松属植物地质历史及生物地理演变
2022-07-14梁莉娜李相传
胡 祎,梁莉娜,肖 良, ,李相传, *
1.长安大学 地球科学与资源学院,西安 710054
2.中国科学院南京地质古生物研究所 现代古生物学与地层学国家重点实验室,南京 210008
3.西北大学 陕西省早期生命与环境重点实验室,西安 710069
松属是现存裸子植物最大的属,约113 种,占松科的50%,广泛分布于北半球(Richardson,1998;Fu and Mill,1999),该属是组成北温带、北亚热带高山针叶林及干旱林地的主要成分(李楠,1995;Gernandt et al,2005),仅有Pinus merkusii Jungh. & de Virese 一种生长在赤道以南的印度尼西亚苏门答腊岛(Mirov,1967)。由于具有悠久的演化历史,松属类群成为研究针叶类植物进化以及物种分化的经典材料(Willyard et al,2007;Liu et al,2014;Zeb et al,2020;Jin et al,2021)。同时松属又是研究植物传播机制演化的模式系统,主要有两类种子传播机制:约75 种通过风力传播,24 种因种翅退化或发育不全由脊椎动物传播,剩下10 余种两种传播方式均存在(Salazar-Tortosa et al,2019)。松属物种具有非常重要的生态及经济意义,是北温带森林的建群树种,也是重要的造林、绿化树种及木材、纸浆、坚果、松脂、家具等工业加工的材料来源(郑万钧和傅立国,1978)。同时,部分种类具有适应森林野火的生态特征(He et al,2012;Keeley,2012;Pausas,2015;Gallien et al,2016;Badik et al,2018;Singh et al,2021),是生态演替中森林火灾后的先锋物种。松针、球果以及松树皮中含有丰富的化学成分,如萜类、类黄酮、糖类以及木脂素等(刘敏莉等,2011;韩松和张文治,2018;Sharma et al,2018),这些成分具有重要的药用价值。这些特点使该属植物成为植物学、生态学和分子系统发育学等的研究热点。
形态上,松属通常具有(1-)2-5(-8)针每束的针叶,着生于短枝鳞叶叶腋内;雌球果两年成熟,种鳞具有显著的鳞脐和鳞盾,分别对应于第一、二生长季种鳞露出部分(Farjon,2005),这些特征将松属与松科其他现代属相区分。分类学上,作为松科中松亚科的唯一类群,其属下划分前人提出40 多个分类方案(Richardson,1998)。Little and Critchfield(1969)在Shaw(1914)的工作基础上,将松属划分为3 亚属、5 组、15 亚组及94种。这为后续划分方案确定了基本框架。后来,许多学者对上述分类体系的亚属、亚组的分类和种的数量进行了修正(Richardson,1998;Gernandt et al,2003;Gernandt et al,2005;Jin et al,2021)。需要指出的是,Gernandt et al(2005)基于101 种松属植物叶绿体基因测序结合细胞核DNA 及形态学证据将松属划分为2 亚属、4组、11 亚组和110 种(表1)。该分类系统在近年被广泛接受(Saladin et al,2017;Zeb et al,2020;Singh et al,2021)。Parks et al(2012)分析了107 种松属的叶绿体基因组,很大程度上证实了Gernandt et al(2005)提出的分类结构的合 理 性。Eckert and Hall(2006) 和Saladin et al(2017)基于化石校准点的系统发育学研究认为松属的冠龄为早白垩世和最早的化石记录时间(Ryberg et al,2012) 相一致,比Willyard et al(2007)、Gernandt et al(2008)、Leslie et al(2012)以及Hao et al(2015)研究得出的年龄早约30 Ma。Jin et al(2021)基于遗传序列及4 个化石校准点的最新研究推测松属的茎龄为155.3 Ma,即晚侏罗世(钦莫利期),这一结论比已发现的松属化石记录略早。两个亚属的分化时间为98.77 Ma,为晚白垩世早期(塞诺曼期),与两个亚属的化石记录较为一致。

表1 松属的现代分类体系(改自Gernandt et al(2005))Tab. 1 Classification of Pinus (modified from Gernandt et al (2005))
松属植物在北半球白垩纪和新生代地层中的化石记录十分丰富,主要为球果、带翅种子、针叶以及木材等大化石,是当时北半球森林的主要组成部分。Axelrod(1986)描述了美国西部松属的新生代历史,分别描述了该属亚组下的化石记录,认为该属物种多样性及迁移扩散和火山活动、构造作用及气候波动相关;Mai(1986)对欧洲古近纪和新近纪的松属球果进行总结且最终将其归为40 多个化石种,这些物种中的多数与现代物种特征不同,属于欧洲古特有种;李楠(1995)、陆素娟和李乡旺(1999)对松属的化石历史以及演化进行了讨论;李楠(1995)认为白垩纪中、晚期松属属内发生一定程度的分化且劳俄古陆可能为该属最早分化的场所,陆素娟和李乡旺(1999)根据松属最早的孢粉化石记录认为其在康滇古陆于三叠纪起源;Richardson(1998)对松属植物早期演化(主要针对始新世)及化石记录进行了综述,发现将板块构造、气候、化石和松属生物地理学相结合时,始新世成为该属演化的重要阶段之一。近年来,随着松属的古植物学和分子系统学研究深入,其起源、演化及扩散等问题可能会有新的见解。可靠化石的不断发现对认识松属物种起源及地史演化有十分重要的作用,同时也为各种分子假说提供直接证据,因此对松属化石地质历史的梳理及研究变得尤为重要。本文总结了全球松属的大化石记录,并绘制了世界范围内松属的现代和地史分布图,讨论了该属的化石历史并对松属的起源时间、地点以及扩散路线提出了新的观点。
1 现生松属的分布
了解现代松属的地理分布可以为更好地研究松属的地质历史提供理论基础。目前全球松属共计113 种(Farjon,2017),属于北半球温带分布类型,自然分布在北美洲、中美洲、欧洲以及亚洲地区(表1, 图1), 其中仅2 种(P. pinaster Aiton 和P. canariensis C. Sm.)延伸到非洲北部,南半球也有许多引种栽培的松属物种(Gallien et al,2016),如P. caribaea Morelet、P. pinaster、P. radiata D. Don 以及 P. patula Schiede ex Schltdl. & Cham(Le Maitre,1998)。松属在松科中的地理分布范围最广,和松科的整个分布范围几乎一致(Farjon and Filer,2013),松属的多样性分布中心为北美洲(约70 种,特别是墨西哥、加利福尼亚)以及东亚(约25 种,在中国尤为集中)(Richardson,1998)。

图1 现生松属的分布(改自Farjon(2005),Farjon and Filer(2013))Fig. 1 Distribution of extant Pinus (redrawn from Farjon (2005), Farjon and Filer (2013))
松属的垂直分布范围从海平面延伸到4000 m(Farjon,2017),其中约58 种分布在海拔2500 m以下的地区;松属垂直分布较松科的其他属低,一般仅分布在3000 m 以下(李楠,1995),只有个别种的最高海拔达到4000 m,如分布于墨西哥与危地马拉的P. hartwegii Lindl.和分布在中国青海南部、四川西部、云南西北部、西藏东部的P. densata Mast.;分布于美国阿拉巴马州和佛罗里达州的P. clausa (Chapm. ex Engelm.) Vasey ex sarg.海拔最低,仅在5 — 90 m。松属植物中还有一些物种适应性较广,如分布于中国及缅甸的P. armandii Franch.在海拔0 — 3800 m 均可生存。
松属物种的垂直分布幅度受地形条件和纬度气候带制约。同种松属物种在不同地区的海拔分布范围有所差异,如分布在北美洲西部落基山脉、喀斯喀特山脉和内华达山脉的P. albicaulis Engelm.,在其分布范围的北部海拔为1350 m,而在南部海拔高达3650 m;油松P. tabuliformis Carrière 在中国东北部及中部海拔位于100 — 2600 m,在四川的海拔高达3800 m;再如红松P. koraiensis Siebold &Zucc.在俄罗斯远东地区海拔仅200 — 600 m,在中国海拔为500 — 1300 m,而在日本海拔又高达2500 m。综上所述,一般情况下分布在北部及东部的物种海拔较低,植物垂直分布结构可能和生态环境如气候、海拔、土壤变异性以及对火的适应性等相关。
虽然对松属现生分布的研究具有指导意义,但要了解松属的系统发育关系及植物地理历史,更多地要依靠化石记录的证据。
2 松属化石的种类及分布
双束松粉属(Pinuspollenites)是一类被认为和现生松属亲缘关系密切的花粉化石属。Li et al(2017)报道了中国四川盆地晚三叠世的双束松粉属Pinuspollenites。虽然松属花粉(具双气囊)在晚中生代和古近纪的沉积物中较常见,但它们几乎没有展示能反映进化状态的形态性状且只能鉴定到属一级,而大化石可以鉴定到亚属、组、亚组甚至种一级。因此本文在讨论地史分布时只采用松属的可靠大化石记录。
松属大化石记录十分丰富,在美国东北部、东部、北部以及南部,日本、中国、俄罗斯的勘察加半岛、比利时、英国及捷克等地的白垩纪地层中发现了约27 种(3 个未定种)(Alvin,1960;杨家驹等,1996;Meijer,2000;Kvaček,2013),包括球果、种子、叶子及木材(图2)。如图2b 所示,该时期松属明显的分布中心在北美洲东部、西欧及日本。需要指出的是,Falcon-Lang et al(2016)报道了加拿大早白垩世(凡兰今期,133 — 140 Ma)的木化石P. mundayi,认为它是松属最早的化石记录;但Hilton et al(2016)对其分类学提出了质疑,不同意Falcon-Lang et al(2016)展示的解剖结构是松属植物的特征。
这样,英国约克郡下白垩统(131 — 129 Ma)中发现的球果P. yorkshirensisRyberg,Stockey, Hilton,Mapes,Riding et Rothwell 是目前世界上最早的关于松属的可靠记录(Ryberg et al,2012)。另外一例早期记录是发现于比利时早白垩世的P. belgicaAlvin(Alvin,1960)。因此,化石记录表明松属起源时间可能为早白垩世或更早。白垩纪时期松属的两个亚属可能已经分化且在类群数量上相差悬殊;到目前为止,仅有发现于美国晚白垩世的针叶Pinussp.(Jeあrey,1908)和球果P. magothensis(Penny,1947)以及日本的针叶P. yezoensis(Stopes and Kershaw,1910)与单维管束松亚属(SubgenusStrobus)有亲缘关系(尽管 Miller and Malinky(1986)和Willyard et al(2007)对此持有异议),其余均属于双维管束松亚属(SubgenusPinus)。Meijer(2000)报道的比利时东北部晚白垩世地层中的木化石Pinuxylonsp.是该时期确定的单维管束松亚属化石记录。综上可知白垩纪时期双维管束松亚属可能在当时的生态系统中占主导位置且更接近祖先类群。
古新统松属化石记录非常少,到目前为止仅有Dolezych and Reinhardt(2020)报道于加拿大努勒维特(Nunavut)的松属木化石Pinuxylon selmeierianumDolezych & Reinhardt,其解剖特征将该种归为单维管束松亚属。
始新世以来,松属是北半球各植物群中重要且较常见的组成部分(Ferguson,1967;Miller,1973;Stockey,1983;Axelrod,1986;Mai,1986;Richardson,1998;Eckert and Hall,2006)。始新世松属的数量较古新世明显增多(图3),主要分布地为美国的华盛顿州(Miller,1974)、科罗拉多州、加利福尼亚州、怀俄明州及内华达州(Axelrod,1986),加拿大的不列颠哥伦比亚(Stockey,1984),日本(Huzioka and Takahashi,1970),欧洲(Mai,1986)以及中国辽宁(《中国新生代植物》编写组,1978)和广东(Xu et al,2015a),该时期的分布中心在北美洲西部(图3)。单维管束松亚属的记录也有所增加,如在中国广东发现了晚始新世地层中的球果P. maomingensis与针叶Pinussp.(Xu et al,2015a),这是单维管束松亚属在中国的最早化石记录。该时期有一个值得注意的化石记录,Klymiuk et al(2011)报道的中始新世基于整株植物的P. arnoldiiMiller.,这对研究植物进化趋势与解决主要分支系统发育的总体格局有重要意义。

图2 松属白垩纪全球地理分布Fig. 2 Cretaceous distribution of global records of Pinus
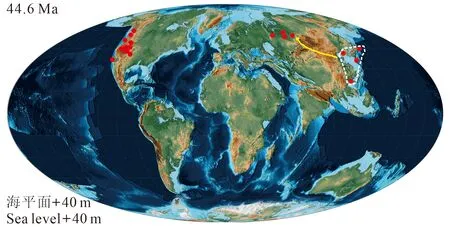
图3 松属始新世全球地理分布Fig. 3 Eocene distribution of global records of Pinus
渐新世松属植物化石记录也较多(图4),主要为球果和针叶化石,来自北美洲的蒙大拿州(Miller,1969)、华盛顿州(Underwood and Miller,1980)、不列颠哥伦比亚省(Banks et al,1981)、爱达荷州(Axelrod,1986)及科罗拉多州(Axelrod,1986;Wolfe and Schorn,1990),亚洲的外高加索东部与西部(Millar,1993)、中国吉林(Guo and Zhang,2002)和云南(Zhang et al,2021)。在日本,松属是一些植物群的次要组成部分(Graham,1972)。在特定植物区系中大量的松属物种可以共存,如德国劳恩贝格(Rauenberg)植物群中记录了6 个针叶化石的 未 定 种、1 个 新 种P. engelhardtiiMenzel 和1个相似种Pinus(Pinus) cf.thomasiana(Goeppert) Reichenbach(Kovar-Eder,2016)。晚渐新世—早中新世,Grote and Srisuk(2021)报道了泰国的3个未定种、1 个相似种Pinuscf.krempfiiLecomte及1 个新种P. nongyaplongensisGrote,这些化石种表明在泰国晚渐新世—早中新世,该属的多样性比现在丰富。

图4 松属渐新世全球地理分布(古地理图据Scotese(2014d))Fig. 4 Oligocene distribution of global records of Pinus (after Scotese (2014d))
中新世全球松属分布更加广泛数量急剧增加,达到地质历史上的繁盛时期(图5),分布类型基本和现生分布格局相近即北半球广泛分布(图1,图6),如欧洲的奥地利(Klaus,1989)、法国(Kvaček et al,2011)、捷克(Teodoridis and Sakala,2008)、德国(Kvaček et al,2014)、希腊(Zidianakis et al,2016)和波兰(Kowalski,2017);美国俄勒冈州、爱达荷州、马萨诸塞州、加利福尼亚州、华盛顿州、内华达州、马里兰州(Miller,1978;Axelrod,1986;Tidwell et al,1986;Miller,1992);日本部分地区(Miki,1939;Miki,1957;Tanai,1961;Tanai and Onoe,1961;Choi et al,2010;Jeong et al,2012;Yamada et al,2015)、中国河北(Li et al,2021)。还有一些种扩散至北亚热带以及热带地区,如中国浙江、云南(陶君容等,1994;Luo et al,2009;Xing et al,2010;Ding et al,2013;Xu et al,2015b;Zhang et al,2015;Wang et al,2017;An et al,2019)和 印 度(Khan and Bera,2017)。该时期中国只有发现于云南的P. uniseriataH. B. Wang, A. A. Oskolski et Z. K. Zhou 属于单维管束松亚属(Wang et al,2017)。Tanai and Onoe(1961)在日本北海道发现的早中新世化石P. palaeopentaphyllaTanai et Onoe,Guo and Zhang(2002)在中国吉林也发现了渐新世P. palaeopentaphylla的针叶化石,这表明该种在东亚生存了很长一段时间。
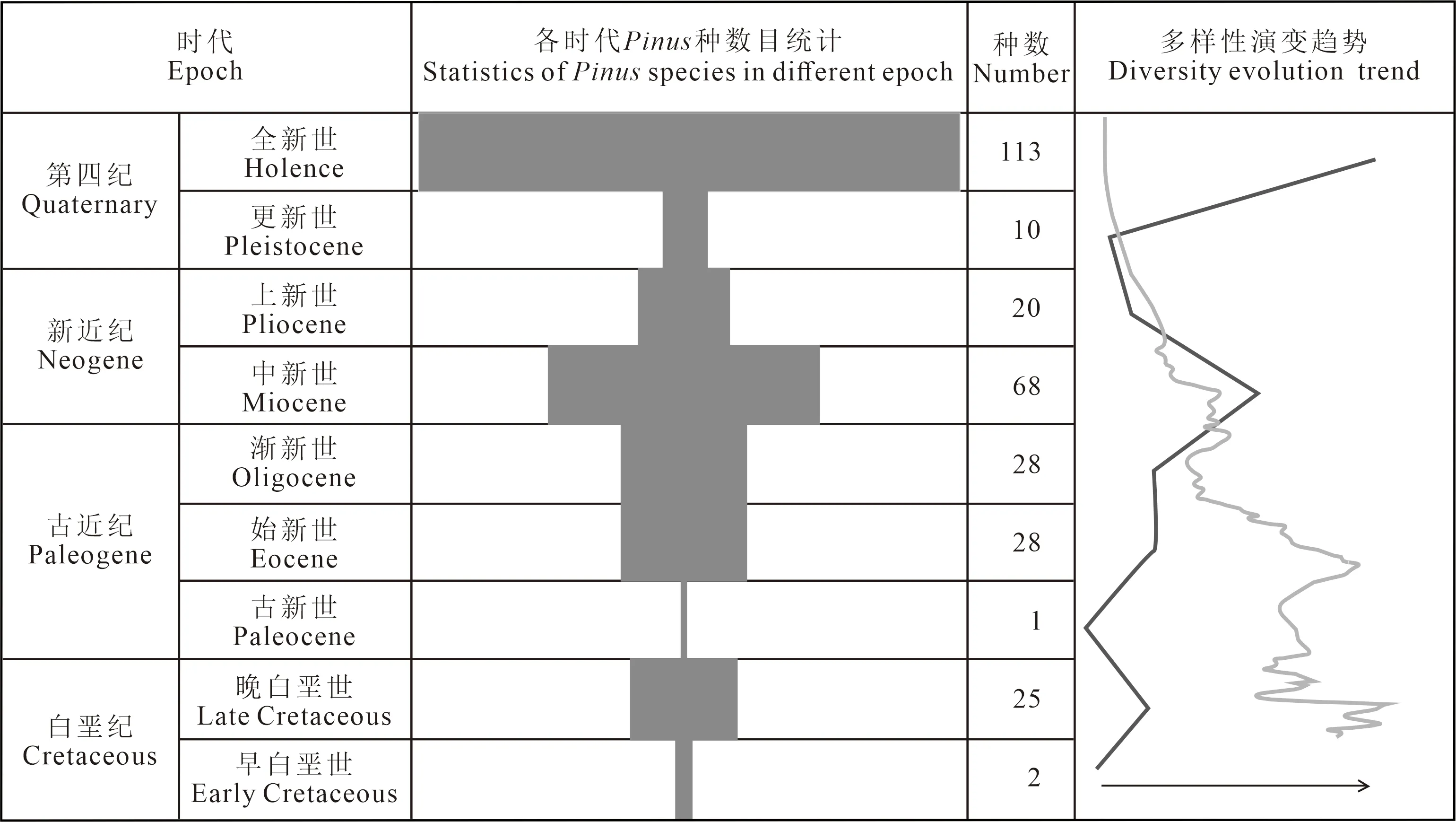
图5 松属多样性地史演化Fig. 5 Diversity evolution of Pinus over geological past

图6 松属中新世全球地理分布(古地理图据Scotese(2014d))Fig. 6 Miocene distribution of global records of Pinus (after Scotese (2014d))
上新世松属在欧洲、北美洲以及东亚均有分布(图7)。如中国云南发现的2 个现生种P. yunnanensis 和P. armandii(陶君容和孔昭宸,1973;扆铁梅等,2002;Zhao et al,2008);McKown et al(2002)发现于加拿大的P. matthewsii McKown Stockey et Schweger;Stults et al(2010)在美国Alabama发现的叶化石 ;Mai and Velitzelos(2007)报道的希腊上新世 — 更新世的球果化石P. palaeostrobus (Ett.) Heer。而到了更新世,松属化石记录减少,可能和成岩较差有关,主要分布在阿尔巴尼亚(Denk et al,2019)、加利福尼亚州(Axelrod,1983;Axelrod,1986;Axelrod and Cota,1993)、日本(Miki,1957)等地区(图7)。

图7 松属上新世 — 更新世全球地理分布Fig. 7 Plio — Pleistocene distribution of global records of Pinus
综上所述的化石证据表明:松属化石和现代种的分布范围相近,但有些许差别。如加拿大的班克斯岛发现化石种(Williams et al,2008),但无现生种分布。从白垩纪到现在,松属物种多样性整体表现为增加的趋势(图5),这在一定程度上反映出该类植物能够适应逐渐变冷的气候,而在古新世时期的化石发现较少,可能受到当时气候及白垩纪末期生物大灭绝的影响,松属植物正在复苏,上新世至更新世气候变冷,松属物种的分布范围有所缩小,当然,也可能是由于该时期采样程度的欠缺所导致的。
3 松属的地史演变
对松属的起源问题,Richardson(1998)和Millar(1993)提出其进化的3 步假设:松属种群中生代广泛分布于中纬度地区;始新世时期种群碎片化,迁移到高纬度、低纬度地区及中纬度高地;渐新世和中新世时期又广泛拓殖到许多中纬度地区。根据全球松属化石记录、现生分布及分子系统发育等研究,李楠(1995)、Eckert and Hall(2006)均支持Richardson(1998)、Millar(1993)的假说。
目前,已知松属可靠的最早化石记录来自比利时和英格兰约克郡下白垩统(Alvin,1960;Ryberg et al,2012),这说明松属早白垩世已经存在于西欧;到晚白垩世,松属植物可能已经在其他地区出现类群分化,这通过发现于加拿大新斯科舍,美国新泽西州、特拉华州、马萨诸塞州、纽约、密西西比州、明尼苏达州和堪萨斯州以及日本北海道和中国河南等多地上白垩统(图2b)的化石记录所证实,说明这类针叶植物在中生代末就已经开始在北半球广泛分布。从目前已知白垩纪化石记录分布来看(图2),其迁移路线从可能的起源地如西欧通过北大西洋陆桥在晚白垩世之前向西传播至北美洲东部;至于欧亚大陆之间此时区系交流应该存在一定的地理障碍(Tiffney and Manchester,2001),因为白垩纪时期Turgai Strait 仍然是一片海域(图2),而古海陆分布格局表明早白垩世亚洲和北美洲之间应有陆地连接(图2),同时北美洲陆中海道尚未形成(李楠,1995),故东亚松属早期分子(中国、日本等)很可能是从北美洲西部经过白令陆桥散布的。Langenheim et al(1960)报道的在北美洲西部阿拉斯加地区的早白垩世松属孢粉化石是这一传播路径很好的佐证。
白垩纪时期丰富的木炭化石记录(Scott,2000;Glasspool and Scott,2010)以及大气中氧浓度的明显增加(Belcher et al,2010;Glasspool and Scott,2010)表明该时期有一个易燃的环境即火灾发生相对频繁(Bond and Scott,2010)。晚白垩世松属分化为单维管束松亚属和双维管松亚属两个亚属,前者相较于后者栖息在更干燥或寒冷的环境中(Salazar-Tortosa et al,2020)且属于规避火灾的类型。两个亚属对不同环境的选择可能是导致其分化的原因,如对火的适应性(Badik et al,2018;Singh et al,2018;Jin et al,2021)。证明在这样一个火灾易发的时期,双维管束松亚属物种由于适应火灾生境而其数量占据优势,晚白垩世仅有比利时东北部的1 种单维管束松亚属Pinuxylonsp.被发现(Meijer,2000),也证明了以上推测。
虽然松属物种对气候和土壤有广泛的耐受性,但它们不适应高温、潮湿的热带环境(Mirov,1967)。新生代早期出现显著的增温趋势(图5),早古新世出现明显的温度和湿度增加,在早始新世气候最适期(约50 — 52 Ma)达到高峰(Richardson,1998;Zachos et al,2001)。在这种更加温暖潮湿的气候背景下,松属物种的分布被限制在高纬度地区,形成不利于该属类群生长的条件,因此古新世地层中松属化石记录极其稀少,目前仅在加拿大古新统中发现了木化石Pinuxylon selmeierianum(Dolezych and Reinhardt,2020),其分布地属于高纬度地区(82°N)。
古新世/始新世极热事件(PETM)使得早始新世全球气候短暂且强烈地变热(Zachos et al,2001;Zachos et al,2006;McInerney and Wing,2011,图5),此事件对陆地和海洋生态系统也产生了显著的影响(Smith et al,2007;McInerney and Wing,2011)。始新世时期,生态群落在多样性和丰富度上均以能够适应干旱的热带和亚热带环境的豆科植物为主(Wing et al,2005;Smith et al,2007),普遍缺乏针叶树成分。这从始新世早期较少的松属大化石记录得到印证(Axelrod,1986)。中始新世暖湿期,松属大化石发现于内华达州的中纬度高地地区(Axelrod,1966,1968),说明在温暖潮湿条件下,松属收缩到有利于它们生存的避难区,这与Richardson(1998)提出始新世松属种群分布在中纬度高地地区相符;此时,Turgai Strait尚未闭合,有效阻止了动植物类群从东亚向西扩散到欧洲(LePage and Basinger,1995),故晚始新世分布于西伯利亚的化石种(Dorofeyev,1963)可能是东亚中、低纬度地区的松属类群(Huzioka and Takahashi,1970;Graham,1972;《中国新生代植物》编写组,1978;Xu et al,2015a)直接散布的。连接北美洲和欧洲洲际传播的DeGeer 路线贯穿整个始新世,因此该时期松属类群在北美洲和欧洲之间继续传播。
始新世 — 渐新世过渡时期,南极大陆冰盖形成且迅速扩张(Zachos et al,2001;Williams et al,2007),发生了显著的早渐新世降温事件。导致晚始新世许多中纬度地区的北方热带植物区系被子植物喜热分子大量灭绝(Millar,1993),而松属植物占据了中纬度地区大量的生态位(图4),这与Richardson(1998)和Millar(1993)认为渐新世松属拓殖到中纬度地区符合。该时期北大西洋陆桥因大洋中脊持续扩张遭到破坏,欧洲和北美洲之间松属类群的交流在此时终止;而Turgai Strait 在此时干涸(LePage,2003),松属种群在欧洲和亚洲之间开始扩散。
中新世时期,构造活动加剧,青藏高原整体隆升,导致了亚洲内陆干旱化以及形成了东南峡谷山脉相间的多样化生境(An et al,2001;Guo and Zhang,2008;柯学等,2013)。该时期松属植物得到了适应辐射,广泛分布于北半球不同纬度地区,如热带泰国(Grote and Srisuk,2021)、亚热带浙江(Ding et al,2013)以及北温带日本(Jeong et al,2012)。Jin et al(2021)通过对松属现生分布以及31 个环境变量关系的研究认为松属物种的丰度和最潮湿季度的降水成反比,并指出中新世以来干旱对松属多样性有促进作用。该时期的大多数松属物种均分布在中纬度地区(图6),这与Richardson(1998)认为中新世时期该属又广泛拓殖到许多中纬度地区的观点相一致。许多现代松属的直接祖先可以追溯到中新世(Choi et al,2010;Xing et al,2010;Ding et al,2013;Yamada et al,2014;Xu et al,2015b;Li et al,2021),甚至在该时期发现的个别化石种属于现生类群,如Klaus(1989)在奥地利发现的P. halepensisMill.及Zhang et al(2015)在中国云南发现的马尾松P. massonianaLambert.。这些化石记录和Jin et al(2021)通过基因测序研究得出90%现生松属类群都是在中新世通过再分化而产生的结论保持一致。
4 总结与展望
松属化石证据表明:松属早白垩世起源于西欧,在晚白垩世之前通过北大西洋陆桥扩散到北美洲,从北美洲西部经过古白令陆桥传播到东亚;晚白垩世分化出两个亚属(SubgenusPinus和SubgenusStrobus),且该时期松属已经开始在北半球中纬度地区广泛分布。全球古气候、地质构造等的变化以及火灾机制等都有利于松属类群物种多样性的产生,促进松属植物的进化。然而,深刻认识松属植物演化和地理迁移历史,仍然需要大量的古植物学证据,尤其是关键时期(如古新世)。
致谢:感谢审稿专家对稿件修改提出的宝贵意见。
