野杏经济性状SSR 关联分析及优异等位变异挖掘
2022-07-20董胜君温佳星卢彩云陈建华刘权钢
董胜君,温佳星,卢彩云,陈建华,刘权钢,金 玲
(沈阳农业大学 林学院,辽宁 沈阳 110866)
野杏Armeniaca vulgaris是蔷薇科杏属植物中普通杏的一个变种,落叶乔木,一般生长在亚热带地区,丘陵地带,主要分布在我国北部地区,栽培或野生[1]。野杏喜光,对土壤的适应性强,根系发达,是重要的造林先锋树种,具有较高的生态价值。其果实含糖量高;果肉中黄酮类成分的质量分数高达269.7 mg/g,具有防动脉硬化、抗脂肪氧化及活化大脑等功能;杏仁可供食用及作为预防心血管疾病和降低胆固醇等的药物[2]。此外,野杏也可作为观赏树种栽培,具有重要的经济价值和社会价值。
野杏分布广泛,种质资源丰富,但多处于野生、半野生状态,自交不亲和、种内杂交现象普遍,易形成较高水平的遗传变异和较低水平的连锁不平衡[3]。因此,在进行野杏资源的开发利用时,首先要进行优良亲本的选育。传统的野杏育种方法主要为凭借育种经验根据表型选择后代进行栽培,表型性状与基因型之间的关系不明确,无法对目的性状进行直接选择,选择效率低,准确性差,且因生长周期长,选育出优良的野杏品种需要较长时间。简单重复序列(SSR)亦称微卫星DNA,SSR 标记的灵敏度高、多态性高且技术简便易操作,已在密叶红豆杉Taxus contorta[4]、美国红枫Acer rubrum[5]等树种的研究中得到应用,是分子标记辅助育种以及发掘优良性状等位变异的较好的标记方法[6],艾鹏飞等[7]建立和优化了仁用杏的SSR 标记反应体系,何天明等[8]则利用SSR 标记对新疆栽培杏品种亚群进行了分子系统学研究。基于连锁不平衡将标记与性状关联起来的分析方法被称为关联分析[9],该方法精度高、周期短,不用构建载体,可同时检测同一位点的多个等位基因[10],已被广泛应用于大麦Hordeum vulgare[11]、大豆Glycine max[12]、玉米Zea mays[13]等 作 物 及 板 栗Castanea mollissima[14]、 桃 树Amygdalus persica[15]、野苹果Malus sieversii[16]等木本植物的研究中,为复杂的数量性状基因定位和分子标记辅助育种提供了快捷、有效的途径。但通过SSR 关联分析仅能在分子水平上找到与性状相关联的位点,在选育优良亲本个体时,仍要进行表型效应分析,进而挖掘出典型载体材料。目前,表型效应分析及载体材料挖掘多见于作物研究中,如陆地棉Gossypium hirsutum[17]、辣椒Capsicum annuum[18]、大 麦Hordeum vulgare[11]、大豆Glycine max[12]等,木本植物中仅见椰子Cocos nucifera[19]、小 叶 杨Populus simonii[20]、杉木Cunninghamia lanceolata[21]等少数树种有相关研究。在杏属植物的研究中:蒋江照等[22]利用关联分析研究了有效积温对小白杏果实品质的影响;张欣[23]对西伯利亚杏SSR 标记与表型性状进行关联分析,找到了与表型相关的SSR 位点;表型效应分析仅见金玲[24]挖掘了携带西伯利亚杏果实优异等位基因的典型载体材料。
目前,应用关联分析发掘与野杏重要经济性状相关联位点的研究报道较少,野杏表型效应分析的相关研究更是鲜见报道。本研究中以40 个野杏无性系为研究对象,利用120 对SSR 引物进行扩增、标记,采用Tassel 软件中的GLM 模型对SSR 标记与20 个重要经济性状进行关联分析,并进行等位变异表型效应分析,发掘携带野杏优良经济性状基因的典型载体材料,以期全面系统地分析已有的育种材料,了解表型性状与基因型之间的联系,为野杏分子标记辅助育种和良种选育工作提供参考。
1 材料与方法
1.1 试验材料
试验材料取自沈阳农业大学喀左县山杏国家林木种质资源保存库。供试野杏无性系编号及种源信息见表1。

表1 供试野杏无性系编号及种源信息Table 1 The information of No. and provenances of tested A. vulgaris
1.2 试验方法
1.2.1 SSR 引物数据获得
基于沈阳农业大学山杏课题组前期对野杏群体的SSR 分子标记开发,委托北京赛百盛基因技术有限公司合成SSR 引物,进行PCR 扩增(于2017 年完成),从中筛选出120 对扩增效果好、多态性高的引物数据。
1.2.2 经济性状测定
2018—2020 年,在试验地观测40 个野杏优良无性系,每个无性系选择3 次重复。成熟期采集果实,每样本采集20 个果实,用数显卡尺测量果长(fruit length,FL)、果宽(fruit width,FW)、果厚(fruit thickness,FT)、果 肉 厚 度(flesh thickness,FLT)、核长(nutlet length,NL)、核宽(nutlet width,NW)、核厚(nutlet thickness,NT)、仁长(kernel length,KL)、仁宽(kernel width,KW)、仁厚(kernel thickness,KT),用电子天平测定单果质量(single fruit mass,SFM)、单核质量(single nutlet mass,SNM)、单仁质量(single kernel mass,SKM),计算果形指数(fruit-form index,FFI)、核形指数(nucleation index,NCI)、仁形指数(kernel-form index,KFI)、出核率(nutlet rate,NR)、出仁率(kernel rate,KR)、产果量(fruit yield,FY)、产核量(nutlet yield,NY)。
1.3 数据处理
1.3.1 关联分析
使用Structure 2.3 软件计算各无性系的Q值。运用TASSEL 2.1 软件的GLM 模型,以Q值作为协变量,进行SSR 标记与经济性状的关联分析。
1.3.2 等位变异的表型效应分析
参考无效等位基因(null allele)法,计算关联标记的各等位变异的表型效应值,获得与野杏重要经济性状关联等位变异的表型效应,再根据表型效应值找出最优无性系,获得典型载体材料。

式中:ai为第i个等位变异的表型效应值;xij为携带第i个等位变异的第j个材料的性状表型测定值;ni为具有第i个等位变异的材料数量;Nk为携带无效等位变异的第k个材料的表型测定值;nk为具有无效等位变异的材料数量。若ai为正,则认为该等位变异为正向等位变异,反之为负向等位变异。
2 结果与分析
2.1 野杏经济性状与SSR 标记的关联分析
供试野杏样本的经济性状均值见表2。与野杏经济性状显著关联的SSR 位点见表3。由表3 可知,共检测出43 个与野杏经济性状极显著关联的SSR 位点(P<0.01),有9 个与产果量呈极显著相关联的位点,有7 个与仁厚呈极显著相关联的位点;共检测出64 个与野杏经济性状显著关联的SSR 位点(P<0.05),除果形指数外其他性状均与多个SSR 关联位点显著相关,与核宽显著关联的位点最多(18 个),产果量次之(17 个)。有40 个位点分别与多个性状相关联,其中:P1 位点与仁长、单仁质量等7 个性状相关联;X36 位点与核厚、出核率等7 个性状相关联;L62H 位点与核长、核宽等6 个性状相关联;Y86、X16H、Y17、L45H、L7 位点分别与5 个性状相关联;L49H、L57H、X19H 分别与4 个性状相关联;L25、L46、L59H、L62、P39H、X15H、X39H、X132、Y11、Y65 分别与3 个性状相关联。

表2 供试野杏无性系20个经济性状均值Table 2 Mean value of 20 economic traits of tested A. vulgaris

续表2Continuation of Table2

表3 与供试野杏经济性状显著关联的SSR位点Table 3 SSR loci associated significantly with economic traits of tested A. vulgaris
在20 个经济性状中,与果实性状相关联的位点有36 个,变异解释率为17.03%~48.96%,其中,果长、果宽、果厚、果肉厚度、果形指数、单果质量分别与Y17(47.35%)、P39H(40.36%)、X11H(39.54%)、X36H(48.96%)、X53H(37.46%)、Y17(47.17%)位点的变异解释率最高。与果核性状相关联的位点有42 个,变异解释率为15.67%~57.21%,其中,核长、核宽、核厚、核形指数、单核质量、出核率分别与L7(47.17%)、L7(57.21%)、X15H(54.73%)、Y11(31.81%)、L7(36.47%)、X42H(56.35%)位点的变异解释率最高。与果仁性状相关联的位点有37 个,变异解释率为14.77%~61.42%,其中,仁长、仁宽、仁厚、仁形指数、单仁质量、出仁率分别与Y17(42.69%)、L7(43.96%)、X34H(54.60%)、X34H(36.75%)、X39H(61.42%)、L79H(56.98%)位点的变异解释率最高。与产量性状相关联的位点有31 个,变异解释率为12.21%~77.78%,其中,X44H 标记与产果量、产核量的最高变异解释率分别为74.21%、77.78%。
2.3 野杏经济性状SSR 关联位点等位基因的表型效应分析
2.3.1 果实性状SSR 关联位点等位基因的表型效应值
果实性状SSR 关联位点等位基因的表型效应值见表4。由表4 可知:与果长相关的表型效应有10 个,表型效应值为-0.73 ~0.43,负向效应和正向效应的典型载体材料分别为241 号(Y65-158)和164 号(Y17-130)无性系;与果宽相关的表型效应有18 个,均为负向效应,最低表型效应值为-0.87,典型载体材料为210 号(P39H-150)无性系;与果厚相关的表型效应有21 个,表型效应值为-0.55 ~0.89,负向效应和正向效应的典型载体材料分别为210 号(X11H-122)和164 号(Y86-146)无性系;与果肉厚度相关的表型效应有32 个,表型效应值为-0.32 ~0.43,负向效应和正向效应的典型载体材料分别为241 号(Y65-158)和211 号(X36H-110/X36H-122)无性系;没有与果形指数相关的表型效应;与单果质量相关的表型效应有10 个,表型效应值为-7.80 ~6.07,负向效应和正向效应的典型载体材料分别为232号(Y17-120)和164 号(Y17-130)无性系。
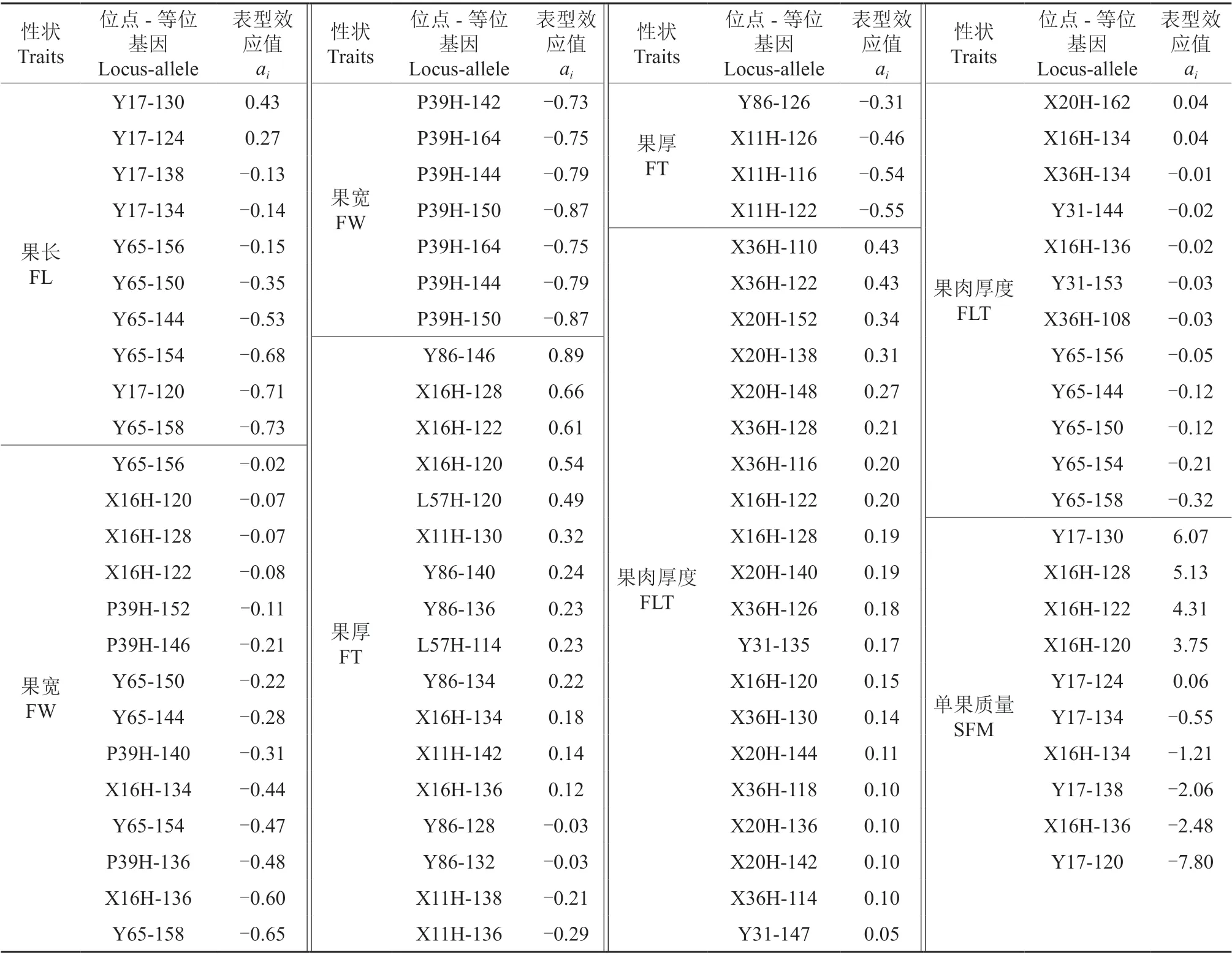
表4 供试野杏果实性状SSR关联位点等位基因的表型效应值Table 4 Phenotypic effects of alleles of SSR loci associated with fruit traits of tested A. vulgaris
2.3.2 核性状SSR 关联位点等位基因的表型效应值
核性状SSR 关联位点等位基因的表型效应值见表5。由表5 可知:与核长相关的表型效应有16个,表型效应值为-0.84 ~1.02,负向效应和正向效应的典型载体材料分别为210 号(L7-117)和207 号(Y86-126)无性系;与核宽相关的表型效应有58 个,表型效应值为-0.58 ~1.05,负向效应和正向效应的典型载体材料分别为210 号(L7-117)和207 号(Y86-126)无性系;与核厚相关的表型效应有2 个,均为负向效应,最低表型效应值为-0.08,典型载体材料为210 号(L49H-123)无性系;与核形指数相关的表型效应有8 个,表型效应值为-0.11 ~0.06,负向效应和正向效应的典型载体材料分别为197 号(L62-144)和248 号(Y11-158)无性系;与单核质量相关的表型效应有7 个,表型效应值为-0.72 ~0.60,负向效应和正向效应的典型载体材料分别为210 号(L7-117)和261 号(L57H-120)无性系;与出核率相关的表型效应有22 个,表型效应值为-9.96 ~9.29,负向效应和正向效应的典型载体材料分别为171号(X16H-122)和120 号(X36H-108)无性系。

表5 供试野杏核性状SSR关联位点等位基因的表型效应值Table 5 Phenotypic effects of alleles of SSR loci associated with nuclear traits of tested A. vulgaris
2.3.3 仁性状SSR 关联位点等位基因的表型效应值
仁性状SSR 关联位点等位基因的表型效应值见表6。由表6 可知:与仁长相关的表型效应有5个,表型效应值为-0.20 ~0.17,负向效应和正向效应的典型载体材料分别为232 号(Y17-120)和164 号(Y17-130)无性系;与仁宽相关的表型效应有22 个,表型效应值为-0.29 ~0.17,负向效应和正向效应的典型载体材料分别为198 号(L7-123)和207 号(L46-153)无性系;与仁厚相关的表型效应有38 个,表型效应值为-0.07 ~0.37,负向效应的典型载体材料为164 号(P21-128)或162 号(X19H-130),正向效应的典型载体材料为248 号(P39H-144)无性系;与仁形指数相关的表型效应有11 个,表型效应值为-0.19 ~0.14,负向效应和正向效应的典型载体材料分别为207号(L62H-132)和247 号(X34H-144)无性系;与单仁质量相关的表型效应有32 个,表型效应值为-0.19 ~0.42,负向效应和正向效应的典型载体材料分别为210 号(L7-117)和207 号(Y86-126)无性系;与出仁率相关的表型效应有18 个,表型效应值为-8.01 ~8.99,负向效应和正向效应的典型载体材料分别为245 号(L25-124)和167 号(L25-112)无性系。

表6 供试野杏仁性状SSR关联位点等位基因的表型效应值Table 6 Phenotypic effects of alleles of SSR loci associated with kernel traits of tested A. vulgaris
2.3.4 产量性状SSR 关联位点等位基因的表型效应值
产量性状SSR 关联位点等位基因的表型效应值见表7。由表7 可知:与产核量相关的表型效应有36 个,表型效应值为-145.61 ~582.68,负向效应和正向效应的典型载体材料分别为263 号(X21H-134)和162 号(X19H-130)无 性 系;与产果量相关的表型效应有40 个,表型效应值为-1 501.23 ~4 429.39,负向效应和正向效应的典型载体材料分别为198 号(Y54-108)和162 号(X19H-130)无性系。

表7 供试野杏产量性状SSR关联位点等位基因的表型效应值Table 7 Phenotypic effects of alleles of SSR loci associated with yield traits of tested A. vulgaris
3 结论与讨论
本研究中基于120 个SSR 标记通过关联分析获得了与野杏20 个经济性状相关联的64 个SSR位点,通过表型效应分析发掘了406 个优异等位变异和18 个典型载体材料。
关联分析已被广泛应用于探索表型性状与基因型之间的关系,近年来已有较多相关报道[25-26]。使用该方法可以同时对多个性状进行定位,使等位变异更加丰富,还可以得到更多的关联位点,能够发现使用相关分析和连锁分析等技术手段无法得到的、隐藏在表型数据之下的遗传变异。本研究中通过关联分析检测到了多个与野杏经济性状相关联的位点。解释率越大,性状与位点的相关性越强,关联结果中与仁厚关联位点的平均变异解释率最大(45.53%),与单核质量关联位点的平均变异解释率最小(23.81%),说明供试野杏群体遗传水平差异较大。有17 个位点对表型变异的解释率在50%以上,其中与产果量和产核量关联的X44H 位点的变异解释率最高,分别为74.21%和77.78%,均在70%以上。除果形指数外的19 个性状均有多个SSR 关联位点,说明这些性状可能受到多基因调控或者受连锁群内及连锁群间的连锁调控,这与其他学者对大麦Hordeum vulgare[11]、辣椒Capsicum annuum[18]、水稻Oryza sativa[27]的研究结果类似。另外,有44 个位点分别与多个性状相关联,前人在研究西伯利亚杏Armeniaca sibirica[24]、荷花NelumboAdans.株型[28]、玉米Zea mays[29]时也发现与多个性状存在“一因多效”或连锁关系的位点基因,这些与同一位点相关联的多个性状可能存在连锁遗传。本研究中最终获得了多个与野杏经济性状相关联的位点,位点基因与性状之间的关系多样且复杂。
本研究中在关联分析的基础上,对得到的关联位点进一步进行解析,分析其表型效应,挖掘优异等位变异。Breseghello 等[30]在2006 年首次提出用无效等位基因的方法分析表型效应,随后该方法被广泛应用于挖掘携带优异等位变异的典型载体材料。Zhou 等[19]在椰子Cocos nucifera果肉脂肪酸含量的研究中挖掘出了具有最大正向效应和最大负向效应的典型载体材料;卫尊征[20]在获得17 个与小叶杨P. simonii叶、茎和根性状极显著关联位点的基础上进行表型效应分析,挖掘出了相关典型载体材料;张胜龙[21]在研究杉木Cunninghamia lanceolata材质性状变异规律时挖掘出了3 个材质性状和3 个木材生长性状的典型载体材料;在杏属植物的相关研究中,仅见金玲[24]采用155 个SSR 分子标记对32 个西伯利亚杏A. sibirica无性系的表型性状进行关联分析,得到49 个与13 个经济性状显著关联的位点,并挖掘出6 个携带优异等位变异的典型载体材料。本研究结果表明,所发掘的406 个有效等位变异中,122 个有效等位变异同时对多个性状产生正向或负向效应。在17 个典型载体材料中,210 号典型载体材料携带了7 个优异等位变异,207 号、164 号典型载体材料携带了5 个优异等位变异,162 号典型载体材料携带了3 个优异等位变异,232 号、241 号、248 号、198 号等典型载体材料均携带了2 个优异等位变异。这些载体材料携带优异基因较多,属于优异亲本。可以利用发掘出的典型载体材料进行有性杂交和连续回交,聚合更多的优异基因。例如,利用164 号无性系和162 号无性系杂交选育大果且高产的品种,利用207 号无性系和248号无性系杂交选育仁用杏品种等。这些优异等位变异的聚合可以实现多个目标性状的同步改良,从而加速野杏优良品种选育的进程。
本研究中通过关联分析和表型效应分析找到了与野杏经济性状相关联的SSR 位点及优异等位变异,但各等位变异调节野杏经济性状的机制,各等位变异间的连锁遗传关系等尚未有明确的答案,后续可以利用克隆、遗传转化、酵母单双杂交等基因工程技术对这些等位变异的功能进行验证,并对其相互作用进行研究。
