华顶山3 种生境的云锦杜鹃毛根发育与菌根侵染比较研究
2022-07-16王日华KaboreArthurFabrice周午祖轩童璐朗月何育栋孟凡博张艳华孙立夫
王日华,Kabore M.Arthur Fabrice,2,周午祖轩,童璐,朗月,何育栋,孟凡博,张艳华,孙立夫
(1.绍兴文理学院 生命科学学院,浙江 绍兴 312000;2.福建农林大学 生命科学学院,福建 福州 350002;3.绍兴职业技术学院,浙江 绍兴 312000)
陆地植物生长发育所需的营养物质主要依靠植物地上、地下部分与外界环境之间的物质交换,根系决定了植物利用土壤资源的能力[1]。有些植物的根与土壤真菌形成菌根,菌根共生能分解复杂的有机物[2],在改善植物营养、调节植物代谢、增强植物抗逆性等方面发挥着重要的作用[3-4]。毛根发育和菌根侵染是宿主植物增强与其他物种竞争能力的重要指标,在没有菌根真菌侵染的情况下,黑麦草Lolium perenne和绒毛草Holcus lanatus的生长情况相似,但是都被丛枝菌根(Arbuscular mycorrhiza,AM)真菌侵染后,黑麦草在生长、根冠比、P 和K的吸收能力方面均高于绒毛草[5],植物种间的竞争平衡被打破。多个宿主植物之间由菌根联结形成的菌根网络,也可能存在光合产物、N 和P 等营养物质的单向或双向传递[6]。在群落水平上,物种的菌根联结是植物与相邻伴生种关系的重要驱动因素,Chen 等在我国亚热带森林的研究中发现,附近AM 幼树和成株的出现显著抑制了外生菌根(Ectomycorrhiza,ECM)幼苗的存活,但是相邻的ECM 植物则大大提高了AM 幼苗的存活率[7]。在美国加州和犹他州草地,伴生种裂稃燕麦Avena barbata和大麦状雀麦Bromus hordeaceus能明显改变当地优势植物根部微生物群落的组成,使其由菌根菌转变为非菌根菌,进而导致当地优势植物竞争力的降低[8]。因此,伴生种的变化可能会直接影响宿主植物的生长和土壤真菌的组成。伴生种也可能通过改变土壤的理化性质,间接地影响宿主根系的发育和地下菌根的形成,如在硝态氮(NO3--N)和有机氮多的环境中,杜鹃花类菌根(Ericoid mycorrhiza,ERM)真菌菌株的生长数量比在铵态氮(NH4+-N)甚至不含N 的环境中更高[9];增加土壤有机质含量,有利于提高蓝莓Vacciniumspp.的菌根侵染率和根系活力[10]。
ERM 是杜鹃花科Ericaceae 植物最主要的菌根类型,由纤细的毛根(hair roots)与土壤真菌形成典型的菌根结构,即在表皮和皮层细胞中有菌丝卷(圈,coil),大部分毛根集中分布在土壤表层[11]。云锦杜鹃Rhododendron fortunei是我国特有种,为常绿的灌木或小乔木,高可达12 m,具有ERM 结构,在浙江天台的华顶山有自然分布的种群。每年5 月杜鹃花盛开时,都会吸引大量游客。云锦杜鹃常作为优势种,与其他木本或草本植物相伴而生。但是,该区域的云锦杜鹃老龄植株有逐渐衰落的趋势,管康林等认为很可能是低灌层箬竹Indocalamus tessellatus扩张导致的[12]。箬竹是禾本科Poaceae 植物,常生长于林缘、林下和沟谷等处[13],根系发达。云锦杜鹃的毛根和箬竹的根在土壤中生长的位置相同。
因此,本研究在浙江华顶山,分别在由箬竹伴生、茶Camellia sinensis伴生和以无下木伴生为对照的云锦杜鹃生长的3 种生境中,采集云锦杜鹃的毛根,收集样株树冠下的凋落物和根围土壤。在不同伴生种的生境下,比较云锦杜鹃毛根发育、菌根侵染,土壤pH 值、NH4+-N 和NO3--N 的差异,以及C、N、P 及其计量比在土壤和凋落物中的差异,探索云锦杜鹃地下菌根对伴生种变化特别是箬竹扩张的影响,为华顶山云锦杜鹃种群的保护和可持续发展提供参考。
1 材料与方法
1.1 研究地点和样株的确定
研究地点位于浙江天台山的华顶山(121°05' E,29°15' N),云锦杜鹃生长在海拔900~ 1 100 m,坡度约为25°,坡向多为西南坡,生境面积约为6 hm2[12,14]。属亚热带中部季风气候,气候温暖,光照充足,年平均气温为17℃,最低温为-5℃,最高温为39℃,年降水量为1 700 mm[15]。云锦杜鹃林下以箬竹最多,草本植物以禾本科和菊科Compositae 种类较多,还有部分蕨类和苔藓等。
2020 年7 月26 日—8 月4 日,在华顶山海拔900 m 以上,选择云锦杜鹃种群生长的3 种生境:箬竹伴生、茶伴生和无下木伴生,在3 种生境中各选择20 株主干基部直径为16~ 28 cm 的云锦杜鹃样株,株间距应不少于10~ 15 m。
1.2 毛根的采集和处理
在3 种生境中确定60 株云锦杜鹃样株后,每一个样株采集1~ 2 根直径2 cm 左右、长30~ 70 cm 带有毛根的侧根,密封,4℃冷藏。同时采集宿主周围的箬竹和茶的根,同样密封,冷藏。将采集回来的侧根,用蒸馏水浸泡冲洗多次,沥干后,剪下云锦杜鹃的活性毛根,剩余的根适当风干,各自称鲜质量。根据称取的侧根和毛根质量,计算毛根质量比,毛根质量比=毛根的质量/根的质量,并以毛根质量比作为说明毛根发育情况的指标。
1.3 土壤的采集和处理
采集每一个样株的毛根时,也同时收集各样株5~ 20 cm 土层处侧根周围的土壤,在样株下半径为2 m 范围内收集凋落物,4℃冷藏,密封。采集的土壤和凋落物样品各60 份。3 种生境中的凋落物样品分别完全混匀后,带回实验室烘干至恒质量;按生境分别将土壤样品完全混匀后,分为新鲜样品、风干样品和烘干样品,分别过筛(孔径0.149 mm),密封保存。
1.4 醋酸墨水染色及显微结构观察
从采集的每一个样株的毛根中随机挑选30 条1.5 cm 左右长的毛根根段,对云锦杜鹃(60 株)、箬竹(5株)和茶(5 株)的毛根根样分别进行醋酸墨水染色[16],在Nikon(Eclipse E800)光学显微镜下观察根的显微结构,并用根段频率法计算云锦杜鹃各样株的侵染率[17]。
1.5 生态化学计量指标的测定
土壤和凋落物总C(TC)和总N(TN)用Vario ELⅢ元素分析仪测定;总P(TP)采用钼锑抗比色法测定;土壤NH4+-N 和NO3--N 用KCl 提取,用全自动流动注射分析仪(AA3)测定;水∶土=2.5∶1,电位法测定土壤pH 值。每个样品测定均重复两次,取平均值。
1.6 数据分析
实验所得数据用SPSS 23.0 进行ANOVA 分析和LSD 多重比较。
2 结果与分析
2.1 不同生境中云锦杜鹃毛根发育水平的比较
毛根是云锦杜鹃形成菌根的载体,毛根发育的好坏直接关系到宿主菌根的形成和功能的发挥,本研究以各样株毛根质量比作为衡量毛根发育的指标。在野外采样时发现,在箬竹伴生的生境中,箬竹都是紧密地长在云锦杜鹃主干周围,如图1A,竹鞭是箬竹细长的地下茎,一般生长在地下5~ 15 cm 土层处[18],鞭上有节,节上会生根(图1E),并能横向扩展成网状,抢占了云锦杜鹃毛根发育的空间。在华顶山,云锦杜鹃和箬竹在地上和地下空间的竞争都非常激烈,与之相比,茶的根系并不发达(图1F),与无下木的生境一样,云锦杜鹃毛根的发育都很好,如图1D,其生活根的质量比在茶伴生和无下木伴生这两种生境之间没有差异,都远高于箬竹伴生生境(P<0.01)。

图1 不同伴生种的野外采样地生境和3 种植物的根Figure 1 Sampling habitat with different accompanying species and root of the three species plant
2.2 不同生境中云锦杜鹃菌根侵染率的比较
对不同生境云锦杜鹃的毛根染色后,在各根段的表皮细胞内都或多或少地有典型的菌丝卷,如图2A;茶也有少量的丛枝菌根(AM)结构,如图2B;箬竹未见菌根结构。云锦杜鹃的菌根侵染率(表1)在箬竹伴生生境中极显著高于茶伴生和无下木伴生的生境(P<0.01),而在茶伴生和无下木伴生的生境之间没有差异。
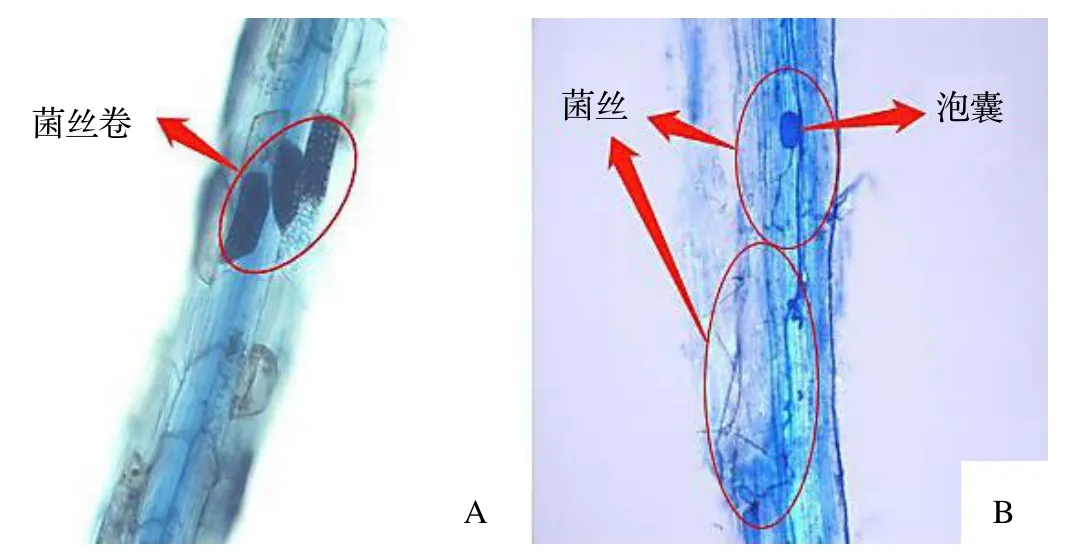
图2 云锦杜鹃毛根(A)和茶根(B)的显微结构Figure 2 Microstructure of hairroots from Rh.fortunei (A) and C.sinensis (B)
2.3 不同生境中土壤和凋落物的主要营养元素的比较
对3 种生境的土壤参数进行分析(表1),结果表明除NH4+-N 外,其他测定的参数在不同伴生种的生境间都有显著差异(P<0.05)或极显著差异(P<0.01)。从表2 可知,TC、TN、C/N和N/P 在箬竹伴生生境中都极显著高于茶伴生和无下木伴生生境中,TP 则相反(P<0.01),在箬竹伴生生境中更低,这5 个参数在茶伴生生境和无下木伴生生境中均未有显著差异。NO3--N 在无下木伴生生境中显著高于其他两个生境中(P<0.05),pH 值则是在茶伴生生境中显著高于在无下木伴生生境中(P<0.05)(表1)。总体上,通过土壤主要营养元素的比较发现,在箬竹伴生生境中土壤各参数的变化更大。
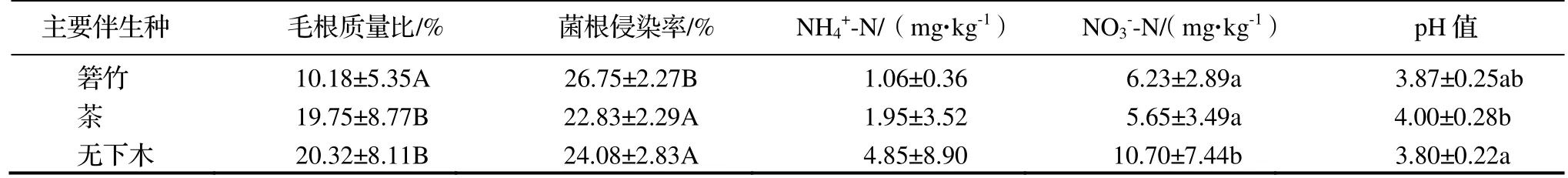
表1 3 种生境土壤无机N 和云锦杜鹃毛根质量比及菌根侵染率Table 1 Inorganic N in soil,hair root mass ratio and mycorrhizal infection rate of Rh.fortunei in 3 habitats
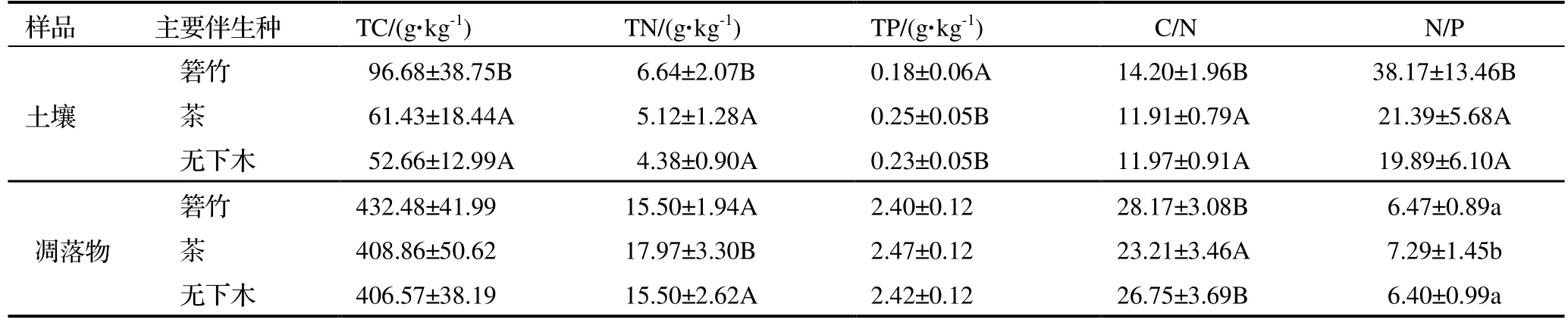
表2 3 种生境云锦杜鹃林下土壤和凋落物的主要元素和元素计量比Table 2 Major elements and their stoichiometric ratio in soil and litter under Rh.fortunei population at 3 habitats
通过对3 种生境凋落物样本的分析,结果发现TN、N/P 和C/N 在茶伴生的生境中前两个参数更高,而C/N更低,各参数在箬竹和无下木伴生的生境间都没有显著差异。将土壤和凋落物各参数与毛根质量比和菌根侵染率进行相关性(Pearson 相关系数)分析发现,土壤TC、TN、C/N 和N/P 与毛根质量比都呈显著负相关,与菌根侵染率都呈显著正相关,土壤pH 值与菌根侵染率呈显著负相关。凋落物中只有C/N 与菌根侵染率呈显著正相关关系。
3 讨论
在华顶山,调查的3 种云锦杜鹃生境各有特点,无下木伴生的生境中没有优势伴生种,仅在树冠周围有一些草本植物。人工种植的茶,根系并不发达,在空间上与云锦杜鹃保持一定的距离。箬竹的扩张占据了地上和地下的空间,阻碍了云锦杜鹃毛根的扩展和发育,导致其毛根数量急剧减少。这与1997 年建立华顶国家森林公园,停止放牧后草食性动物减少,促进了箬竹的进一步扩张有关。
2006 年和2007 年5 月,Zhang 等在华顶山、黄山、幕阜山和四明山调查的野生云锦杜鹃的菌根侵染率均在65%以上[19],本文调查的云锦杜鹃的菌根侵染率仅为19.67%~ 28.63%,菌根侵染率偏低,可能与采样时间为7—8 月的夏季高温有关,因为菌根侵染率不仅会受到不同水分状况的影响[20],还与温度呈负相关[21]。Hutton 等在澳大利亚西南部对Epacris obtusifolia毛根发育和ERM 菌根侵染的研究发现,它们在较冷湿的月份中更活跃,菌根侵染率呈现季节动态,夏季最低[22]。Read 认为在高海拔区杜鹃花科植物群落占优势与ERM 的存在有关[23],因此,随着全球气候的变暖,适应冷凉气候的很多高山杜鹃的优势可能会逐渐丧失。
本次测量的华顶山云锦杜鹃林地总N 为4.38~ 6.64 g·kg-1,远高于20 年前吴家森等测定的1.62~ 1.97 g·kg-1[24],这可能与全球的N 沉降有关。在荷兰和英国部分欧石南荒地(heathland),杜鹃花科的矮灌木欧石南Erica tetralix和帚石南Calluna vulgaris在N 沉降加剧的背景下正在被草本植物曲芒发草Deschampsia flexuosa和天蓝麦氏草Molinia caerulea所取代[25],因为在高营养水平下,草本植物的生长速率更快,而杜鹃花科植物在低营养水平下更占优势[26]。菌根真菌在根部的侵染程度和菌根生物量会随着N 沉降而减少[27]。另外,人工施肥(NH4+-N)可能会使土壤微生物多样性减少[28],有茶伴生的云锦杜鹃的菌根侵染率,很有可能也会随着施肥的加剧和时间的推移逐渐降低。在箬竹伴生的生境中,总N 水平最高,但是云锦杜鹃的菌根侵染率也最高,很可能是因为宿主需要菌根真菌在抵御胁迫和抵抗土壤病原体方面为其提供帮助[29]。因此,监测伴生种及其生境土壤的变化,对于保护野生杜鹃花科植物资源是非常必要的。
4 结论与建议
生长在华顶山的云锦杜鹃历史悠久,曾经枝繁叶茂。但是,箬竹扩张使云锦杜鹃地下部生长发生变化,其毛根的发育受阻,而菌根侵染率显著升高(P<0.01)。箬竹伴生生境的土壤中的总C、总N、总P、C/N 和N/P均与在茶伴生和无下木伴生的生境中有极显著的差异(P<0.01)。箬竹扩张不仅能侵占云锦杜鹃毛根的发育空间,还改变着土壤中营养元素的资源比率,长期作用对云锦杜鹃植株的生长不利。因此,应该采取人为措施,铲除云锦杜鹃周围的箬竹,包括地上和地下两部分,结合人工促进云锦杜鹃幼苗的天然更新,确保华顶山云锦杜鹃种群的健康和可持续发展。
