不同土地整治类型新增耕地土壤微生物群落特征研究
2022-07-22朱奕豪李青梅刘晓丽李娜宋凤玲陈为峰
朱奕豪,李青梅,刘晓丽,李娜,宋凤玲,陈为峰*
1. 山东农业大学资源与环境学院,山东 泰安 271018;2. 山东省土地发展集团有限公司,山东 济南 250014;3. 山东省国土空间生态修复中心,山东 济南 250014;4. 泰安市国土空间生态修复中心,山东 泰安 271099
土地整治包含土地整理、复垦和开发,一般采用工程、生物等措施对散乱、荒废、退化及配置不合理、未被充分利用的土地实施综合治理与深度开发,是提高土地利用率和产出率、改善农业生产条件和土地生态环境、实现土地资源可持续利用和促进土地生态环境良性循环的一种高效土地管理工具(Stańczuk-Gałwiaczek et al.,2018;王军等,2018;陈坤秋等,2020)。近年来,土地综合整治为国家耕地“占补平衡”战略和粮食安全做出了重要贡献,也成为驱动耕地质量变化的一项重要因素(李肖肖等,2019;赵瑞等,2019)。
土地整治工程多采用挖高填低、表土剥离等方式进行土地平整与地块合并,在改善农业生产条件、优化国土空间布局的同时,这一过程不可避免地会对土体构型和土壤质量产生影响,不仅影响土壤理化性状,还会对土壤微生物群落造成一定干扰(Djanibekov et al.,2018;Legrand et al.,2018),而土壤微生物在维持农田生态系统稳定中发挥着重要作用,它们对外界干扰极为敏感,其中细菌因在土壤微生物数量中占有绝对优势并可通过固氮和有机质分解、转化等方式提高土壤养分含量,从而改善土质以达到作物增产的效果(Wang et al.,2018;林耀奔等,2019)。同时,土壤理化性状变化也会造成土壤微生物群落结构的改变。研究表明,通过国土综合整治及强化后续管控,土壤质量会得到良好的改善(Ertunç et al.,2018;林耀奔等,2019;李肖肖等,2020),但也有报道证实工程配套型土地整治短期内对土壤质量和作物长势存在不利影响(李肖肖等,2019)。
目前,土地整治涉及土壤微生物研究主要集中于工矿区土地复垦(李肖肖等,2020;施昊坤等,2020;马静等,2021;原野等,2021)、新增耕地培肥(林耀奔等,2019;罗友进等,2020)、整治质量评价(林耀奔等,2019;赵瑞等,2019)等方面,而不同来源新增耕地土壤微生物群落特征的影响尚缺乏深入研究。探索不同类型新增耕地微生物种群的组成变化、厘清微生物群落分布的主控因素,对新增耕地后期管护至关重要。为此,本研究选取山东省泰安市34个土地整治项目(工矿复垦、占补平衡)作为研究对象,涉及棕壤、褐土、潮土不同土壤类型,分布在丘陵、山地、平原不同地貌区域,选择泰安区域,基本可作为山东开展工矿复垦和占补平衡项目补充耕地区域的典型代表区,以此作为本研究选择区域的切入点,测定土壤理化性状、三大类微生物数量,并利用高通量测序技术测定土壤细菌16 S rRNA,探索不同类型新增耕地土壤微生物群落组成变化,揭示土壤微生物群落变化的关键影响因子,为新增耕地培肥和养护提供技术支撑。
1 材料与方法
1.1 研究区及样品采集与处理
研究区泰安市位于鲁中山区,地理位置116°20—117°59′E,35°38′—36°28′N,属温带大陆性半湿润季风气候区,雨热同季,具有良好的环境条件和悠久的农业历史,境内拥有多种地貌类型,如山地、丘陵、平原、洼地、湖泊等,丰富的地形便于作物的多样化种植,但因地形变化导致地块不集中、耕地不成片的现状限制了规模化的土地经营与流转。因此,通过土地平整、地块合并、水田改造、改善灌排及生态护坡等措施于一体的土地整治工程可有效缓解土地零散的难题,保障土地集中连片、规模经营,极大提高了机械使用率、土地利用率与土壤产能。
本文选择对目前土地整治项目类型中常涉及大规模土壤重构的工矿复垦项目(GK)和占补平衡项目(ZB)进行研究,这两类新增耕地是目前山东省新增耕地指标的主要类型,是社会投资项目的重点,其新增耕地质量问题,也是社会关注的焦点。两类土地整治项目中的土地整理均涉及到客土回填,工矿复垦项目客土一般在60—80 cm,占补平衡项目客土一般在30 cm。2021年10月对泰安市34个项目(GK项目18个,ZB项目16个)开展调查研究,从项目区附近选取非土地整治形成的农田作为对照(CK),采样项目区分布如图1,每个样地采样时均采用五点法采集0—20 cm的表层土,每个项目区取20个样点,5个样点混匀成一份土样,共计208份,现场筛除碎石、枯落物及其他杂物后放于无菌密封塑料袋中,低温保存运输至实验室。之后土样分为3份,一份土样风干后过筛待测,用于土壤理化性状测定;第2份根据实验需求以鲜土方式保存用于土壤细菌、真菌、放线菌测定;第 3份储存于−80 ℃冰箱用于土壤细菌测序(从工矿复垦、占补平衡及对照耕地各选一个样地取样)。
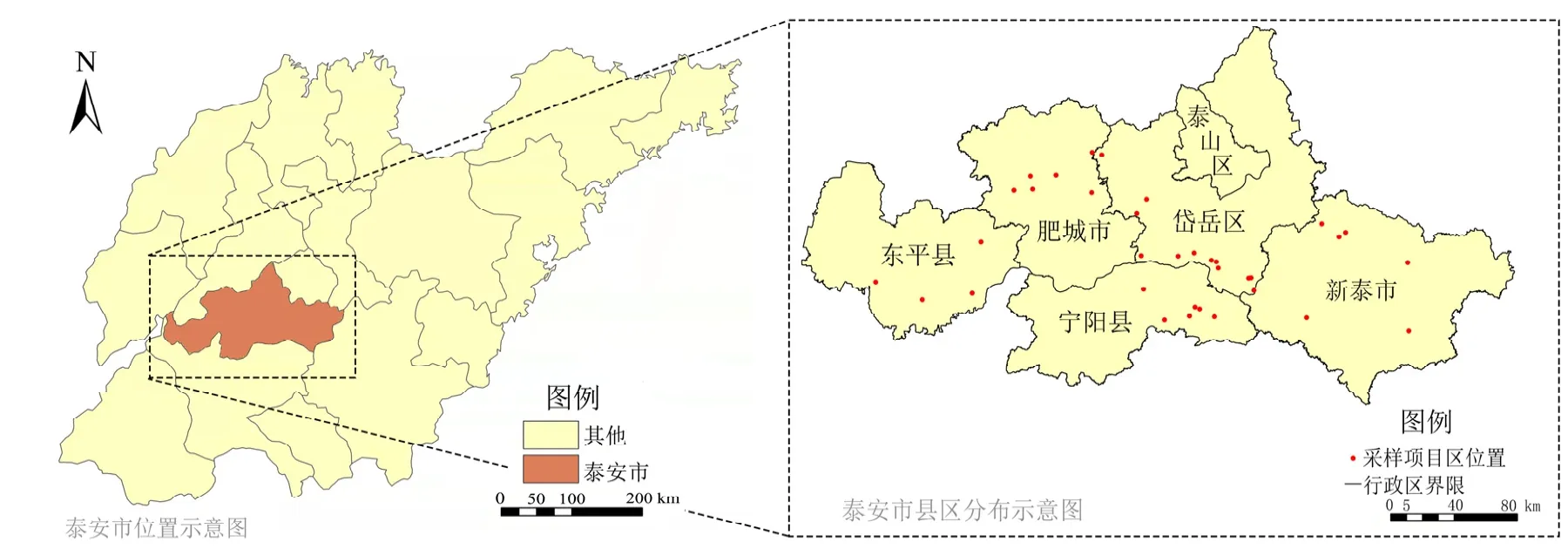
图1 采样项目区位置图Figure 1 Location map of sampling project area
1.2 土壤理化性质测定
采用环刀法测定土壤容重(BD)与含水量(SWC);电极电位法测定土壤pH值;重铬酸钾外加热法测定土壤有机质(SOM);半微量凯氏定氮法测定土壤全氮(TN);碳酸氢钠浸提-钼锑抗比色法测定土壤有效磷(AP);pH 7.0醋酸铵浸提-火焰光度计法测定土壤速效钾(AK)(鲍士旦,2000)。
1.3 土壤微生物测定
细菌培养基采用牛肉膏蛋白胨培养基;放线菌培养基采用改良高氏培养基;真菌培养基采用马铃薯葡萄糖培养基;3种细菌均采用稀释平板计数法计算(吴金水等,2006)。
1.4 DNA提取及高通量测序
使用土壤DNA提取试剂盒(Omega Bio-tek,Norcross,GA,美国)进行土壤微生物总DNA抽提,利用1%琼脂糖凝胶电泳对抽提总DNA纯度和浓度进行检测。对细菌16S rRNA基因的V3—V4区域进行PCR扩增,引物序列为:338F和806R。PCR扩增采用高保真 DNA聚合酶和 ABI GeneAmp®9700型的 PCR仪。PCR扩增体系包括:10×快速Pfu DNA聚合酶(FastPfu)缓冲液2 μL,脱氧核苷三磷酸 dNTPs(2.5 mmol·L−1)2 μL,引物(5 μmol·L−1)0.8 μL,rTaq 聚合酶 0.2 μL,DNA 模板10 ng和无菌水补至总体系20 μL。PCR扩增条件:95 ℃预变性3 min;95 ℃变性30 s,55 ℃退火30 s,72℃延伸30 s,35个循环;72 ℃延伸10 min。PCR扩增结果进行 2%琼脂糖凝胶电泳检测,使用AXYGEN公司的AxyPrepDNA凝胶回收试剂盒切目的片段并且回收PCR产物。PCR扩增产物回收后进行纯化和定量后形成测序文库,对测序文库质检合格后使用Illumina公司的Mi Seq PE 300平台进行双向测序。将土壤样品保存在干冰中送至上海美吉生物科技有限公司进行土壤微生物高通量测序。
1.5 统计分析
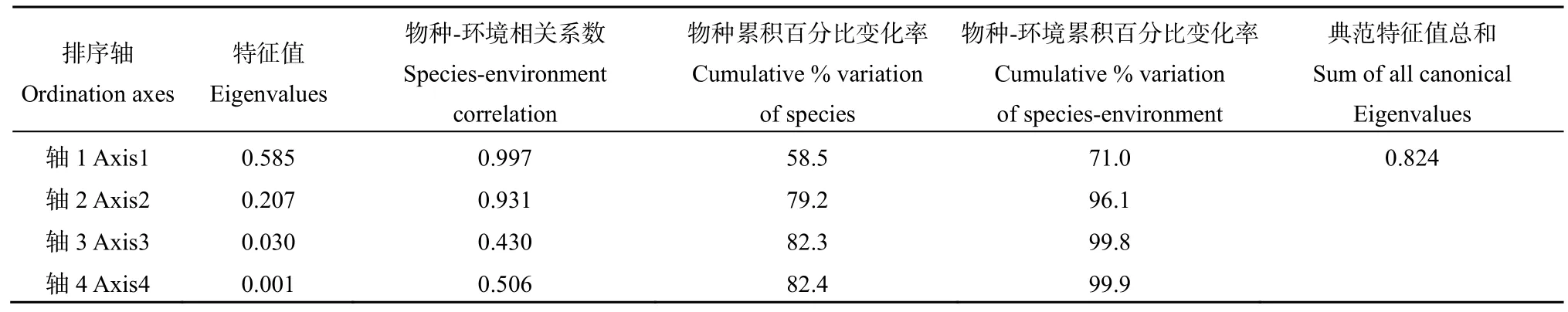
采用QIIME进行土壤细菌群落相关指标分析,采用 Mothur(1.30.2)软件计算 α多样性指数(Shannon、Simpson、Ace、Chao1和Coverage等),采用SPSS 22.0对土壤理化性状进行显著性分析及对土壤理化性状与细菌、真菌、放线菌进行相关性分析,采用Canoco 5.0对土壤群落组成与土壤理化性状进行冗余分析(RDA)。
2 结果与分析
2.1 土壤理化性状
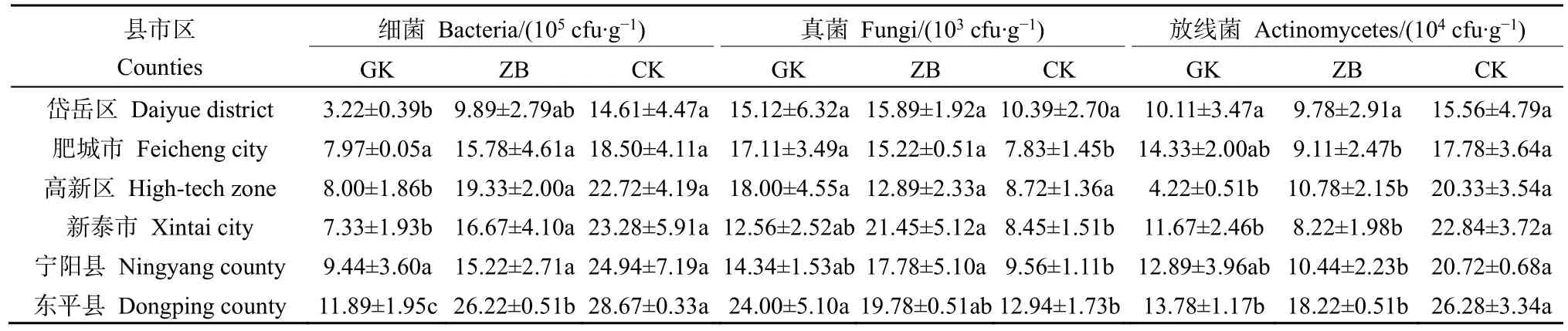
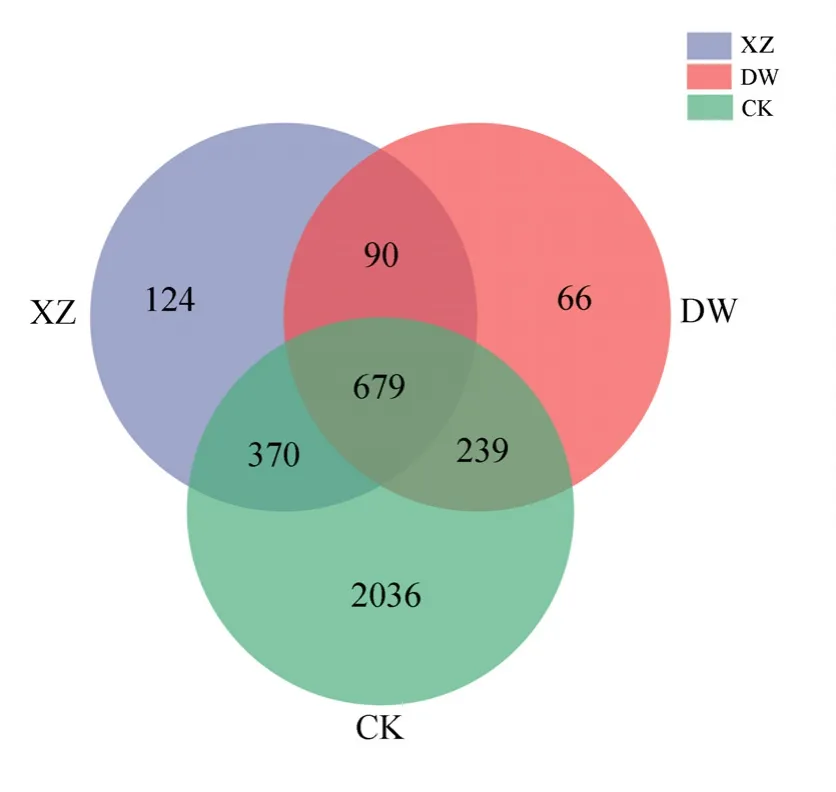
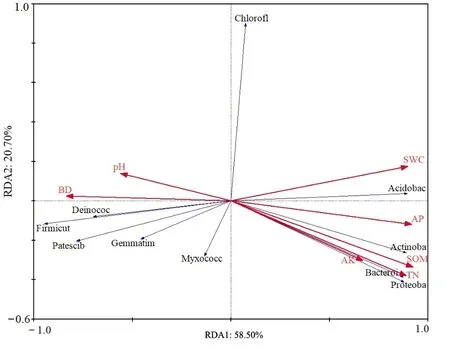
表1总结了工矿复垦(GK)、占补平衡(ZB)两类新增耕地与对照耕地(CK)土壤的基本属性。结果显示,除高新区土壤偏酸性(pH<6.5)外,其余县区大部分土壤偏碱性。两类新增耕地与对照耕地土壤容重大小为:GK>ZB>CK(岱岳区、肥城市、高新区);ZB>GK>CK(新泰市、宁阳县、东平县),与对照耕地相比,岱岳区、肥城市、高新区、新泰市、宁阳县和东平县GK项目土壤容重分别增大了19.59%、17.33%、21.68%、7.38%、22.96%和10.00%,ZB项目土壤容重分别增大了 15.54%、8.67%、13.29%、10.53%、25.19%和16.43%。GK、ZB、CK土壤含水量的变化范围分别为 15.85%—22.03%、13.66%—23.14%、18.07%—26.25%。两类新增耕地土壤有机质、氮磷钾含量均显著低于对照耕地,其中全氮和有效磷含量变化规律明显,均表现为GK 表1 新增耕地和对照耕地土壤基本属性Table 1 Soil basic properties of newly cultivated land and control farmland 微生物数量与土壤质量密切相关,是反映土地恢复过程中土壤状况的重要指标,土壤三大类微生物数量常被作为生物学指标来评价土壤肥力水平的高低。本研究中,新增耕地及对照耕地表层土壤微生物数量总体特征如表2所示,从各类微生物数量的均值来看,总体呈现出两类新增耕地土壤细菌、放线菌数量低于对照耕地,而真菌数量高于对照耕地的趋势。两类新增耕地及对照耕地土壤细菌数量最大值均出现在东平县,分别达到了11.89×105、26.22×105和 28.67×105cfu·g−1,两类新增耕地GK、ZB土壤真菌数量最大值分别出现在东平县(24.00×103cfu·g−1)和新泰市(21.45×103cfu·g−1)。 表2 新增耕地和对照耕地土壤微生物数量Table 2 Number of soil microorganisms in newly cultivated land and control farmland 土壤理化性状与土壤细菌、真菌、放线菌相关性分析(表3)可知,土壤细菌数量与土壤容重呈极显著负相关关系(P<0.01),与土壤含水量、速效钾含量及有机质、全氮、有效磷含量呈显著正相关(P<0.05)或极显著正相关(P<0.01)关系;土壤放线菌数量与土壤容重呈极显著负相关(P<0.01)关系,与土壤含水量及有机质、全氮、有效磷、速效钾含量呈极显著正相关(P<0.01)关系;土壤真菌数量与土壤理化性状的相关性呈现与放线菌数量相反的趋势。 表3 土壤理化性状与土壤细菌、真菌、放线菌相关性分析Table 3 Correlation analysis between soil physio-chemical properties and soil bacteria, fungi and actinomycetes 2.4.1 测序数据分析 基于16S rRNA高通量测序共获得540479个有效序列,基于相似度 97%进行聚类分析,得到3604个OTUs,共34个门、107个纲、264个目、417个科、795个属和1544个种。两类新增耕地和对照耕地土壤共有细菌OTUs数为679,其中工矿复垦(GK-XZ)特有细菌OTUs数为124,占补平衡项目(ZB-DW)特有细菌 OTUs数为 66,对照耕地(CK)为2036(图2)。 图2 新增耕地和对照耕地土壤细菌组成差异Figure 2 Differences of soil bacterial composition between newly cultivated land and control farmland 2.4.2 土壤细菌α多样性和群落组成分析 在 97%相似性水平下对两类新增耕地及对照耕地土壤细菌进行高通量测序,结果表明各样地土壤细菌OTUs覆盖度均大于99%(表4),说明此次测序合理,能够反映新增耕地及对照耕地土壤细菌群落的真实情况。工矿复垦土壤细菌Shannon指数、Ace指数和Chao1指数均大于占补平衡土壤,Simpson指数相反。与对照耕地相比,两类新增耕地土壤细菌Shannon指数、Ace指数和Chao1指数小于对照耕地,Simpson指数大于对照耕地。两类新增耕地土壤细菌Shannon指数、Simpson指数、Ace指数和 Chao1指数均与对照耕地差异显著(P<0.05)。 表4 新增耕地和对照耕地土壤细菌α多样性Table 4 The α-diversity indexes of soil bacteria in new cultivated land and control farmland 由图3可以看出,在门分类水平上,两类新增耕地与对照耕地的优势菌门主要有放线菌门(20.65%—40.40%)、厚壁菌门(9.42%—34.98%)、变形菌门(10.52%—21.13%)、绿弯菌门(7.63%—33.73%)、酸杆菌门(2.07%—7.87%)、芽单胞菌门(0.49%—9.08%)、异常球菌-栖热菌门(0.00%—10.27%)、拟杆菌门(0.51%—3.00%)、髌骨细菌门(0.74%—2.87%)、粘球菌门(0.08%—2.00%)。与对照耕地相比,两类新增耕地土壤放线菌门、变形菌门、酸杆菌门、拟杆菌门数量低于对照耕地,厚壁菌门、芽单胞菌门数量高于对照耕地。这表明,土地整治导致的土壤生态环境改变会使得土壤细菌群落中的一些物种逐渐被某些更适应在新环境中生长的物种替代,土壤微生物群落因与土壤间的强相互关系,当其发生变化时将影响新增耕地土壤的恢复过程。 图3 新增耕地和对照耕地土壤细菌群落组成Figure 3 Composition of soil bacterial community in newly cultivated land and control farmland 2.4.3 土壤细菌群落结构与土壤理化性状的冗余分析 通过 RDA揭示了土壤细菌群落组成结构与土壤理化性质之间的关系,结果(表5,图4)发现,门分类水平上,轴 1和轴 2分别解释了变量的58.50%和20.70%,本文所选的7个土壤理化性状的特征值共解释了82.40%的总方差(表5)。置换检验的冗余分析显示(表6),SOM对土壤细菌群落结构的影响最大(P=0.0080),单独解释量达52.00%。 表5 冗余分析Monte Carlo置换检验结果Table 5 Results of Monte Carlo permutation test 图4 土壤细菌群落与土壤理化性状冗余分析Figure 4 Redundancy analysis for bacterial community and soil physico-chemical 表6 新增耕地及对照耕地土壤细菌群落冗余分析Table 6 Redundancy analysis of bacterial communities in newly cultivated land and control farmland 土壤理化性质的变化是影响微生物群落特征发生变化的主要因素,土壤微生物数量、多样性及群落组成等群落特征均可反映土壤质量(纪立东等,2020;张振佳等,2021;周虹等,2022)。土地整治使得土壤生物和非生物因素在相互作用的过程中改变了土壤的理化性质和微生物群落(Legrand et al.,2018;Wang et al.,2018)。土地整治类型不同,土壤理化性状存在一定差异。本研究发现,两类新增耕地土壤pH值和土壤含水量总体上与对照耕地差异不显著,但土壤容重、有机质及氮磷钾含量呈现出了较大差异。两类新增耕地土壤容重总体上显著高于对照耕地,有机质及氮磷钾含量显著低于对照耕地,其中全氮、有效磷含量趋势明显,均为 GK 利用Illumina MiSeq高通量测序技术对两类新增耕地的土壤细菌α多样性和群落组成进行研究,在评价土壤细菌群落多样性时主要采用Shannon指数、Simpson指数、Ace指数及Chao1指数,Shannon和Simpson指数可以反映土壤微生物群落物种多样性;Ace和Chao1指数可以反映土壤微生物群落中物种数量的丰富度(罗友进等,2020)。一般而言,多样性指数越高,微生物群落结构越复杂,生态系统相对越稳定(杨君珑等,2018)。本研究发现,两类新增耕地土壤多样性指数显著低于对照耕地,造成这种差异的原因在于两类新增耕地是通过土壤重构形成的,而重构土壤彻底改变了土壤物理结构及养分含量,导致其土壤质量远低于对照耕地,使得作为微生物生存活动的土壤环境状况较差及微生物生命代谢来源的养分资源缺乏,不利于某些偏好微生物的生长繁殖,致使两类新增耕地土壤微生物多样性较低。工矿复垦土壤细菌 Ace指数及Chao1指数高于占补平衡土壤,表明工矿复垦土壤细菌群落中物种数量的丰富度高于占补平衡土壤。两类新增耕地与对照耕地的优势菌门均为放线菌门、厚壁菌门、变形菌门、绿弯菌门和酸杆菌门等,其他研究人员也在不同类型的土壤中发现了这些细菌优势菌门(Wang et al.,2020;刘洋等,2016;程分生等,2021;郑兰香等,2021)。这一结果说明,虽然两类新增耕地为重构土壤,但土壤中执行生态功能的优势菌门依然存在,并表现出了较高的相对丰度,因此其本身具备土壤微生物功能恢复的构建条件。放线菌门和变形菌门细菌生态幅宽广、环境适应强,是陆地生态系统的主要优势类群(程分生等,2021),因此,土地整治工程中人为扰动强度大小对其优势种群地位的影响较小。放线菌门能够降解纤维素和几丁质,是土壤养分供给的主要来源(李宏等,2021),本研究表明,对照耕地的放线菌门细菌相对丰度显著高于两类新增耕地(图3),有机质及养分含量也高于两类新增耕地(表1),研究结果证实了放线菌门对土壤养分的贡献;RDA分析发现变形菌门与SOM相关性较大,该结果支持变形菌门与碳利用方面有关的结论(郑兰香等,2021)。两类新增耕地中厚壁菌门细菌相对丰度显著高于对照耕地(图3),这是由于厚壁菌门细菌为耐受强烈的外界环境变化而进化出了较厚的细胞壁,保护自身在异常条件下可以更好地存活(郑文玲等,2021),这也反映出了某些微生物在应对环境变化时具有相应的响应策略。 影响土壤微生物群落结构变化的环境因子多为pH、有机质/有机碳含量(Thakur et al.,2019;王蓥燕等,2019),本研究得出 SOM 含量是影响土壤微生物群落结构变化的主要因素,SOM含量能解释细菌群落结构变化的52.00%(表6),SOM作为土壤固相部分的重要组分,与新增耕地土壤的结构、通气、渗透及缓冲性能密切相关。土壤pH对土壤微生物群落结构的影响较为复杂,本研究中土壤pH值与细菌群落不存在显著相关性,这可能是由于两类新增耕地土壤pH值与对照耕地相比变化不大且差异不显著所致(表1),另外,与土壤类型、土壤性质、人为干扰程度及采样时间等因素间的交互作用也有一定关系。前人的研究中也会出现pH的作用不突出的情况,如丁嘉宁等(2020)对煤矿区复垦土壤研究发现,土壤全磷、有机质等是影响土壤群落结构差异的主要因素;裴广廷等(2021)对草地土壤的研究得出微生物生物量碳和细根生物量的改变是土壤微生物群落结构变化的主要影响因素,这些研究均表明pH值并非一直是影响土壤微生物群落结构变化的主导因子。因此,土壤有机质成为两类新增耕地重点调控的指标,今后应通过大量施加高碳基有机肥或高碳基有机改良材料的方式,快速增加土壤有机碳,从而为土壤微生物生命活动提供充足的碳源及更适宜的生存环境,有利于增加微生物数量,加速动植物残体等有机物料的分解、转运,最终达到新增耕地土壤质量有效改善、作物持续稳定增产的效果。 (1)两类新增耕地土壤容重高于对照耕地;有机质及氮磷钾含量低于对照耕地,其中全氮和有效磷含量变化规律明显,均为CK>ZB>GK。 (2)两类新增耕地和对照耕地中细菌数量占绝对优势;两类新增耕地细菌、放线菌数量低于对照耕地,而真菌数量高于对照耕地;细菌、放线菌数量与土壤有机质及养分呈显著正相关,真菌则相反。 (3)两类新增耕地与对照耕地的优势菌门均为放线菌门、厚壁菌门、变形菌门、绿弯菌门、酸杆菌门等,两类新增耕地放线菌门相对丰度均低于对照耕地,厚壁菌门相对丰度高于对照耕地。RDA分析发现,土壤有机质是影响土壤微生物群落变化的主要因子。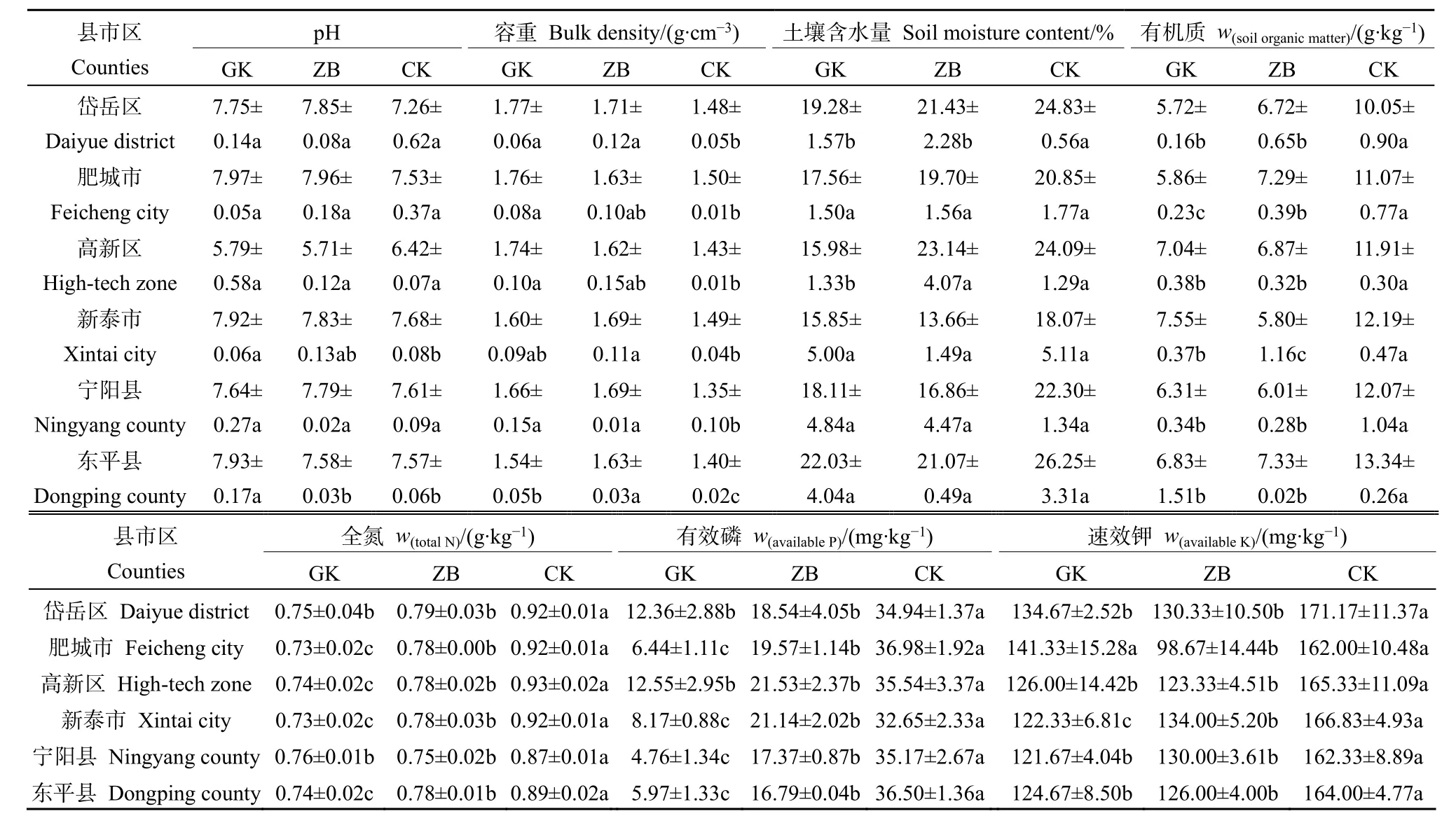
2.2 土壤微生物数量
2.3 土壤理化性状与土壤微生物数量相关性分析

2.4 土壤细菌多样性和群落组成



3 讨论
4 结论
