天山北坡中段山地紫花鸢尾草甸植物群落动态分析
2022-07-13张鲜花李江艳吴咏梅朱进忠
张鲜花,李江艳,吴咏梅,朱进忠
(新疆农业大学草业与环境科学学院/新疆草地资源与生态实验室,乌鲁木齐 830052)
0 引 言
【研究意义】近几十年来,草地有不同程度的退化[1-2],一个群落及其组成种群特征的消长变化,预示着草地的演替趋向[3-4],对其进行动态研究,可以了解当前草地数量与质量及其发展趋势,在一定时间尺度下的变化分析,揭示某一草地区自然环境变化和人类活动干扰的程度[5-8]。杨晓玫等[9]对天祝不同退化程度草地植物群落结构与物种多样性研究,认为草地退化程度的增加,其植物多样性逐渐降低,植物群落结构分布单一;戴黎聪等[10]通过2008~2016年定位监测数据,分析了青藏高原高寒小嵩草草甸和高寒金露梅灌丛两种植被净初级生产力年际动态,并探讨了气候因子对其影响,认为在全球气候变暖的背景下,对2种草地地上生物量的影响是不同的。【前人研究进展】紫花鸢尾草甸草地是天山北坡中段山地由典型的适中温、中湿条件的植物组成草地类型,与雪岭云杉(Piceaschrenkiana)群落复合分布,是天山北坡中西段山地草甸植被中分布较为广泛的一种类型[11-12]。草地以紫花鸢尾为优势种,与多种杂类草构成群落建群层片,草群茂密,形成能量与物质聚集的最大地段,是重要的山地牧场。紫花鸢尾是鸢尾科(Iridaceae)、鸢尾属(Iris)植物,再生性强、耐践踏,具有极强的侵占性,在当草地利用过度时,仍具有较强恢复生长与繁殖能力,抑制其它植物生长,使草地质量降低,在紫花鸢尾聚集性较大的地段,常被认为是草地退化的表现[13]。【本研究切入点】以紫花鸢尾草甸草地为对象,分析1985~2004年近20年定位连续测定各项群落学指标,了解这一阶段草地植被动态变化特点与趋势。【拟解决的关键问题】以优势种紫花鸢尾和主要伴生种草原老鹳草、草地早熟禾和异燕麦为对象,采用连续定位测定方法,分析1985~2004年间群落的植物种类组成、数量特征,研究天山北坡中段山地紫花鸢尾草甸植物群落动态分析。
1 材料与方法
1.1 材 料
研究区位于天山北坡中段,距乌鲁木齐市西南80 km南山种羊场谢家沟地区,地处87°25′~87°33′E, 43°07′~43°47′N,海拔1 900~2 400 m。年降水量450~560 mm,一般年份雨季6月下旬至8月上旬,有时出现春旱和秋旱;年平均气温1.2~2.8℃;土壤为山地黑钙土。草地与森林呈复合分布,由杂类草、中生禾草与灌丛组成不同群落,占据山地半阴坡、台地和林间空地。草地春秋两季放牧利用,放牧时间春季3月中旬至6月下旬,秋季从9月上旬至11月下旬,共160 d左右。
以优势种紫花鸢尾和主要伴生种草原老鹳草、草地早熟禾和异燕麦为对象,分析群落植物种类组成的变化情况。
1.2 方 法
在自然放牧利用状态下,采用区域定位观察的样地法,对植物群落的种类组成、高度、盖度和地上生物产量进行长时段连续测定。测定时间段为1985~2004年,每年于相同地点、相同季节进行取样。
1.2.1 取 样
以草地型为单位采取典型地段取样法进行。用于群落特征分析与记载样地面积为10 m×10 m,地上生物产量测定样方面积为1 m×1 m,每年调查样方不少于30个,调查时间为每年7月10~15日植物生长旺季进行。
1.2.2 测定指标
群落盖度:采用目视法估测。测定群落投影总盖度和种群投影分盖度。
地上生物量:采用样地实测法测定群落植物种和类群地上生物产量。
植物优势度:采用综合优势度(SDR)计算。SDR=(C+H+W)/3.
式中,C、H、W分别为草地群落组成植物的相对盖度、相对高度和相对地上生物量。
2 结果与分析
2.1 群落主要植物种类组成变化
研究表明,在1985~2004年时段内,草地植物群落组成优势种和主要伴生种是稳定的,以杂类草与禾草层片构成的群落格局没有发生改变。杂类草成片中紫花鸢尾、草原老鹳草出现率达100%,禾草层片中以草地早熟禾出现率达93.33%,异燕麦为80%。紫花鸢尾、草原老鹳草、草地早熟禾和异燕麦始终是构成群落组成的基本成分,决定着群落外貌、结构和功能。建群种紫花鸢尾草地群落虽然长期处于放牧干扰之下,但目前干扰强度无论是自然的还是人为的,还不足以引发群落种类发生演替。表1

表1 紫花鸢尾草甸群落主要种群年际动态Table 1 Dynamic changes of major population in Iris ruthenica Ker.-Gawl. Meadow
2.2 群落数量特征变化
2.2.1 植物种群高度变化
研究表明,草地群落主要植物种群高度在近20年间的时段内,始终处于波动状态,有时在年度间的波动幅度还非常大,如紫花鸢尾种群,在1987与1988年之间,1994年与相邻的1992~1995年,株高可相差近1倍;其他年份之间或高或低也存在一定的差别,整个时段内最高与最低年可相差2倍之多;草原老鹳草、草地早熟禾种群高度在年际之间差别也存在类似情况,但这种变化均表现出具有一定的可逆性,从而使群落高度始终处于波动状态。图1
利用埋地管道运输石油、石油产品及常温下流体状的各类化工产品已逐渐成为最经济且对环境破坏最小的运输方式。管道运输多为有压力运输,而长距离输送过程中埋地管道难免会跨越活动断层,震例显示约70%的管线损伤发生在断层区[1],因此,研究内压对跨断层埋地输液管道的抗震性能很有必要。对此问题的研究目前有3种分析方法:理论解析法、数值模拟法和试验研究法[2]。数值模拟是利用有限元软件设计出符合实际的模型,以此来描述管道在断层区域复杂的力学响应,其优点是计算精度高,被要求在跨断层管道抗震设计中使用。

图1 紫花鸢尾与主要伴生种植株高度变化Fig.1 The changes in height of Iris ruthenica Ker.-Gawl. and main associated plants
2.2.2 植物群落与种群盖度变化
2.2.2.1 群落的总盖度变化
研究表明,紫花鸢尾植物群落总盖度在近20年来无明显变化,没有出现随着时间变化,或者是渐进的增大或者是渐进的变小。但在不同年际间有小幅度波动,而这种波动在很小的时间尺度内就会复原。草地盖度出现了2个低峰值,1988年出现1个高峰值,与其相邻年之间达到14.6%~20.2%的差值,但在很短的时间内就恢复到了正常值范围。分布于湿润地段的草地,在诸多植物群落数量特征中,盖度与高度、地上生物产量相比具有较高的稳定性,在没有强烈干扰下,一般是不会出现较大幅度变化。图2

图2 紫花鸢尾群落总盖度变化Fig.2 The changes in total coverage of Iris ruthenica Ker.-Gawl.communities
2.2.2.2 群落种群盖度变化
研究表明,紫花鸢尾的盖度,在1985~1998年间,除了1991和1992年其盖度在50%以上,其他年份基本上是在30%~50%波动,1999年之后种群在群落中的盖度开始持续增大,均保持在50%~60%。其他3种植物种群盖度除了草原老鹳草在1987和1988年达到20%以上,其他年份虽有波动,但波幅不大。图3

图3 紫花鸢尾与主要伴生种盖度变化Fig.3 The changes in coverage of Iris ruthenica Ker.-Gawl. and main associated plants
2.2.3 群落与种群地上生物量变化
2.2.3.1 群落地上生物量变化
研究表明,草地地上生物产量在近20年的时间尺度上,一直处于1个波动变化状态,有些年间产量可相差2.2倍。从整个变化规律来看,1986~1991年,草地生物量处于1个较低阶段,1992~1998年期间草地产量持续增高,1995年达到796.3 g/m2;2000年草地产量出现大幅度下降,与1986、1987年形成在20年间草地生产力最低年份。2001年草地生产力陡然升高,从2000年的425.4 g/m2,升高到749.2 g/m2,2003、2004年又开始下降。图4

图4 紫花鸢尾草甸群落地上生物量变化Fig.4 The changes in total above ground biomass of Iris ruthenica Ker.-Gawl.meadow
2.2.3.2 群落种群地上生物量变化
研究表明,1985~1991年间,其紫花鸢尾的地上生物量,在241.3~372.3 g/m2波动,1992~1998年开始持续增长,最大年份达到548.3 g/m2;1999、2000年又出现较大幅度下降,之后又持续升高。草地早熟禾属于优良牧草,在草地中是家畜首选采饲对象,经过放牧对其生长影响较大。从种群地上生物产量历年的变化,在2000年之后,除2001年较高外,其余年份产量均有所下降。其他2类,地上生物量基本保持着在群落中固有的状态,虽有波动,但波幅不大。同时也说明,紫花鸢尾生物量的变化已成为控制草地产量的决定性因素。图5
图5 紫花鸢尾与主要伴生种地上生物量变化Fig.5 The changes in above ground biomass of Iris ruthenica Ker.-Gawl. and main associated plants
2.2.4 群落组成种群优势度变化
研究表明,紫花鸢尾在草地群落中一直保持着绝对优势地位,并随着时间的推移种群优势度呈持续增长趋势,紫花鸢尾在草地群落中随着年限的延续,种群在群落中的个体数量、盖度、生物产量是逐年增加的,基本成为群落的主宰者。其他草类优势度基本保持了一个稳态发展态势,年际间虽有变化,但变化幅度均不大。图6

图6 紫花鸢尾群落植物种类组成优势度变化Fig.6 The changes in dominance of Iris ruthenica Ker.-Gawl.meadow
2.3 降水量的波动对草地消长变化影响
研究表明,降水年度间不平衡与草地群落的波动规律基本一致,降水欠年一般也是草地群落各项数量指标的低值年,降水丰年草地地上生物产量也随之出现相应增加。降水的季节分配,多雨季节气温偏低也直接会影响到当年草地各项数量特征的高低。图7
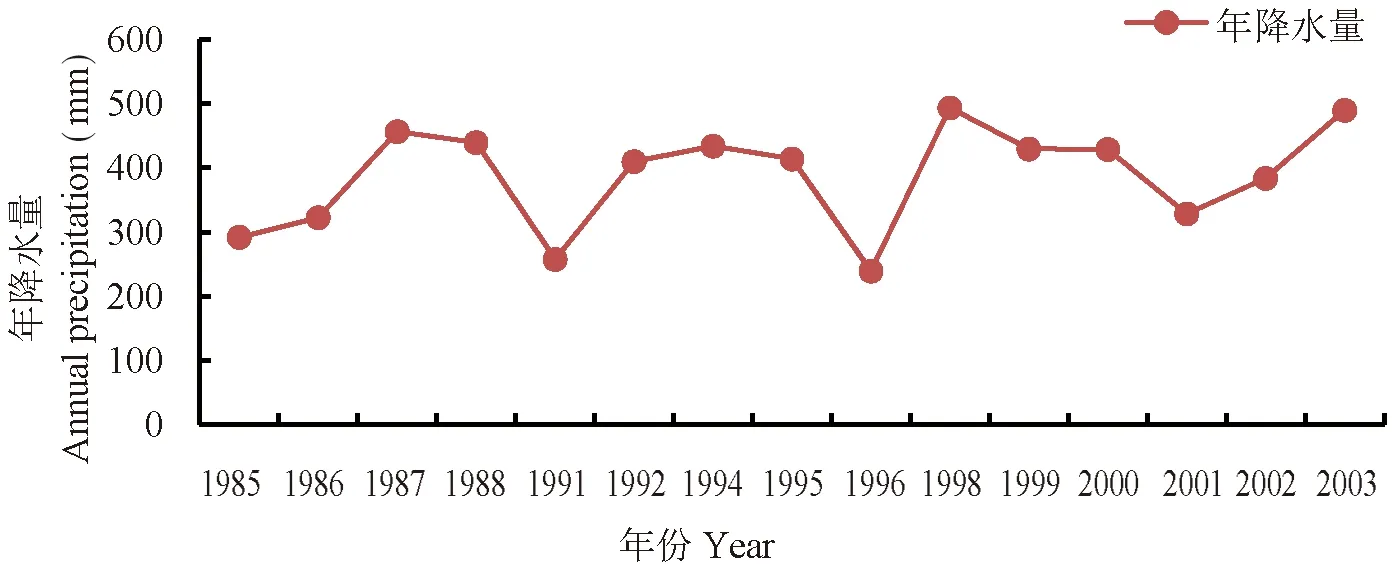
注:数据取自于海拔2 118 m与研究区相邻的小渠子气象站资料
3 讨 论
3.1 草地发展趋势
草地植物群落高度是反映草地群落数量特征的重要指标之一,高度变化常被作为草地群落动态研究考虑内容之一[13]。草地群落盖度高低可以反映草地植物同化面积大小和生物量积累的多少,群落中某种植物的盖度可说明该种在群落中的地位与作用[10]。
1985~2004年近20年的时间尺度内,草地群落种类组成优势种和主要伴生种,在群落中一直保持着原有的地位和功能作用,表现出较强稳定性。但在群落数量特征上,种间功能分化比较突出,优势种紫花鸢尾在盖度、地上生物量、优势度均表现出持续增长趋势,到上世纪末,在短短20年间,紫花鸢尾盖度从1998年以前的平均44.6%增长到1999年以后的57.1%、地上平均生物量从1985~1991年的263.6 g/m2,增加到1992~2004年的464.8 g/m2,增长了56.7%,群落种类组成原有结构失衡。
紫花鸢尾是一类低等牧草,在生长季内叶片有毒,各类家畜一般不采食;另外,紫花鸢尾株丛密集、耐践踏,再生性强,在持续过牧后,在其他草类暂得不到繁殖与更新时,而其能首先恢复生长与繁殖。紫花鸢尾所固有的饲用特点和生物生态学特性,是草地中种群数量持续增长的主要原因,草地在向劣质化演替,牧用价值下降。
3.2 引起草地波动与劣化的影响因素
植物群落是一个动态的系统,每一特征的稳定性都是相对的,而且不同的群落类型稳定程度各不相同。在自然状态下,群落波动性是草地适应环境的响应,群落通过自我不断的调节,以保持草地发展与生态过程的协调性[13-16]。研究分析结果表明,降水与牧压是造成群落各项数量特征年度波动、紫花鸢尾种群持续增长的主要因数,年度间降水的不稳定性引发草地数量特征的波动,而持续超载的牧压是促使紫花鸢尾种群在群落中持续增长,草地质量变劣的主因。
4 结 论
4.1通过对近20年时段草地群落的动态分析,草地植物群落组成优势种和主要伴生种是稳定的,以杂类草与禾草层片构成的群落格局没有发生改变。杂类草成片中紫花鸢尾、草原老鹳草出现率达100%,禾草层片中以草地早熟禾出现率达93.33%,异燕麦为80%。
4.2群落优势植物紫花鸢尾盖度、地上生物产量以及在群落中的优势度随着时间推移持续增长,盖度呈递增趋势,介于30%~50%;生物量呈先增加后降低再增加的趋势,最大年份达到548.3 g/m2。
4.3草地群落始终处于波动状态,而这种波动在一些年间表现出较大幅度,但无论是大幅度的波动还是小幅度的变化,均具有可逆性。年度间降水的不稳定性是引发草地群落数量特征年度波动的主要因素。
