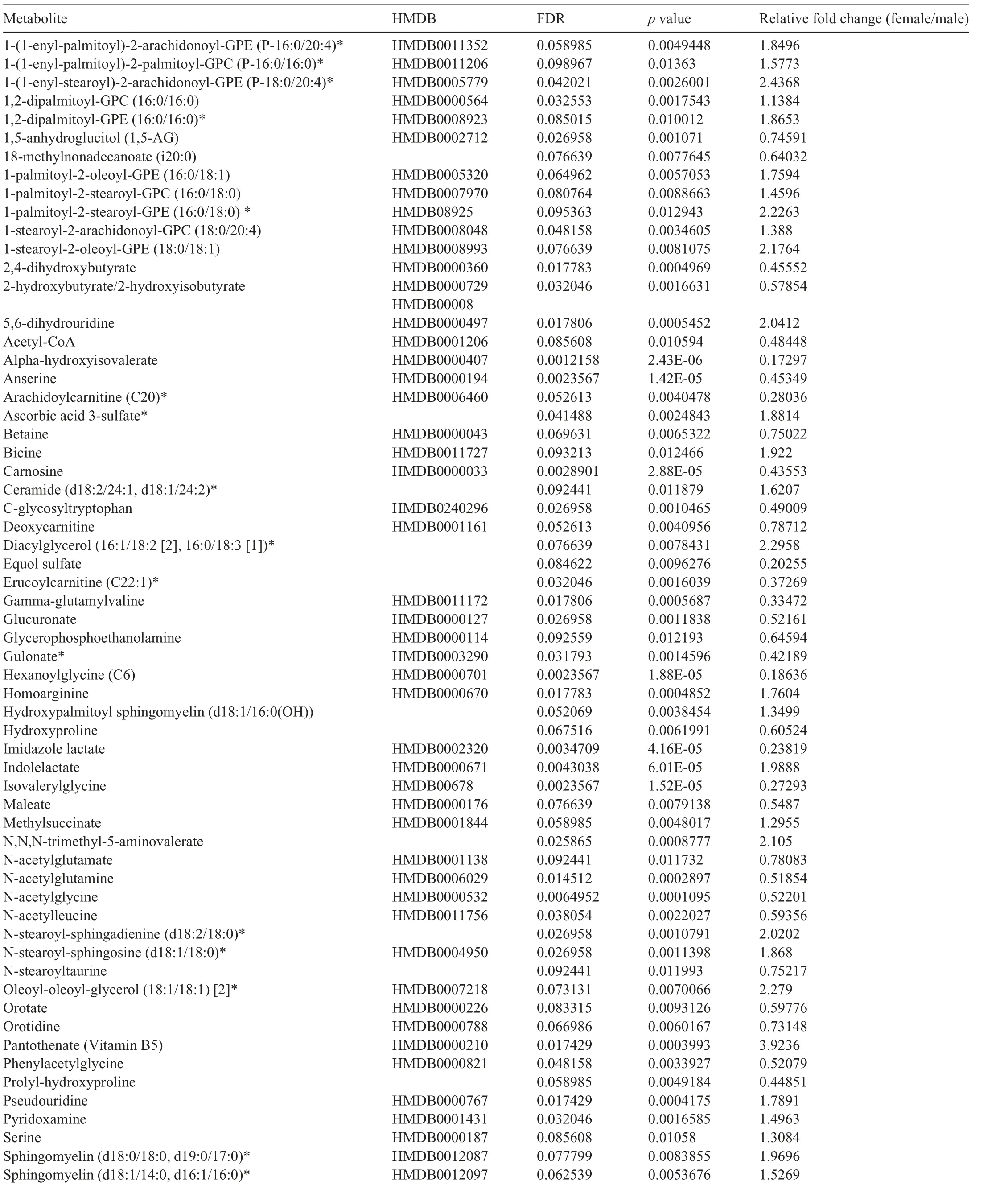
Table 1 Significantly different metabolites in male vs.female mouse hearts under sedentary conditions.
To visualize further baseline sex differences in cardiac metabolite abundance, the 50 most changed metabolites in male and female sedentary hearts are displayed as a heatmap(Fig. 4A). As summarized in the Venn diagram in Fig. 4B,female hearts had higher levels of vasodilatory metabolites,such as homoarginine, and of B vitamins, such as pantothenate and pyridoxamine, as well as higher levels of several sphingomyelin and glycerophospholipid species; however, there were lower levels of some long-chain acylcarnitines (arachidoylcarnitine, erucoylcarnitine) and collagen precursor-breakdown products (hydroxyproline, prolyl-hydroxyproline). Furthermore,glucuronate, gulonate, carnosine, anserine, acetyl CoA, and glycinated metabolites were lower in female hearts. Pathway impact analyses further confirmed the significance of these changes to individual metabolic pathways (Fig. 4C). Collectively, these data suggest that biological sex influences the cardiac metabolome in mice.
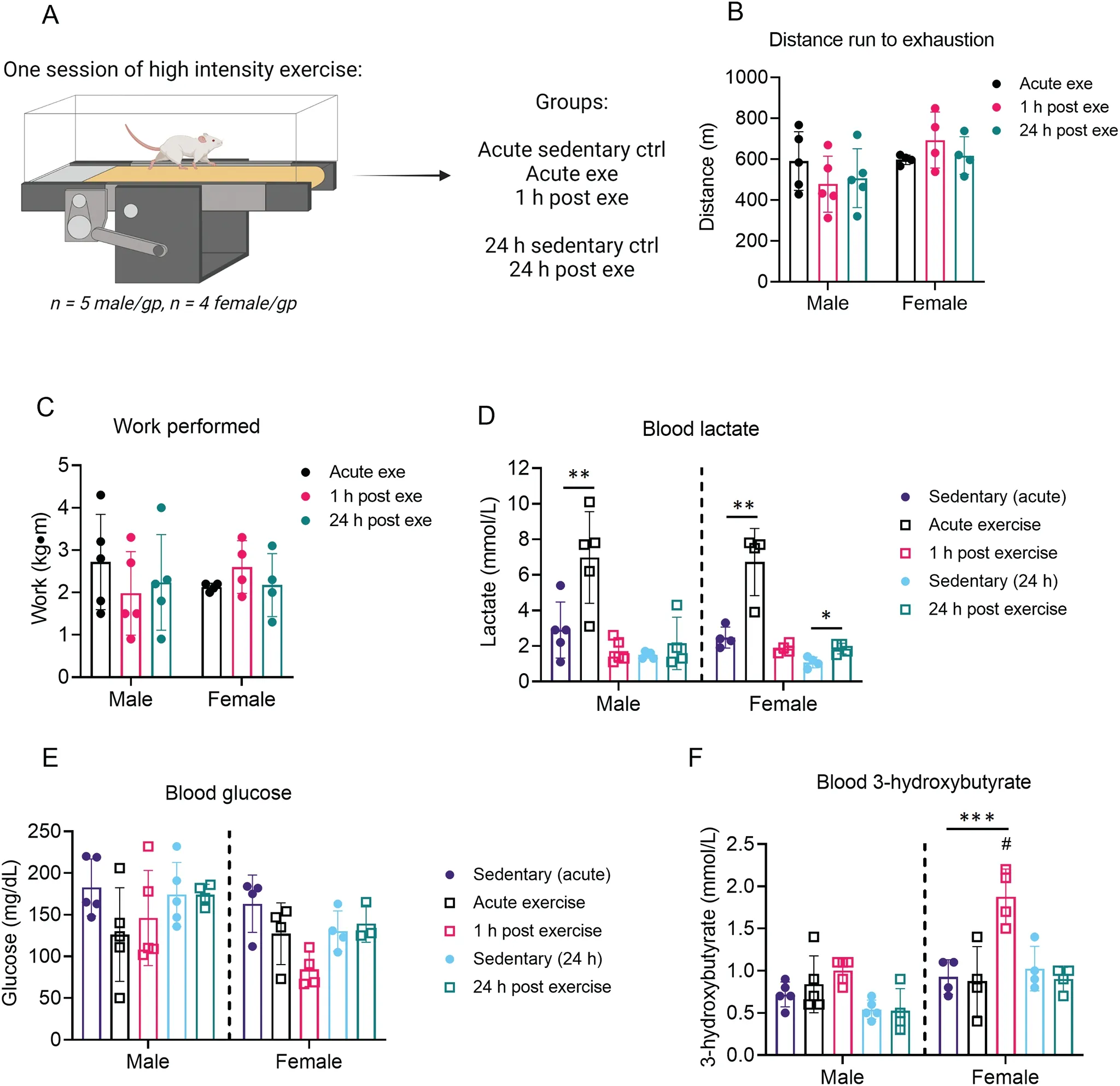
Fig.2. Time-dependent changes in circulating substrates after 1 bout of high-intensity exercise.Mice were subjected to 1 bout of high-intensity exercise(i.e.,exercise capacity test)followed by measurement of circulating substrates immediately,1 h,and 24 h after the exercise bout.(A)Schematic of study design;(B)Distance run to exhaustion; (C) Work performed during the exercise session; and measurements of circulating (D) lactate, (E) glucose, and (F) 3-hydroxybutrate.n=4-5 mice per group.**p <0.01,***p <0.001,two-way ANOVA with Bonferroni’s post hoc test(B,C);*p <0.05,**p <0.01,***p <0.001, #p <0.05 female vs.male,two-way ANOVA with Bonferroni’s post hoc test for acute sedentary ctrl,acute exe,and 1 h post exe group;Student’s t test for 24 h sedentary ctrl and 24 h post exe group(D-F).ANOVA=analysis of variance;ctrl=control;exe=exercise;gp=group.
3.3. Acute effects of exercise on the female cardiac metabolome
Because of these sex-dependent differences in baseline cardiac metabolite abundance, we separately analyzed metabolomics data from hearts of exercised male and female mice. In female hearts, 30 metabolites changed immediately after exercise or upon 1 h of recovery (FDR< 0.10) from an acute bout of high-intensity exercise.Heatmap and relative abundance analyses suggest exercise-induced increases in a cluster of metabolites, including corticosterone, N-acetylleucine, indolelactate,allantoin, and amino acids such as tyrosine and tryptophan,all of which return to near sedentary levels 1 h following exercise (Fig. 5A, 5B). Abundances of alanine and serine decreased immediately following exercise and were even lower in the 1 h recovery period. Interestingly, the majority of metabolite changes occurred 1 h following highintensity exercise, with prominent increases in 3-hydroxybutyrate and isoleucine as well as lipid pathway metabolites, such as the major intermediate of phospholipid biosynthesis, CDP-choline. Several long chain fatty acids and glycerolipid species were also increased 1 h after exercise in the female hearts (Fig. 5A). PLS-DA and VIP score plots further support a prominent response in the female cardiac metabolome 1 h after exercise, with 3-hydroxybutyrate contributing most to group separation (Fig. 5C, 5D).Pathway impact analysis suggest that amino acid metabolism and biosynthesis are acute responses to exercise(Fig. 5E), which could highlight the importance of amino acid mobilization or utilization in the female heart during exercise.
3.4. Acute effects of exercise on the male cardiac metabolome
In hearts from male mice subjected to an acute bout of highintensity exercise, we observed few significant changes in metabolite abundances following exercise (Fig. 6A); corticosterone was the only significantly changed metabolite (FDR<0.10)following exercise in male hearts.Furthermore,PLS-DA plots suggest modest overlap of group clustering (Fig. 6B)and no indication of substantial divergence. Nevertheless,three metabolites contributed substantially (VIP score >5) to group differences (Fig. 6C), including the bile acid tauroβ-muricholate,the ketone body 3-hydroxybutrate,and the phenylsulfate 4-vinylphenol sulfate. Collectively, these findings suggest that a single bout of high-intensity exercise increases corticosterone levels acutely in the murine heart and that female hearts have more pronounced metabolic responses to exercise compared with male hearts.
3.5. The murine heart metabolome 24 h after an exercise bout
To determine how the cardiac metabolome responds to a longer period of recovery following a bout of exercise,we performed separate analyses on samples collected 24 h after 1 bout of high-intensity exercise.In female hearts,heatmap analysis of the top 50 most significantly changed metabolites indicate distinct clustering of sedentary and exercised mouse hearts;however,only 1,5-anhydroglucitol and 4-chlorobenzoic acid reached an FDR <0.10(Supplementary Fig.1A).Nevertheless, PLS-DA analysis suggested significant separation of groups(Supplementary Fig.1B),with heme,α-hydroxyisovalerate,and hexanoylglycine contributing greatest to group separation(data not shown).
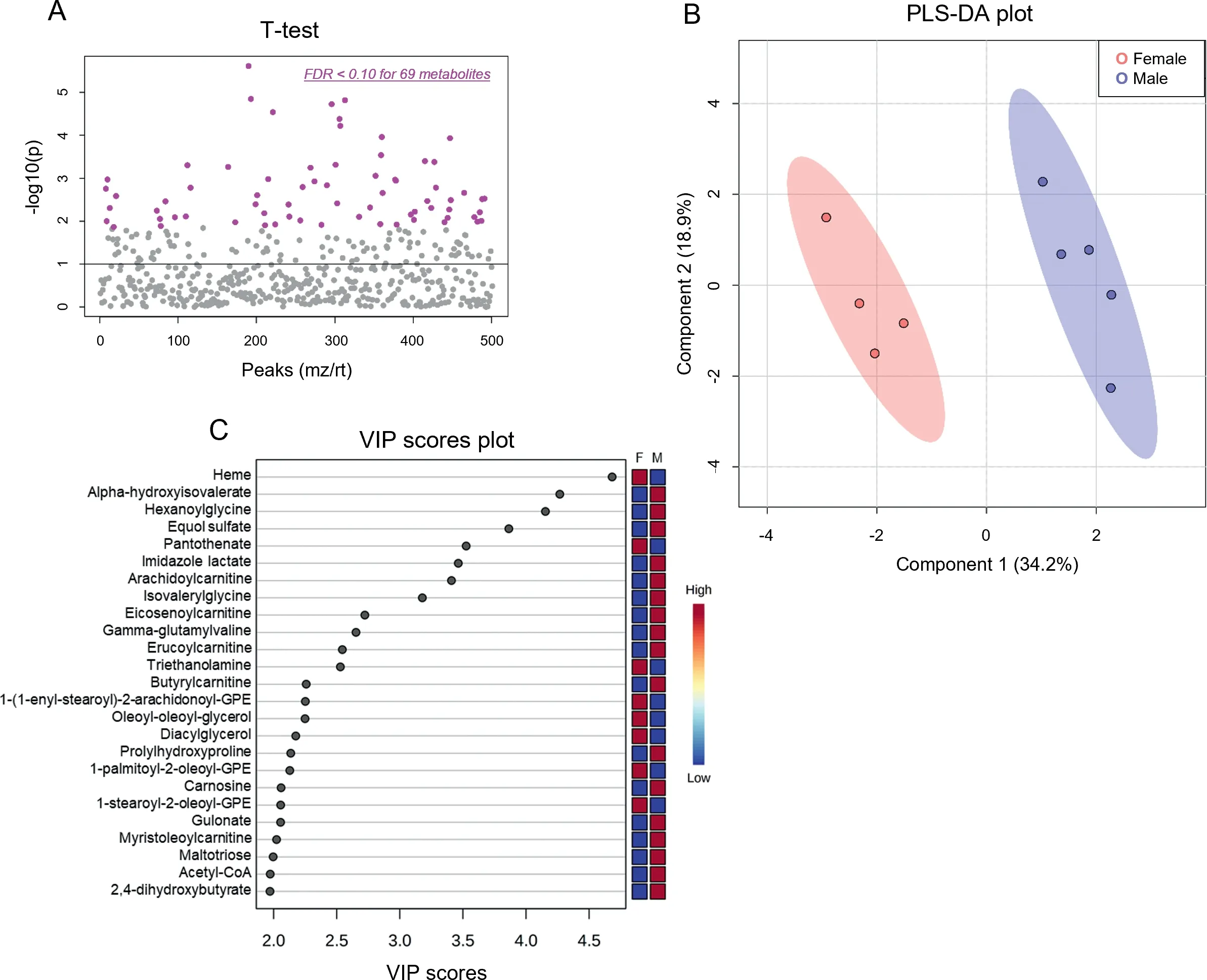
Fig. 3. Biological sex influences baseline cardiac metabolite abundances. Unbiased metabolomics of male and female sedentary mice. (A) Dot plot showing 69 significantly different(FDR <0.10)metabolites between male and female hearts;(B)Partial least-squares discriminant analysis;and(C)Variable importance plot assessing metabolite contribution to the PLS-DA model. n=4-5 mice per group. F=female; FDR=false discovery rate; GPE=glycerophosphoethanolamine;M=male;mz=mass to charge ratio;PLS-DA=partial least-squares discriminant analysis;rt=retention time;VIP=variable importance plot.
Because 1,5-anhydroglucitol is known to be an indicator of glycemic control24,25and could suggest differences in intermediary metabolism, we further examined glucose-derived metabolites in the female hearts. We observed no changes in glucose, glucose-6-phosphate, or fructose-1,6-bisphosphate in hearts 24 h following exercise;however,compared with hearts of sedentary mice,the abundances of 3-carbon glycolytic intermediates (3-phosphoglycerate, phosphoenolpyruvate, and pyruvate) appeared lower in the hearts from exercised mice(Supplementary Fig. 1C). The Krebs cycle metabolites citrate and aconitate were 35%-50% higher in exercised female hearts; however, α-ketoglutarate, fumarate, and malate were 10%-25%lower than sedentary controls(Supplementary Fig.1D).Because exercise has been suggested to be a robust regulator of the antioxidant response,26we also examined the effect of exercise on antioxidants (e.g., reduced glutathione, α-tocopherol, anserine, carnosine) and species derived from oxidative stress (e.g., oxidized glutathione, 4-hydroxynonenal); although no significant differences were observed, the levels of oxidized glutathione and 4-hydroxynonenal appeared lower in female hearts 24 h after exercise(Supplementary Fig.1E).
In male hearts,there were no significantly changed metabolites 24 h after the exercise bout (Supplementary Fig. 2A);however, PLS-DA analysis suggested modest separation between groups (Supplementary Fig. 2B). In general, glycolytic intermediates seemed higher 24 h after exercise(Supplementary Fig.2C),and few changes in Krebs cycle metabolites were observed, with only slight reductions in α-ketoglutarate and succinate (15% and 52% reduction, respectively) compared with sedentary hearts(Supplementary Fig.2D).Distinct patterns in antioxidant and oxidation products were observed in male hearts as compared with female hearts 24 h after an exercise bout, with glutathione, anserine, and carnosine appearing lower(Supplementary Fig.2E male,Supplementary Fig. 1E female). Although these sex-dependent patterns warrant further investigation,these findings indicate that exerciseinduced changes in the murine cardiac metabolome generally return to resting levels 24 h after an exercise bout.
3.6. Influence of a single bout of exercise on cardiac mitochondrial function
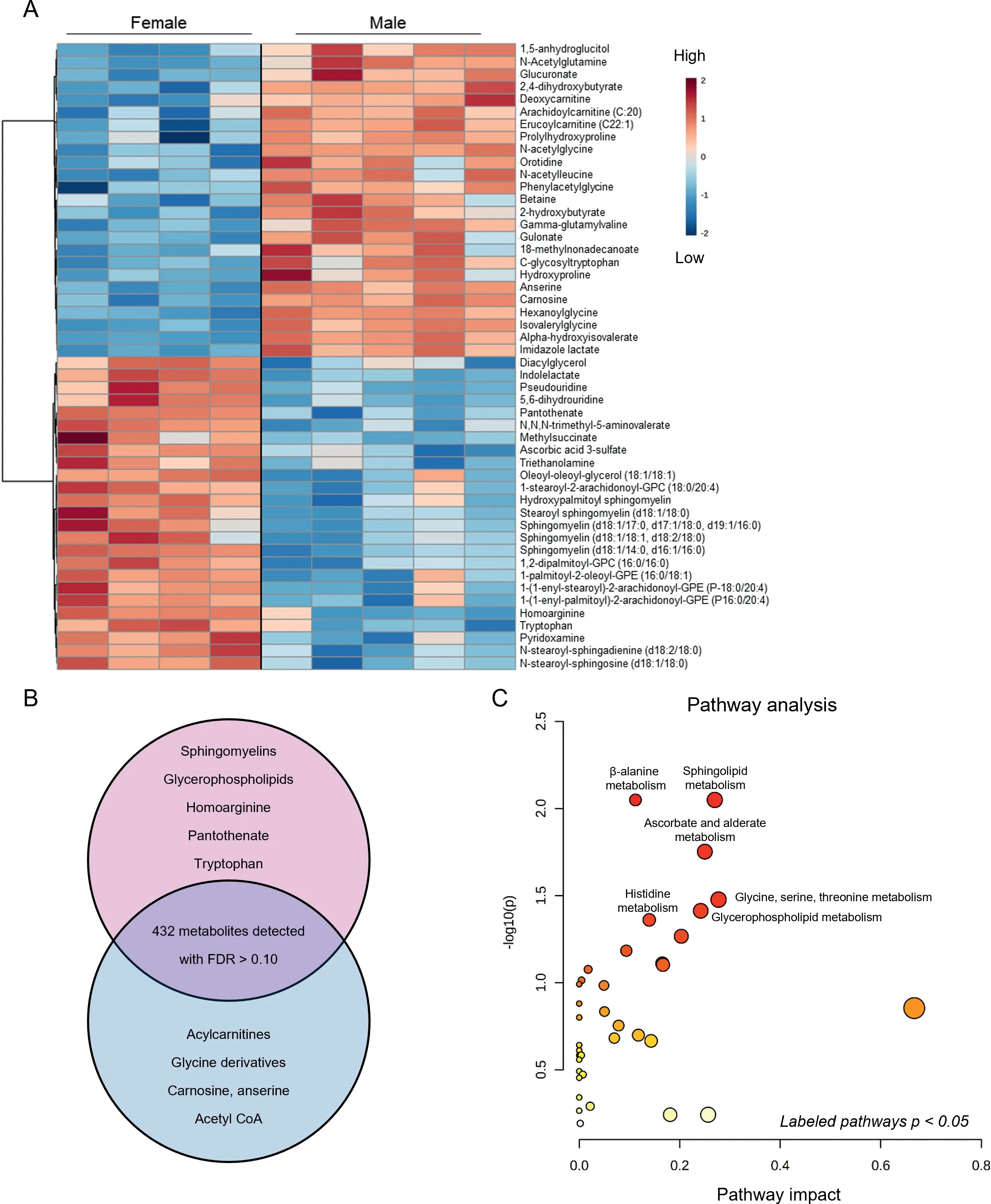
Fig.4. Major metabolomic differences between male and female hearts.Metabolomic analyses highlighting the influence of sex on basal metabolite abundances in the hearts of sedentary mice.(A)Heatmap of top 50 significantly different metabolites;(B)Venn diagram displaying major metabolites and metabolite classes that differ based on sex;and(C)Pathway impact analysis.n=4-5 mice per group.FDR=false discovery rate;GPC=glycerophosphocholine.
To determine whether exercise acutely influences mitochondrial function in the heart,we subjected a second group of mice to an exhaustive bout of high-intensity exercise and then immediately isolated cardiac mitochondria and performed respirometry studies.Female and male mice ran similar distances to exhaustion(Fig. 7A). Cardiac state 3 respiration measured using glutamate,pyruvate,succinate,or octanoylcarnitine as substrates was similar between exercised and sedentary mouse hearts(Fig.7B).Furthermore,there were no differences in state 4 respiration(Fig.7C)or in respiratory control ratios(Fig.7D)between cardiac mitochondria from exercised and sedentary mice.Nevertheless,succinatesupported state 3 and state 4 respiration were generally higher in male cardiac mitochondria compared with female cardiac mitochondria.
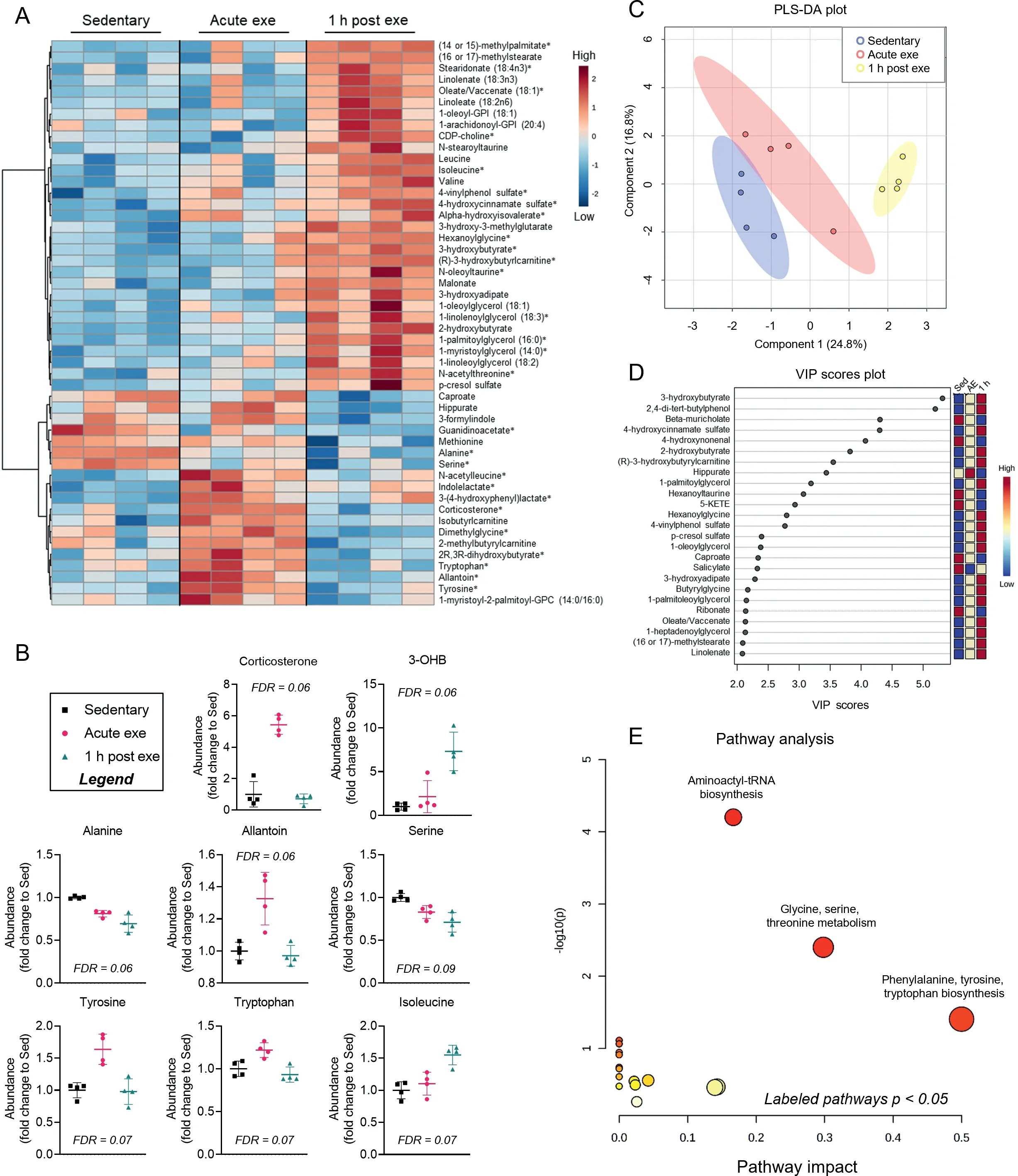
Fig.5. Changes in cardiac metabolite abundances following 1 session of exercise in female murine hearts.Female mice were subjected to 1 bout of high-intensity exercise and hearts were freeze-clamped immediately or 1 h after the exercise bout for unbiased metabolomic analyses.(A)Heatmap of the top 50 most changed metabolites in hearts from exercised versus sedentary female mice;bolded metabolites indicate FDR <0.10 following one-way ANOVA;(B)Graphs of individual metabolites that showed the most prominent changes caused by exercise;(C)PLS-DA plot;(D)corresponding VIP plot;and(E)pathway impact analysis derived from most significantly changed metabolites.n=4 female mice per group.3-OHB=3-hydroxybutyrate;ANOVA=analysis of variance;Exe=exercise;FDR=false discovery rate;GPC=glycerophosphocholine;GPI=glycosylphosphatidylinositol;PLS-DA=partial least-squares discriminant analysis;VIP=variable importance plot.
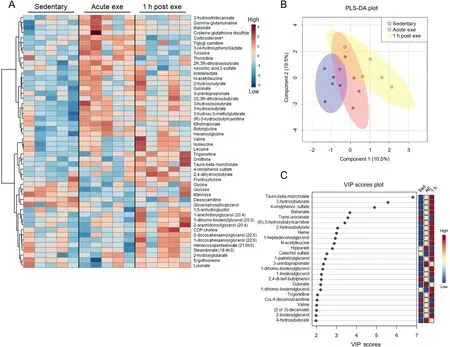
Fig.6. Male hearts demonstrate few significantly changed metabolites after a bout of high-intensity exercise.Male mice were subjected to one bout of high-intensity exercise and hearts were freeze-clamped immediately or 1 h after the exercise bout for unbiased metabolomic analyses. (A) Heatmap of the top 50 most changed metabolites in hearts from exercised and sedentary male mice;bolded metabolites indicate FDR <0.10 following one-way ANOVA;(B)PLS-DA plot,and(C)corresponding VIP plot.n=5 male mice per group.ANOVA=analysis of variance;CDP=Cytidine diphosphate;exe=exercise; FDR=False discovery rate;PLS-DA=partial least-squares discriminant analysis;VIP=variable importance plot.
Because ADP sensitivity could influence mitochondrial function,27we next titrated ADP in cardiac mitochondria isolated immediately after the exercise bout and measured the respiratory response when pyruvate was supplied as a substrate. We found that exercise did not influence ADP sensitivity; however, ADP sensitivity was found to be generally higher in female hearts(Fig.7E).
4. Discussion
Guided by the premise that changes in metabolism influence structural and functional adaptations of the heart to exercise, we examined cardiac metabolite profiles and mitochondrial respiration in mice after a session of treadmill exercise. We found that exercise intensity differentially affects levels of circulating lactate and 3-hydroxybutyrate in both male and female mice, and that high-intensity exercise alters circulating 3-hydroxybutrate only in female mice 1 h following exercise. Untargeted metabolomics also revealed marked differences in the female and male cardiac metabolomes and distinguished the female heart as having more marked changes in cardiac metabolite profile after exercise compared with male hearts. Notable sex-dependent differences in the basal cardiac metabolome include higher levels of heme, pantothenate, homoarginine, and several sphingolipid and phospholipid species but lower levels of several metabolites, including acetyl CoA, glucuronate, carnosine, anserine, hydroxyproline, prohydroxyproline, and carnitinylated and glycinated species, in female hearts compared with male hearts. Female mice also demonstrated more extensive exercise-induced changes in the cardiac metabolome,characterized by significant increases in tyrosine, tryptophan, branched-chain amino acids, and 3-hydroxybutyrate and decreases in serine and alanine. Although a single high-intensity session of exercise did not affect cardiac mitochondrial respiration in either sex, female cardiac mitochondria showed basally higher sensitivity to ADP.Together, these findings in mice suggest that the cardiac metabolite profile is distinct in male and female mice and that female mice demonstrate more robust changes in cardiac metabolite abundance following a session of highintensity exercise.
We chose the FVB/NJ mouse strain and treadmill exercise as our model system for several reasons.Compared with other mouse strains, FVB/NJ mice are elite treadmill runners and show clear cardiac adaptation to regular treadmill training regimens.8,19,28-30Also, this strain is devoid of the known nicotinamide nucleotide transhydrogenase (Nnt) mutations present in the commonly used C57BL/6J strain,31-34which could influence metabolic responses to exercise.The treadmill exercise modality was chosen because it allows control of work and intensity and because compliance with the treadmill protocol is not an issue with the FVB/NJ strain.19,35Using this protocol, we observed apparent sex-based differences in work performed at low and moderate intensities;however,there was no difference in work performed during the high-intensity exercise capacity test. Thus, because we chose to study metabolic changes in the heart after a high-intensity bout of exercise, which showed no difference in work between male and female mice, differences in work do not seem to underlie the disparate responses in the cardiac metabolomes to exercise in male and female mice. Nevertheless, we found that different intensities of exercise training elicit differential changes in circulating lactate and ketone bodies, which are known to influence cardiac metabolism.4,7The finding that the highintensity protocol increased circulating 3-hydroxybutyrate only in female mice and that this corresponded with lower blood glucose levels suggests different systemic glucose handling compared with male mice. This could be due to sexdependent differences in hormones,which could affect metabolism and are known to influence critical processes such as liver gluconeogenesis and glycogenolysis.36Regardless, the higher blood levels of 3-hydroxybutyrate following exercise in female mice could explain the increase in intracardiac 3-hydroxybutryate, especially since recent studies confirmed a mass-action relationship between circulating 3-hydroxybutyrate and its levels and utilization in tissue.37These differences in ketone body metabolism could be important given recent advances in our understanding of the significance of ketone bodies to cardiac biology.38-40
One of the most surprising findings of our study was the marked differences in cardiac metabolite profiles between male and female mice, even in the absence of exercise stress.Unfortunately, few comparisons between female and male sexes in cardiac biology and metabolism are available in preclinical research literature; however, numerous recent studies address sex-based differences in cardiac structure, function,metabolism, and responses to stress. For example, gonadal hormones influence cardiac cellularity and modify the levels of mesenchymal cell and leukocyte populations.41,42Thus, it remains possible that hormonal differences between male and female mice affect the levels of resident and circulating cells in the heart, which could influence the cardiac metabolome at the whole-organ level. Furthermore, estrogen hormones such as 17β-estradiol influence mitochondrial dynamics43and could influence the steady state levels of metabolites in cardiomyocytes. As the field progresses, we anticipate that our understanding of hormone-mediated, sex-based differences in cardiac metabolism will be further illuminated.
Consistent with previous studies showing that lipid metabolism between sexes may be different,44,45we found higher levels of sphingomyelin and glycerophospholipid species in female hearts. Sphingomyelins are a common sphingolipid in mammalian tissues and have important structural and signaling roles.46The higher levels of several sphingomyelins and sphingomyelin-related species (as well as glycerophospholipids) in the female heart could imply higher synthesis or transport, or lower turnover, of these species than in the male heart; however,the significance of these differences to sex-dependent differences in cardiac biology remain unclear and require further investigation. Similarly, tryptophan, pantothenate, pyridoxamine,and homoarginine were higher basally in female hearts compared to male hearts.Tryptophan is a precursor for several intermediates and end products,including kynurenine,nicotinamide, NAD+, and acetyl CoA. Uncontrolled catabolism of tryptophan has been demonstrated in conditions of cardiovascular disease,47and it remains possible that female mice have generally lower catabolism of tryptophan, which could in part underlie their endogenous cardioprotected phenotype.48Pantothenate and pyridoxamine, which were also higher in female hearts,are B vitamins(vitamins B5 and B6,respectively) that could affect energy metabolism. Pantothenate is required for CoA biosynthesis, and deletion of pantothenate kinase has been shown to exacerbate ventricular dysfunction in pressure overload and to cause marked metabolic changes:49;thus,it is possible that the higher basal levels of pantothenate in female hearts could play a role in maintaining CoA levels and bioenergetics.Higher levels of pyridoxamine in female hearts could also underpin the more resilient nature of the female heart.50because pyridoxamine has been shown to protect against cardiac dysfunction caused by aging51or myocardial infarction.52Also markedly higher in female hearts was homoarginine.Homoarginine is a non-proteinogenic, vasoactive amino acid that is a candidate cardiovascular risk factor,with low circulating levels associated with cardiovascular disease.53,54It also has protective actions in the context of myocardial responses to ischemic injury,55,56which may in part underlie the known cardioprotected female phenotype.50
Several metabolites critical in intermediary metabolism were lower in non-exercised female hearts than in corresponding male hearts. The generally lower levels of acetyl CoA,anserine,and carnosine suggest potential differences in central carbon metabolism. Acetyl CoA participates in carbohydrate,protein, and lipid metabolism, with a primary function being the delivery of acetyl groups to the tricarboxylic acid cycle.Although these data could suggest a limitation in energy metabolism in the female heart,the fact that none of the tricarboxylic acid cycle intermediates were significantly lower suggests that the lower levels of acetyl CoA are likely not related to energy deficits. Metabolomic data further indicated sexdependent differences in glucose metabolism. Maltotriose, an intermediate in glycogen metabolism, was lower in female hearts,as were carnosine and anserine,which are small dipeptides that influence glycolytic rate by buffering protons and can scavenge reactive electrophiles generated from oxidative stress.57Moreover,glucose-derived intermediates in the ascorbic acid synthesis pathway, i.e., glucuronate and gulonate,were lower in female hearts. Further understanding of basal sex-dependent differences in cardiac glucose metabolism appears to require carefully designed glucose tracer studies.
After exercise, corticosterone was the only metabolite that was consistently higher in both male and female hearts.While not measured in this study,blood levels of corticosterone have been shown to increase immediately after exercise in several species,58-64which could underlie its higher abundance in the heart after exercise. Corticosterone is required in fetal cardiac development,65and corticosterone signaling is essential for maintaining cardiac function in adult mice.66Although it remains unclear what the role of corticosterone may be in the post-exercised heart, it has been suggested to influence exercise-induced cardiac hypertrophy.67Nevertheless, the extent to which it does so remains unclear. Given that it influences inflammatory responses,68known to be important for tissue repair,69it could play a role in tissue remodeling by altering immune cell responses. Furthermore, glucocorticoids such as corticosterone can influence the expression of genes such as cyclooxygenase-1 and -2 in cardiomyocytes70-72in part through its interaction with C/EBPβ,73which is known to be involved in exercise-induced cardiac remodeling.74Moreover,glucocorticoid receptor activation in myocytes regulates the expression of numerous genes that contribute to cardiac hypertrophy.75Therefore,it is plausible that acute,exercise-induced increases in corticosterone could influence gene programs important for cardiac adapttion to exercise.
ADP sensitivity could be important for metabolic responses of tissues to exercise, as deletion of the adenine nucleotide transporter in mice promotes severe exercise intolerance.76The finding that female cardiac mitochondria have intrinsically higher ADP sensitivity compared with male cardiac mitochondria could help explain why exercise had a more robust effect on the female cardiac metabolome. Because mitochondrial activity is inextricably linked with cellular redox state and cytosolic metabolite pools, the higher sensitivity of female mitochondria to ADP could suggest a more sensitive and responsive metabolic network. This would be consistent with the more pronounced changes in the cardiac metabolome of female mice following exercise.Immediately after high-intensity exercise, female hearts had higher levels of tryptophan,tyrosine, and allantoin but lower levels of alanine and serine.One hour after the exercise bout, several species (in addition to 3-hydroxybutryate) were elevated, including long chain amino acids, branched-chain amino acids, CDP-choline, and glycerophospholipid species, This could suggest alterations in catabolic pathways such as fatty acid and BCAA oxidation as well as phospholipid metabolism. Understanding the significance of elevations in these metabolites to cardiac responses to exercise requires further inquiry.
There are some limitations to this study that deserve mention. First, the untargeted metabolomics approach used in this study to measure cardiac metabolite abundances does not deliver confident assessments in flux.21Thus, additional studies using stable isotope tracers in vivo77could provide a more refined view of how cardiac metabolism changes with exercise.Second,although we showed that exercise intensity influences circulating substrate availability, we did not perform metabolomics analyses on hearts of mice exercised at low or moderate intensities; rather, we examined metabolomic changes in the heart after an acute,exhaustive bout of exercise,which is thought to be a stronger stimulus for growth. Future studies will be required to delineate how low- and moderateintensity exercise affects cardiac metabolism. Last, because the focus of this study was on acute changes in cardiac metabolism after exercise, we did not include an exercise-adapted group as a comparison. After a chronic training regimen, it is possible that some of the metabolic changes would differ after a bout of exercise or that the state of adaptation influences sexdependent differences.
5. Conclusion
The findings of this study show intrinsic differences in cardiac metabolite profiles between male and female mice and show that,following exercise,the female cardiac metabolome changes to a greater extent than the male cardiac metabolome.Whether these sex-dependent differences are a result of a different hormonal milieu or are due to intrinsically higher sensitivity to metabolic stress, as suggested by higher ADP sensitivity in female cardiac mitochondria, requires further study. Understanding how these metabolic differences influence exercise-induced cardiac adaptations and contribute to cardiac resilience to insult or injury is an exciting goal for future studies.
Acknowledgments
Supported in part by grants from the NIH (HL154663,HL147844,HL130174,HL078825,GM127607).Additionally,HEC was supported by the Jewish Heritage Fund for Excellence. Figures in Fig. 1A and 2A were created at BioRender.com.
Authors’contributions
KF participated in financial support, design and execution of experiments, data analysis, and manuscript writing; HEC participated in design and execution of experiments, data analysis, and manuscript writing; SPJ and BGH participated in financial support, design of experiments, data analysis, and manuscript writing. All authors have read and approved the final version of the manuscript, and agree with the order of presentation of the authors.
Competing interests
All authors declare that they have no competing interests.
Supplementary materials
Supplementary material associated with this article can be found in the online version at doi:10.1016/j.jshs.2022.06.001.
杂志排行

Journal of Sport and Health Science的其它文章
- The Journal of Sport and Health Science:Commemorating a decade of publishing milestones and impact
- Prevalence of meeting 24-Hour Movement Guidelines from pre-school to adolescence:A systematic review and meta-analysis including 387,437 participants and 23 countries
- Which psychosocial factors are associated with return to sport following concussion?A systematic review
- Do people with low back pain walk differently?A systematic review and meta-analysis
- Lymphangiogenesis contributes to exercise-induced physiological cardiac growth
- Exercise promotes angiogenesis by enhancing endothelial cell fatty acid utilization via liver-derived extracellular vesicle miR-122-5p