三种微藻的生物量与其细胞氮磷碳的相互关系
2022-07-27曹煜成孙志伟徐煜胡晓娟苏浩昌徐武杰文国樑鲁敏余招龙
曹煜成, 孙志伟, 徐煜, 胡晓娟,2, 苏浩昌,2, 徐武杰,2, 文国樑,*, 鲁敏,余招龙
三种微藻的生物量与其细胞氮磷碳的相互关系
曹煜成1,2,3, 孙志伟1, 徐煜1,2,3, 胡晓娟1,2, 苏浩昌1,2, 徐武杰1,2, 文国樑1,*, 鲁敏1,余招龙3,4
1. 中国水产科学研究院南海水产研究所/农业农村部南海渔业资源开发利用重点实验室/广东省渔业生态环境重点实验室, 广州 510300 2. 中国水产科学研究院南海水产研究所深圳试验基地, 广东 深圳 518121 3. 岭南现代农业科学与技术广东省实验室茂名分中心, 广东 茂名 525400 4. 广东冠利达海洋生物有限责任公司, 广东 茂名 525400
为明确钝顶螺旋藻 () 、绿色颤藻 ()、牟氏角毛藻 ()等三种池塘水体环境中常见优势微藻的细胞数量与其所含总氮(total nitrogen, TN)、总磷(total phosphorus, TP)和总碳(total carbon, TC)质量之间的数量关系, 对以上微藻分别进行纯培养, 在明确微藻细胞数量的条件下, 将藻细胞冷冻干燥制备为冻干粉, 准确测定其干重(cell dry weight, CDW), 以及所含TN、TP和TC的质量, 并就CDW与其所含TN、TP、TC质量间的数量关系, 以及细胞数量与CDW的相互关系等建立数学方程。以上三种微藻的CDW与其所含TN、TP、TC的质量间均存在显著的线性正相关关系(<0.05), 其中在与TN、TP关系的数学回归方程中线性拟合程度优于TC; 同时, 三种微藻的细胞数量与CDW之间同样呈显著的线性正相关关系(<0.05)。其中钝顶螺旋藻细胞中的TN、TC、TP含量占其CDW的比值分别为29.3%、13.6%和4.4%, 均远高于绿色颤藻和牟氏角毛藻。
钝顶螺旋藻; 牟氏角毛藻; 绿色颤藻; 总氮; 总磷; 总碳
0 前言
微藻在水生态系统中扮演生产者的角色, 它对促进池塘水体环境中的物质循环, 通过光合作用为水体提供持续的溶解氧供给, 维系生态系统的健康运转等具有极其重要的意义。有研究显示, 颤藻、角毛藻、螺旋藻等多为水产养殖池塘常见的微藻优势种群[1–2], 对水体微藻群落的组成及其生态功能具有重要影响。有学者指出在水体中培育以有益硅藻或绿藻为优势的浮游微藻群落, 不仅有利于促进养殖对虾的健康生长和获得良好的生产效益, 还可有效降低水体中的氮磷营养盐的含量, 避免氨氮、亚硝酸盐等有毒有害物质对养殖动物的胁迫影响[3]。在水环境净化工程技术领域, 螺旋藻和小球藻等有益微藻也常被用于水质的净化处理, 均取得了良好的效果, 然而此前大多数的研究主要聚焦于微藻对水体中氮磷营养盐的净化效果分析[4], 而关于微藻对水中碳元素的作用效应研究则相对较少[5–6], 并且在评估微藻的氮磷净化效果时, 大多数学者主要是通过测定水体中特定化学形式的氮磷营养盐含量的变化量来评定其净化效率[7–8], 而从微藻细胞组成成分的角度对其吸收氮、磷、碳的定量分析研究也相对少见。该研究选择了三种池塘水体环境中常见的微藻优势种群, 其中包括蓝藻门(Cyanophyta)的钝顶螺旋藻()SP1、绿色颤藻 () OC1, 硅藻门(Bacillariophyta)的牟氏角毛藻()CM1等, 收集它们的纯培养藻液, 测定微藻细胞数量, 并将藻细胞冷冻干燥制备为冻干粉, 准确测定其干重(cell dry weight, CDW), 以及所含总氮(total nitrogen, TN)、总磷(total phosphorus, TP)和总碳(total organic carbon, TC)的质量。就藻细胞CDW值与其所含TN、TP、TC质量间的数量关系, 以及细胞数量与CDW值的相互关系等建立数学方程, 进而说明随着藻细胞的增殖它们可从外界环境中吸收碳氮磷营养元素的绝对质量关系。通过构建“氮磷碳质量——藻细胞干重——活藻细胞数量”的关系方程, 还可为后续进一步研究测算将以上3种微藻规模化培养用以吸收利用环境碳氮磷营养的效果提供基础参考。
1 材料和方法
1.1 藻种与培养基
钝顶螺旋藻SP1、绿色颤藻OC1、牟氏角毛藻CM1均分离自对虾养殖池塘水体环境, 由中国水产科学研究院南海水产研究所农业农村部南海渔业资源开发利用重点实验室提供。钝顶螺旋藻SP1参考Zarrouk培养基配方配制[9–11], 绿色颤藻OC1和牟氏角毛藻CM1分别采用BG11培养基[12]和f/2培养基[13]。
1.2 微藻的培养与收集处理
1.2.1 微藻的培养
钝顶螺旋藻SP1、绿色颤藻OC1、牟氏角毛藻CM1分别以106个·mL-1、106个·mL-1、104个·mL-1的初始接种密度将藻种液接至容量为3L的锥形瓶中, 每组4个平行, 培养瓶置于恒温光照培养箱(Mgc800H, 上海一恒)中进行培养, 培养液盐度20, 培养温度(25±1)℃, 光照强度2500—3000 lx, 光暗比12 h : 12 h, 每日摇动藻液培养瓶3次, 避免藻细胞贴壁生长[14]。
1.2.2 藻液的收集与处理
收集的钝顶螺旋藻SP1和绿色颤藻OC1藻液, 取20 mL藻液用于藻细胞密度测定, 检测前利用超声波细胞粉碎机(JY92-IIDN, 宁波新芝)进行处理, 在不影响藻细胞形态完整的条件下将蓝藻细胞团块破碎为大量易于计数的短杆状藻细胞群体, 角毛藻CM1藻液则无需超声波处理。利用光学生物显微镜(BX43, Olympus)和血球计数板对不同藻液中的藻细胞进行准确计数。
根据不同微藻的细胞形态大小及藻液中细胞数量的具体状况, 分别各取钝顶螺旋藻SP1藻液80 mL、40 mL、20 mL和15 mL置于50 mL离心管, 绿色颤藻OC1和牟氏角毛藻CM1的藻液取样量则设为800 mL、400 mL、200 mL和100 mL, 其余操作均与螺旋藻样品相同。利用大容量冷冻离心机(J-26XP, Beckman coulter), 16128 g, 4℃离心10 min, 弃上清收集藻泥沉淀, 并反复3次对藻泥沉淀以无菌生理盐水进行振荡重悬、清洗、离心。最终获得的藻泥沉淀置于-80 ℃冰箱预冷4 h, 用冷冻干燥机(Alpha 1-4, Christ)将藻泥沉淀冻干至粉末状, 再以分析天平(BS224S, Sartorius)对微藻冻干粉样品进行准确称重。
1.3 微藻生物质氮磷碳的检测
分别各称取三种微藻的冻干粉10 mg, 以100 mL的容量瓶和无菌生理盐水进行定容, 制备浓度为0.1 mg·mL-1的微藻干粉标准使用液。各取标准使用液0 mL、1 mL、2 mL、4 mL、6 mL、8 mL, 再以100 mL的容量瓶进行定容, 获得分别含有微藻冻干粉0 mg、0.1 mg、0.2 mg、0.4 mg、0.6 mg、0.8 mg的溶液样品。参照GB/T 12763.4—2007方法测定样品中的总氮和总磷, 参照GB 17378.4—2007方法测定总碳。
1.4 数据分析
采用Microsoft office excel 2016软件对数据结果进行线性拟合分析, 以单因素方差分析法比较各组数据的差异显著性, 显著水平设定为< 0.05。定量分析三种微藻的细胞干重与其TN、TP、TC含量的相关性, 以及不同微藻的细胞数量与其干重的关系, 获得三种微藻的细胞数量、细胞干重与其藻细胞中TN、TP、TC含量之间的数学方程。
2 结果与分析
2.1 三种微藻的细胞干重与其TN含量的关系
检测结果显示钝顶螺旋藻SP1、绿色颤藻OC1、牟氏角毛藻CM1的细胞干重与其TN的含量均呈显著的线性正相关关系(<0.05), 它们的数学方程及其判定系数2值分别为= 0.2845+ 0.0044,2= 0.9919;= 0.1242+ 0.0009,2= 0.9912;= 0.1315- 0.0016,2= 0.9955; 其中表示藻细胞的TN含量,表示藻细胞的干重值(图1和表1)。该结果表明, 随着微藻细胞干重的增加其细胞中的TN含量也呈线性关系随之不断升高。经测定以上三种微藻的TN含量与其细胞干重的比值分别为29.3%、12.6%和12.8%。
2.2 三种微藻的细胞干重与其TP含量的关系
如图2和表1所示, 钝顶螺旋藻SP1、绿色颤藻OC1、牟氏角毛藻CM1的细胞干重与其TP的含量均呈显著的线性正相关关系(<0.05), 它们的数学方程及其判定系数2值分别为= 0.045- 0.0004,2= 0.9988;= 0.00475– 0.00002,2= 0.9941;= 0.0081- 0.0003,2= 0.9911; 其中表示藻细胞的TP含量,表示藻细胞的干重值。这表明随着微藻细胞干重的增加其细胞中的TP含量也呈线性关系随之不断升高。经测定以上三种微藻的TP含量与其细胞干重的比值分别为4.4%、0.5%和0.8%。干重,表示藻细胞数量的log对数值。这表明随着微藻细胞数量的增长其细胞干重也呈线性关系随之不断增加
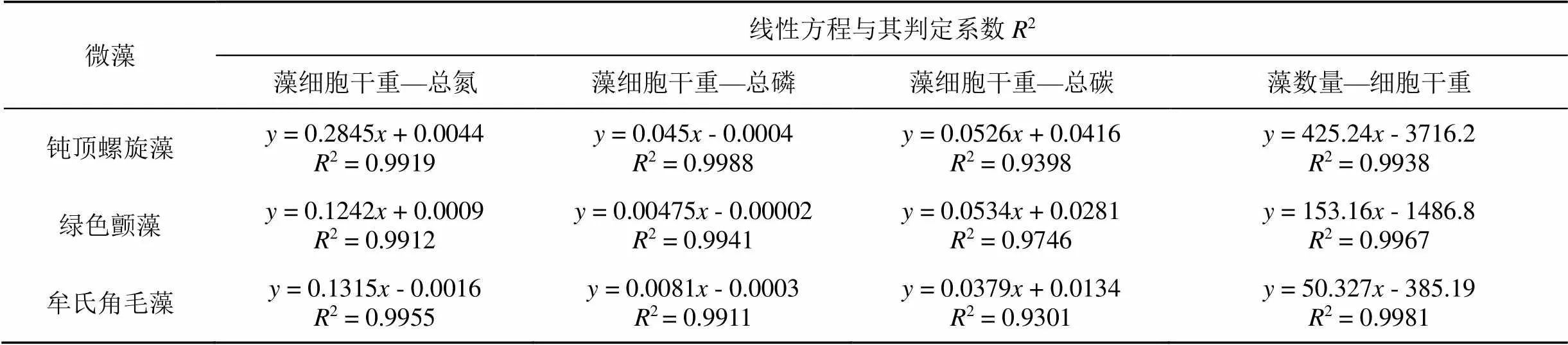
表1 微藻数量、细胞干重和细胞氮磷碳质量的线性关系方程
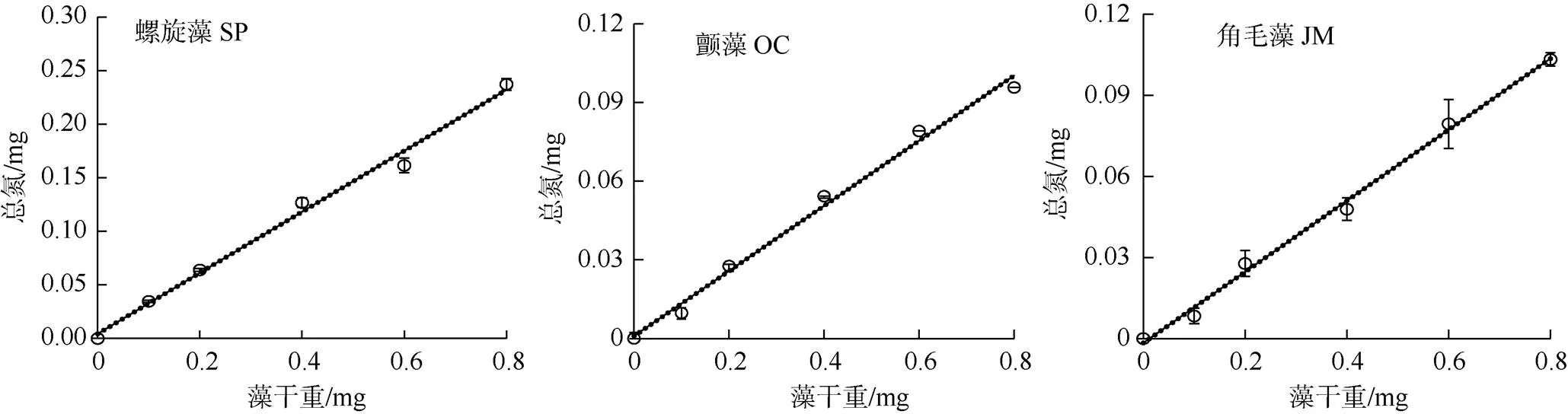
图1 微藻细胞干重与其TN含量的关系
Figure 1 Relationship between dry weight of microalgal cell with their TN content
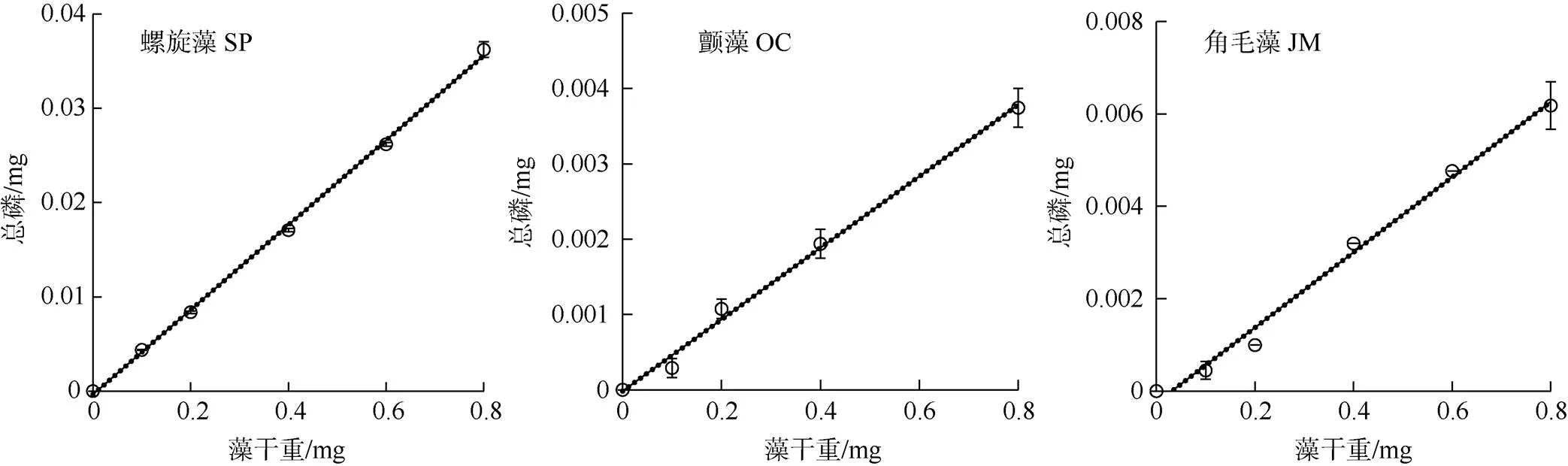
图2 微藻细胞干重与其TP含量的关系
Figure 2 Relationship between dry weight of microalgal cell with their TP content
2.3 三种微藻的细胞干重与其TC含量的关系
钝顶螺旋藻SP1、绿色颤藻OC1、牟氏角毛藻CM1的细胞干重与其TC的含量均呈线性正相关关系(<0.05), 它们的数学方程及其判定系数2值分别为= 0.0526+ 0.0416,2= 0.9398;= 0.0534+ 0.0281,2= 0.9746;= 0.0379+ 0.0134,2= 0.9301; 其中表示藻细胞的TC含量,表示藻细胞的干重值(图3和表1)。可见, 随着微藻细胞干重的增加其细胞中的TC含量也随之不断升高。经测定以上三种微藻的TC含量与其细胞干重的比值分别为13.6%、11.0%和6.5%。
2.4 三种微藻的细胞数量与其干重的关系
如图4和表1所示, 钝顶螺旋藻SP1、绿色颤藻OC1、牟氏角毛藻CM1的细胞数量与其干重之间均呈显著的线性正相关关系(<0.05), 它们的数学方程及其判定系数2值分别为y = 425.24- 3716.2,2= 0.9938;= 153.16- 1486.8,2= 0.9967;= 50.327- 385.19,2= 0.9981; 其中表示藻细胞的。
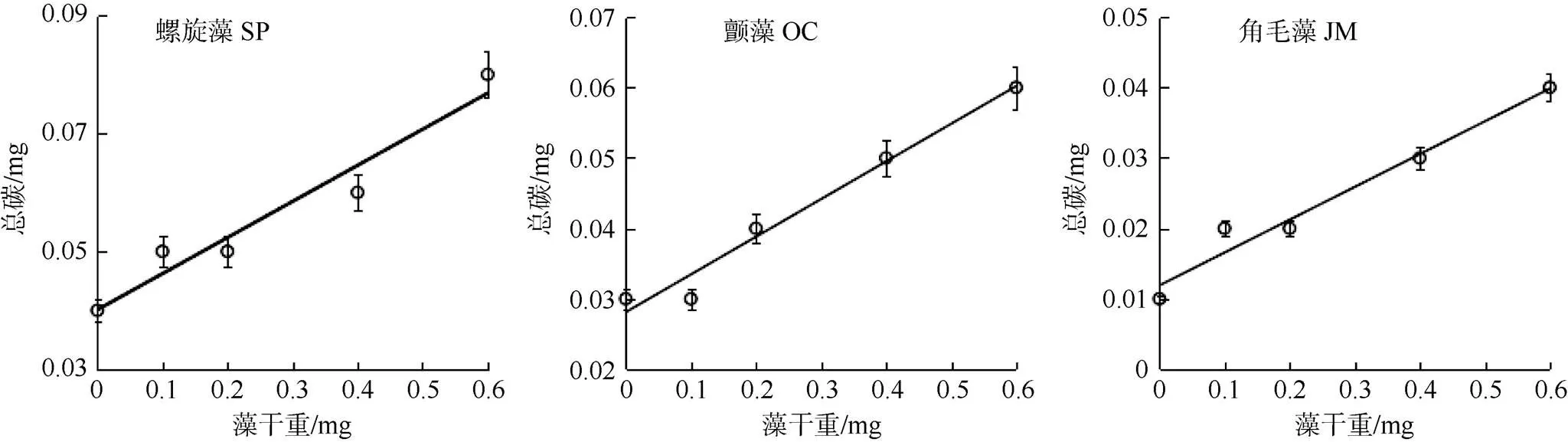
图3 微藻细胞干重与其TC含量的关系
Figure 3 Relationship between dry weight of microalgal cell with their TC content
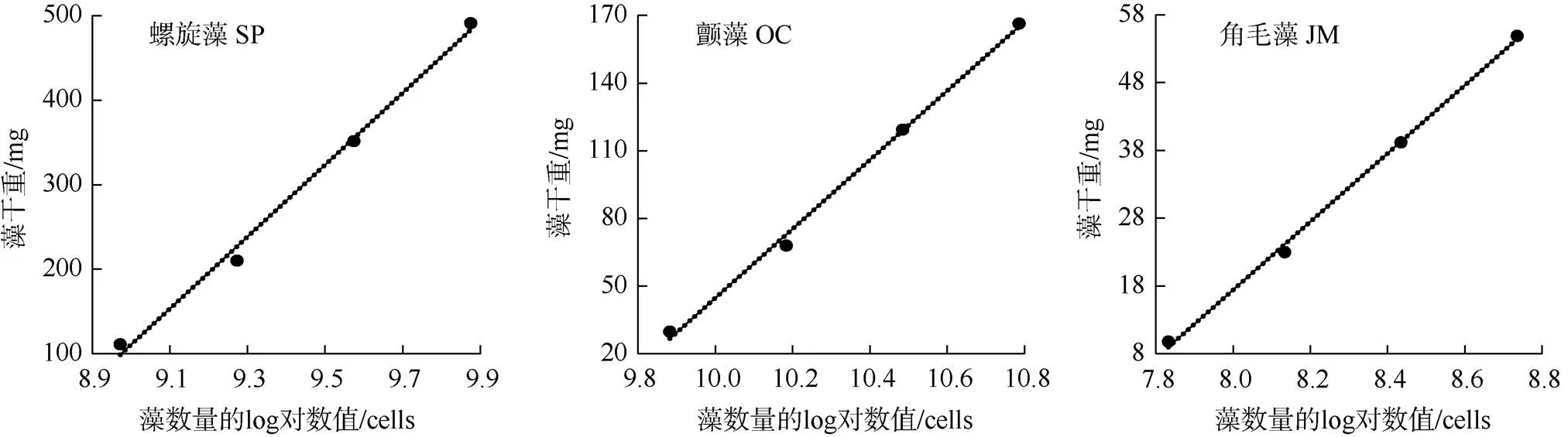
图4 微藻细胞数量与其干重的关系
Figure 4 Relationship between microalgal cell quantities with their dry weight
3 讨论
微藻可高效消减水体中的氮磷营养盐含量, 常被用于各种养殖尾水或工业废水的净化。有学者报道螺旋藻可使水产养殖尾水中的硝酸盐氮(NO3--N)、总无机氮(TIN)和磷酸盐(PO43--P)从初始浓度130.04 mg·L-1、130.85 mg·L-1、10.23 mg·L-1降低至26.53 mg·L-1、70.58 mg·L-1和0.15 mg·L-1, 其最大去除率分别达到79.6%、46.1%和98.5%[14]; 使生猪养殖原废水中氨氮(NH3-N)、TIN和PO43--P从初始浓度164.11 mg·L-1、184.68 mg·L-1、33.85 mg·L-1降低至83.76 mg·L-1、93.89 mg·L-1、19.83 mg·L-1, 去除率为49.0%、49.2%、41.4%[5]; 使啤酒酿制废水中的总氮(TN)、总磷(TP)、化学耗氧量(COD)从初始浓度31.13 mg·L-1、2.08 mg·L-1、700 mg·L-1降低至1.70 mg·L-1、0.80 mg·L-1、102 mg·L-1, 去除率为94.54%、61.54%、85.43%[4]; 使乳品厂废水的TN、TP、COD从初始浓度13.7 mg·L-1、0.376 mg·L-1、110.7 mg·L-1降低至3.2 mg·L-1、0.09 mg·L-1和50.0 mg·L-1, 去除率为76.4%、76.1%和54.8%[6]。可见, 螺旋藻对水体中氮磷净化效果极为明显, 具有广泛的应用前景。
颤藻对尾水的净化效果同样显著, 它可将尾水中的NO3--N、TIN、PO43--P浓度由初始的136.77 mg·L-1、124.32 mg·L-1、11.07 mg·L-1分别降至54.85 mg·L-1、60.12mg·L-1、0.60 mg·L-1[14]。但是绝大部分颤藻会分泌微囊藻毒素, 对养殖生物和人体健康产生严重影响[15-16], 存在较大的安全隐患, 因此即使它作为池塘水体中常见的微藻优势种能对水质起到良好的净化效果, 还是必须对其加以严格防控。角毛藻对水体氮磷的去除效果相对差于螺旋藻和颤藻, 有研究显示牟氏角毛藻可将集约化对虾养殖尾水中的NO3--N、TIN和PO43--P从初始浓度9.35 mg·L-1、10.2mg·L-1和1.18 mg·L-1降至0 mg·L-1、0.44 mg·L-1和0.05 mg·L-1; 其去除率为100%、95.7%和95.8%[14]; 叶志娟等的研究结果显示[7], 用牟氏角毛藻净化海水鱼养殖尾水, 尾水的TN、PO43--P和COD的初始浓度2.212 mg·L-1、0.232 mg·L-1、约35 mg·L-1, 处理后水体的无机氮去除率为90-95%, 磷酸盐为90%, COD含量基本没变化。虽然角毛藻对水体中氮磷营养盐的去除率能达到90%以上, 但它对氮磷去除的绝对数量仍远不及螺旋藻和颤藻。这可能与不同微藻细胞的生理特征、微藻对水环境的适应性[17-19]及其对光照、温度等气候因子的选择性有关[20-21]。
还有不少学者更为关注微藻的营养组分研究与利用。刘如冰等研究认为螺旋藻的蛋白质含量为42.8%—67.4%[6]; 还有学者认为螺旋藻的蛋白质含量为48.1%—69.3%、糖类为5.4%—10.4%、脂肪为4.4%—5.4%、灰分6.2%—7.4%、粗纤维4.2%, 并且螺旋藻所含氨基酸的种类齐全, 总量可达309.94—505.95 mg·g-1[22-24]。所以, 螺旋藻常被用于开发保健食品和养殖动物的饲料蛋白源替代物[25-26]。虽然角毛藻细胞的蛋白质远低于螺旋藻, 但其脂肪含量则远高于后者, 角毛藻的蛋白质含量为11.0%—18.7%, 总脂肪为36.0%—57.1%[27], 所以许多学者将角毛藻作为生物石油的研究对象[28]。总体而言, 对于螺旋藻、颤藻、角毛藻细胞中的氮、磷、碳元素的相关分析则鲜见报道。
由上可见, 目前在微藻与环境营养相关作用方面的研究一方面主要关注于微藻对水体中氮磷营养盐的净化效果分析, 通过测定水体中NO3--N、NO2--N、NH3-N、TIN和PO43--P等特定化学形式的氮磷营养盐含量的变化量来评定其净化效率; 另一方面则聚焦于通过不同种类营养因子或培养条件的优化控制提高微藻细胞中蛋白质、总脂肪、氨基酸、糖类等营养物质的转化与积累效率。然而关于微藻细胞中氮、磷、碳元素与其细胞数量的相关性研究却少见报道。其实, 水体环境中氮、磷、碳元素的化学存在形式具有多样化特征, 例如NO3--N、NO2--N、NH3-N、TIN、PO43--P、H2PO4-、HPO42-、HCO3-、CO32-等的化学形式在水体中均较为常见, 而且它们之间还会随各种环境因子的变动而发生频繁的相互转化[29]。因此, 在利用微藻吸收环境营养的过程中应基于藻细胞数量的增长状况探讨其所含总氮、总磷、总有机碳的数量变化, 这才能准确评测微藻从环境中究竟获取了多少数量的营养物质用于其绝对生物量的增长, 对水环境的绝对净化效应如何, 从而有效避免水环境中元素化学存在形式变动、营养盐的吸附与沉降、水生生物的呼吸与新陈代谢等众多背景干扰因子的影响。虽然钝顶螺旋藻、绿色颤藻、牟氏角毛藻等3种微藻均可通过光合作用途径, 吸收二氧化碳或碳酸氢根合成有机质, 以供其生长所需。而螺旋藻和颤藻作为蓝藻种类, 有研究表明在自然的开放水域环境中蓝藻能通过固氮效应, 从空气中摄取气态氮进行固氮营养[30]; 微生物学领域则将蓝藻定义为蓝细菌, 认为它们不仅可营光能自养生活还可在一定条件下营异养生活从环境中吸收溶解态有机营养[31]。但无论以上3种微藻通过何种营养方式进行生长, 它们的增殖终归需要从外界环境中摄取碳氮磷营养元素。基于此, 该研究的焦点是探讨在适宜的恒温光照纯培养条件下, 以上3种微藻细胞中所含氮、磷、碳的含量状况, 通过建立细胞干重与其碳、氮、磷含量的线性关系方程, 说明随着藻细胞的增殖它们可从外界环境中吸收碳氮磷营养元素的绝对质量关系。其次, 通过逐步分析构建“藻细胞氮磷碳质量——藻细胞干重——活藻细胞数量”的关系方程, 还可为后续进一步研究测算将以上3种微藻进行规模化培养用以吸收环境中碳氮磷营养的效果提供基础参考。
该研究结果显示, 钝顶螺旋藻、绿色颤藻、牟氏角毛藻的细胞干重CDW值与其所含TN、TP、TC的质量间均存在显著的线性正相关关系(<0.05), 而且细胞数量与CDW之间也同样呈显著的线性正相关关系(<0.05), 由此可用其数学回归方程推算随着以上三种微藻细胞数量的增长, 它们从水体环境所吸收氮、磷、碳的绝对数量。此外, 该研究还显示钝顶螺旋藻细胞中的TN、TC、TP含量占其CDW的比值分别为29.3%、13.6%和4.4%, 均远高于牟氏角毛藻, 这也与上述学者报道螺旋藻的高蛋白质含量特征相吻合。由此推算, 每获得干重1 g的钝顶螺旋藻SP1, 它可从环境中吸收氮元素293 mg, 碳元素136 mg、磷元素44 mg; 换而言之, 若以1000 m3的零换水集约化养殖池系统为例, 在其水体中按103个·mL-1浓度接种钝顶螺旋藻SP1, 培养10天左右当它的藻细胞密度增长至108个·mL-1数量水平时, 可获得干重约11.1kg的螺旋藻, 进而从环境中吸收氮元素3.3 kg、碳元素1.5 kg和磷元素0.5 kg用于微藻细胞的生长增殖; 按对虾养殖周期90天计算, 对所培养的螺旋藻进行实时采收并进行利用[32], 那么一个养殖季即可通过螺旋藻的培养获得干重约99.9 kg的螺旋藻, 并从环境中吸收氮元素29.7 kg、碳元素13.5 kg和磷元素4.5 kg。可见, 选择螺旋藻用于水产养殖水体环境的净化, 可实现水中氮、碳、磷等营养元素的高效吸收, 或将之与有机质降解菌、硝化功能菌等进行科学配伍, 应该能有效解决水环境中微生物对水质净化不充分的难题[33]。
4 结论
钝顶螺旋藻SP1、绿色颤藻OC1、牟氏角毛藻CM1等三种微藻的细胞CDW与其所含TN、TP、TC的质量间均存在显著的线性正相关关系(<0.05), 其中在与TN、TP关系的数学回归方程中线性拟合程度优于TC; 同时, 三种微藻的细胞数量与CDW之间亦呈显著的线性正相关关系(<0.05)。钝顶螺旋藻细胞中的TN、TC、TP含量占其CDW的比值分别为29.3%、13.6%和4.4%, 均远高于绿色颤藻和牟氏角毛藻(<0.05)。
[1] 彭聪聪, 李卓佳, 曹煜成, 等.凡纳滨对虾半集约化养殖池塘浮游微藻优势种变动规律及其对养殖环境的影响[J]. 海洋环境科学, 2011, 30(2): 193–198.
[2] 查广才, 周昌清, 黄建荣, 等.凡纳对虾淡化养殖虾池微型浮游生物群落及多样性[J]. 生态学报, 2004, 24(8): 1752–1759.
[3] 李卓佳, 李奕雯, 曹煜成, 等. 对虾养殖环境中浮游微藻、细菌及水质的关系[J]. 广东海洋大学学报, 2009, 29(4): 95–98.
[4] 王钰舟, 杨雷, 张景来, 等. 利用啤酒废水培养极大螺旋藻[J]. 化工环保, 2014, 34(3):257–261.
[5] CAN S S, DEMIR V, CAN E. Evaluating the dilution of municipal wastewater on biomass increase, lipid production and nutrient removal by the blue-green algae(Geitler)[J]. Fresenius Environmental Bulletin, 2015, 24(3): 904–909.
[6] 刘如冰, 马金才, 马维琦, 等. 乳品厂污水养殖螺旋藻的研究[J]. 海洋通报, 2000, 19(1): 68–72.
[7] 叶志娟, 刘兆普, 王长海. 牟氏角毛藻在海水养殖废水中的生长及其对废水的净化作用[J]. 海洋环境科学, 2006, 25(3): 9–12.
[8] 刘林林, 黄旭雄, 危立坤, 等. 15株微藻对猪场养殖污水中氮磷的净化及其细胞营养分析[J]. 环境科学学报, 2014, 34(8): 1986–1994.
[9] REINEHR C O, COSTA J. Repeated batch cultivation of the microalga[J]. World Journal of Microbiology and Biotechnology, 2006, 22(9): 937–943.
[10] RADMANN E M, REINEHR C O, COSTA J. Optimization of the repeated batch cultivation of microalgain open raceway ponds[J]. Aquaculture, 2007, 265(1-4): 118–126.
[11] ZHANG L L, CHEN L, WANG J F, et al. Attached cultivation for improving the biomass productivity of[J]. Bioresource Technology, 2015, 181(1): 136–142.
[12] RAY S, BAGCHI S N. Nutrients and pH regulate algicide accumulation in cultures of the cyanobacterium[J]. New Phytologist, 2001,149(3): 455–460
[13] LANANAN F, JUSOH A, ALI N, et al. Effect of Conway Medium and f/2 Medium on the growth of six genera of South China Sea marine microalgae[J]. Bioresource Technology, 2013, 141: 75–82.
[14] 鲁敏, 曹煜成, 胡晓娟, 等. 钝顶螺旋藻 SP1()对集约化养殖尾水氮磷的去除效果[J]. 生态科学, 2021, 40(2): 125–132.
[15] 徐煜, 徐武杰, 文国樑, 等. 颤藻浓度和水温对凡纳滨对虾响应颤藻粗提液毒性的影响[J]. 南方水产科学, 2017, 13(1): 26–32.
[16] 王朝晖, 张玉娟, 曹宇, 等. 微囊藻毒素对水环境的影响研究进展[J]. 生态科学, 2006, 25(2): 188–191.
[17] CAMPOS D J, ELIAS J A L, JUAREZ L Á M, et al. Chemical composition, fatty acid profile and molecular changes derived from nitrogen stress in the diatom[J]. Aquaculture Reports, 2020, 16(1): 100281.
[18] LIANG C W, ZHANG Y F, WANG L, et al. Features of metabolic regulation revealed by transcriptomic adaptions driven by long term elevated p CO2in[J]. Phycological Research, 2020, 68(3): 236–248.
[19] 胡晗华, 高坤山. CO2浓度倍增对牟氏角毛藻生长和光合作用的影响[J]. 水生生物学报, 2001, 25(6): 636–639.
[20] MINGGAT E, ROSELI W, TANAKA Y. Nutrient Absorption and Biomass Production by the Marine Diatom: Effects of Temperature, Salinity, Photoperiod, and Light Intensity[J]. Journal of Ecological Engineering, 2021,2 2(1): 231–240.
[21] LI Y F, LI R Q, YI X Y. Effects of light quality on growth rates and pigments of(Bacillariophyceae)[J]. Journal of Oceanology and Limnology, 2020, 38(3): 795–801.
[22] 付云, 庞一扬, 赵谋明, 等. 螺旋藻渣营养成分分析及其发酵产物功能活性评价[J]. 食品研究与开发, 2020, 41(24): 186–192.
[23] 董育红, 封涛, 张振兰, 等. 螺旋藻的营养成分分析[J]. 食品研究与开发, 2003, 24(3): 70–71.
[24] 闫春宇, 王素英, 董世瑞. 22株螺旋藻(节旋藻)氨基酸成分分析及营养评价[J]. 食品与机械, 2015, 31(6): 21–27.
[25] 虞为, 杨育凯, 陈智彬, 等. 饲料中添加螺旋藻对花鲈生长性能、消化酶活性、血液学指标及抗氧化能力的影响[J]. 南方水产科学, 2019, 15(3): 57–67.
[26] ADEL M, YEGANEH S, DADAR M, et al. Effects of dietaryon growth performance, humoral and mucosal immune responses and disease resistance in juvenile great sturgeon (Linnaeus, 1754)[J]. Fish and Shellfish Immunology, 2016, 56: 436–444.
[27] 张娜, 胡文峰, 靳翠丽, 等. 3种处理对纤细角毛藻生长及细胞生化组成的影响[J]. 应用海洋学学报, 2018, 37(2): 248–254.
[28] TOKUSHIMA H, INOUE-KASHINO N, NAKAZATO Y, et al. Advantageous characteristics of the diatomas a sustainable biofuel producer[J]. Biotechnology Biofuels, 2016, 9(1): 235.
[29] 李卓佳, 李奕雯, 曹煜成, 等. 凡纳滨对虾海水高位池养殖水体理化因子变化与营养状况分析[J]. 农业环境科学学报, 2010, 29(10): 2025–2032.
[30] 胡鸿钧, 魏印心. 中国淡水藻类-系统、分类及生态[M]. 北京: 科学出版社. 2006, 920–923.
[31] DAVID R B, RICHARD W C. Bergey’s Manual of Systematic Bacteriology (Second Edition) Volume one[M]. NewYork: Springer, 2005.
[32] YU W, WEN G L, LIN H Z, et al. Effects of dietaryon growth performance, hematological and serum biochemical parameters, hepatic antioxidant status, immune responses and disease resistance of Coral trout(Lacepede, 1802)[J]. Fish and Shellfish Immunology, 2018, 74: 649–655.
[33] 韩天骄, 徐武杰, 徐煜, 等. 停加红糖对凡纳滨对虾生物絮团养殖系统水质和氮收支的影响[J]. 南方水产科学, 2020, 16(6): 81–88.
Relationship between cell quantity of three microalgae with their carbon, nitrogen and phosphorus
CAO Yucheng1,2,3, SUN Zhiwei1, XU Yu1,2,3, HU Xiaojuan1,2, SU Haochang1,2, XU Wujie1,2, WEN Guoliang1*, LU Min1, YU Zhaolong3,4
1. South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences/Key Laboratory of South China Sea Fishery Resources Exploitation & Utilization, Ministry of Agriculture and Rural Affairs/Guangdong Provincial Key Laboratory of Fishery Ecology and Environment, Guangzhou 510300, China 2. Shenzhen Base South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Shenzhen 518121, China 3. Maoming Branch of Guandong Laboratory for Lingnan Modern Agriculture, Maoming525400, China 4. Guangdong Guanlida Marine Biology Co., Ltd., Maoming 525400, China
In order to evaluate the relationship between the cell quantity and the contents of total nitrogen (TN), total phosphorus (TP) and total carbon (TC) in,and, which were dominant microalgal species in aquaculture pond, the microalgae were pure cultured in biological incubator with constant temperature and illumination respectively, and cell quantity were determined continuously. Microalgal cells were collected by centrifugation and freeze-dried to prepare dry powder, the cells dry weight (CDW) and the contents of TN, TPandTCalso were accurately measured. Based on the measured data, the mathematical equations were established on quantitative relationship between CDW and the contents of TN, TP and TC, as well as the relationship between microalgal cell quantity and CDW. There was significant linear positive correlation between CDW of the three microalgae with their contents of TN, TP and TC (< 0.05); the linear fit degree in the mathematical regression equation of TN and TP was better than that of TC; meanwhile, the cell quantity of three microalgae also was linear positive correlation with CDW significantly (< 0.05). The ratio of TN, TC and TP to CDW incells was 29.3%, 13.6% and 4.4% respectively, which were much higher than those ofand.
;;; total nitrogen; total phosphorus; total carbon
10.14108/j.cnki.1008-8873.2022.04.024
X703.1
A
1008-8873(2022)04-204-08
2021-10-28;
2021-12-09
国家重点研发计划(2020YFD0900401); 中国水产科学研究院基本科研业务费专项资金(2020TD54); 财政部和农业农村部: 国家现代农业产业技术体系(CARS-48); 广东省现代农业产业技术体系创新团队建设专项资金(2019KJ149); 中国水产科学研究院南海水产研究所中央级公益性科研院所基本科研业务费专项资金项目(2021SD08)
曹煜成(1979—), 男, 浙江东阳人, 博士, 研究员, 主要从事水产健康养殖和养殖生态环境调控研究, E-mail: cyc_715@163.com
通信作者:文国樑, 男, 研究员, 主要从事水产健康养殖研究, E-mail: guowen66@163.com。
曹煜成, 孙志伟, 徐煜, 等. 三种微藻的生物量与其细胞氮磷碳的相互关系[J]. 生态科学, 2022, 41(4): 204–211.
CAO Yucheng, SUN Zhiwei, XU Yu, et al. Relationship between cell quantity of three microalgae with their carbon, nitrogen and phosphorus[J]. Ecological Science, 2022, 41(4): 204–211.
