干旱胁迫下鸭茅苗期生长特性及耗水规律
2022-07-13李江艳张鲜花袁小强刘雯欣
李江艳,张鲜花,袁小强,袁 惠,刘雯欣
(新疆农业大学草业与环境科学学院/新疆草地资源与生态重点实验室,乌鲁木齐 830052)
0 引 言
【研究意义】天然草地补播建植因苗期干旱缺水或缺少适宜当地草种而受到影响,干旱限制了草地补播、建植及生态环境的修复,选择优良草种、恢复天然草地植被及建植高产优质人工草地,已成为亟待解决的重大问题。新疆具有丰富的野生牧草种质资源,鸭茅(Dactylisglomerata)即是一种优良牧草,因再生性强,适口性好,高产优质、适应性好等特点[1-2],是建植人工割草地、刈牧兼用草地及改良天然草场的优等牧草[3]。新疆属典型的大陆性干旱气候,干旱少雨且草地退化严重,解决草地退化问题,需通过天然草地补播改良、退化草地恢复重建等一系列措施,可有效恢复和重建受损生态系统,提高草地覆盖度及生产力,实现草地的可持续发展。新疆属典型的大陆性干旱气候,干旱少雨且草地退化严重,筛选优良抗旱性的草种是生态恢复的必要措施之一。【前人研究进展】轻度干旱对鸭茅株高无显著影响,而中度和重度干旱下株高显著低于对照及轻度干旱处理,随干旱加剧,其根冠比也逐渐提高[4]。对采自全国各地不同生态环境下的16个代表种进行筛选结果显示,来自新疆的材料的耐热性和抗旱性普遍较好[5]。【本研究切入点】不同种源的鸭茅(Dactylisglomerata)应对不同干旱胁迫表现不一,鸭茅喜温和湿润气候,在新疆分布较为广泛,但新疆属于干旱半干旱区域,春季干旱少雨,大部分鸭茅在补播后苗期不能正常发育,制约了天然草地补播建植。针对鸭茅抗旱适应性研究,主要集中于萌发期PEG胁迫[6-7]、生理生化及分子机制[8-10],苗期干旱适应性研究较少。【拟解决的关键问题】以新疆野生鸭茅和栽培鸭茅为材料,设定不同水分梯度,研究水分胁迫下鸭茅的生长特性及耗水特性,分析鸭茅苗期生长发育特性和抗旱适应性,为天然草地重建、补播及草种的选择提供理论依据。
1 材料与方法
1.1 材 料
设置2017~2019年的大田试验,筛选出生长性能较好的5份野生驯化材料(采自于中国新疆阿尔泰山地、天山及准噶尔西部山地野生种质资源材料),2份表现较好的栽培品种:来自中国四川农业大学的栽培种(YM-6,宝兴鸭茅)及美国俄勒冈州栽培种(CYM-6,美国俄勒冈鸭茅)。表1
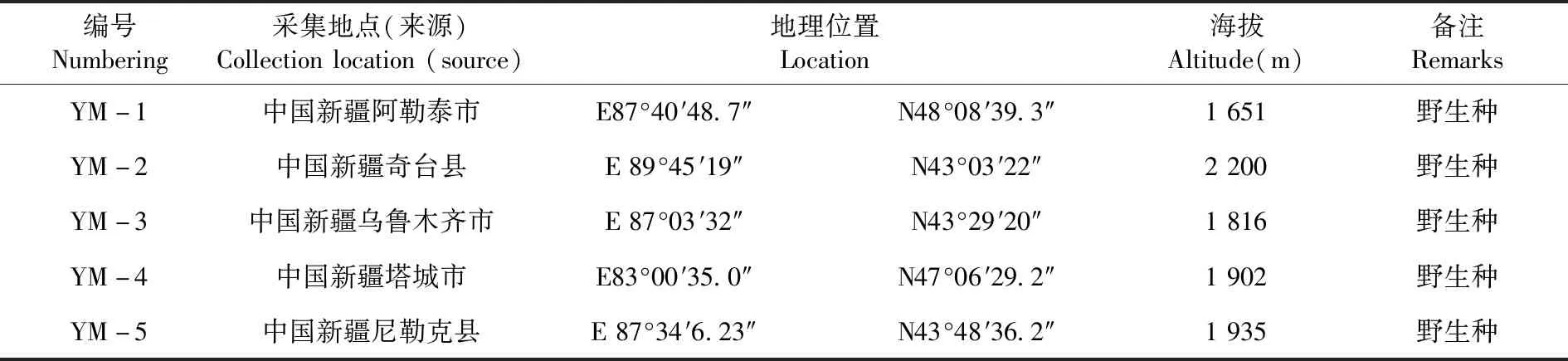
表1 材料编号及来源地Table 1 Material number and origin
1.2 方 法
1.2.1 育苗及移栽
2019年10月下旬挑选出7份鸭茅材料均匀饱满种子各300粒,均匀置于培养皿中,每个培养皿100粒种子,每个材料3个重复。放置光照培养箱内,按照10 h光照,14 h黑暗处理,定时补充水分,待幼苗长到5 cm高度,2019年11月上旬将各材料中长势一致的幼苗移栽至底部无孔,上口径11 cm,下口径7 cm,高18 cm的花盆中,每个花盆中装有过2 mm筛等量的沙子,每个盆中移栽1个单株,测定的饱和田间持水量为232.6 g。移栽后的幼苗置于新疆中泰创新技术研究院农业实验室(植物工厂)内进行后续试验,试验区温度为25±1(℃),湿度为46±1(%),每天保证10 h的光照时间。
1.2.2 干旱处理
移栽后,按照测定的土壤饱和持水量充分灌溉,待鸭茅单株能适应移栽环境并正常生长,于2019年11月下旬将植株统一修剪至7 cm高度。参照覃凤飞等[11]实验设计设置3个水分处理,适宜水分(L)、中度干旱(M)和重度干旱(H),分别为饱和田间持水量的85%、60%及35%。每个材料每个处理设置20个重复,共计420盆,同时设置对照(裸土),以除去盆中土壤表面的蒸发水分。处理期间,用称重法控制土壤含水量,每周补充灌溉至相应的土壤含水量,并指标测量,直至重度干旱下死亡率达到30%以上,于2020年1月上旬结束干旱处理。
1.2.3 测定指标
1.2.3.1 生长特性
干旱处理开始后每周测量植株高度,计算生长速度,处理结束后,测定绝对高度,统计分蘖数,将鸭茅植株齐地面刈割,并将根系部分冲洗干净,分别于烘箱85℃烘干后记录地上及地下生物量,并计算根冠比。
1.2.3.2 耗水特性
从干旱处理开始使用称重法确定土壤含水量,排除对照盆中土壤水分蒸发后,以盆重的减少量为幼苗的耗水量,并加水补充至设定土壤含水量。耗水量测定采用量程为0.1 kg的电子秤称重,周耗水量为每周耗水量总和,总耗水量为干旱处理内耗水量的总和。称量单株干物质量,计算耗水量,用公式水分利用率(WUE)= 干物质量/耗水量计算水分利用效率。
1.2.3.3 综合评价
采用公式对各指标进行函数值计算。
(1)
(2)
式中:Uij为鸭茅材料第j个指标的隶属函数值,Xij为鸭茅材料第j个指标的测定值,Xjmin为各材料j性状最小值,Xjmax为各材料j性状最大值,当j性状与抗旱性呈正相关采用公式(1)计算函数值,反之用公式(2)。
将各指标在不同水分条件下的函数值求平均,对所有指标平均函数值进行累加并求平均值,均值越大,抗旱性越强,反之越弱。
1.3 数据处理
采用Microsoft Excel软件进行数据汇总及制图,采用SPSS2 5.0 软件中单因素ANOVA检验法对各指标进行统计分析。
2 结果与分析
2.1 干旱胁迫下鸭茅生长速度变化
研究表明,处理前,7份材料均保持较高的生长速度,随着干旱时间的延长,鸭茅生长速度受水分条件的限制,生长逐渐减缓,均呈现先下降后升高再下降的趋势。生长速度总体呈现出L(适宜水分)>M(中度干旱)>H(重度干旱)。
水分胁迫前,各材料生长速度具有一定的差异,表现出生长速度的不一致性,其中YM-6及YM-7生长初期生长速度最大,分别达到1.876和1.321 cm/d;水分胁迫后,随水分胁迫增大,生长速度受到抑制,各材料间差异逐渐缩小,表现出一致的生长趋势,但YM-6和YM-7始终保持较高的生长速度,胁迫结束时生长速度分别达到0.253和0.351 cm/d,远高于其他材料,YM-6和YM-7生长速度高于其他材料。图1
2.2 干旱胁迫下鸭茅植株高度变化
研究表明,7份鸭茅材料随水分胁迫程度增大,株高呈现逐渐降低的趋势,具有一定的规律性。株高均呈现出L(适宜水分)>M(中度干旱)>H(重度干旱)。
无论何种水分梯度下,株高均呈现出栽培品种高于野生驯化材料,YM-6和YM-7在重度干旱下株高分别达24.75和26.25 cm,显著高于其他几个材料。图2
注:大写字母表示同种材料在3个不同水分下的显著差异(P<0.05),小写字母表示同一水分条件下各材料间的显著差异(P<0.05),下同
2.3 干旱胁迫下鸭茅分蘖数变化
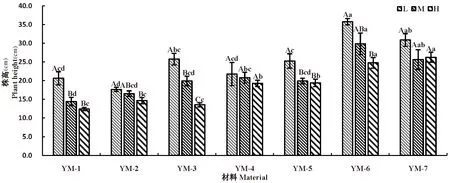
研究表明,7份材料随水分胁迫程度增大,分蘖数呈现先降低后升高的变化趋势,表现为L(适宜水分)>H(重度干旱)>M(中度干旱)。
不同水分胁迫下,均呈现出野生驯化材料高于栽培品种。栽培品种在中度干旱下平均仅有4个分蘖。野生材料随胁迫程度的增大,分蘖数呈先下降后升高的趋势,野生材料较栽培材料具有一定的耐旱性。图3
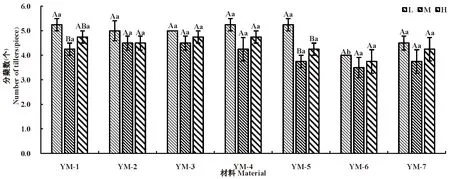
图3 干旱胁迫下鸭茅分蘖数变化Fig.3 Changes in the number of tillers of Dactylisglomerata under drought stress
2.4 干旱胁迫下鸭茅生物量及根冠比变化
2.4.1 生物量变化
研究表明,随水分胁迫程度增大,地上及地下生物量呈逐渐降低趋势,均呈现出L(适宜水分)>M(中度干旱)>H(重度干旱)。
不同水分胁迫下,YM-1地上生物量最低,重度干旱下仅有0.11 g,其余野生驯化材料与栽培品种之间地上生物量差异不显著。各材料地上生物量差异基本呈现出L(适宜水分)条件下地上生物量下均显著(P<0.05)高于其他水分处理,在M(中度干旱)及H(重度干旱)条件下差异不显著。
不同水分胁迫下,野生驯化材料YM-5较其余材料均保持较高的地下生物量,在中度及重度干旱下分别达到0.75和0.72 g,但差异不显著。各材料在3种水分胁迫下差异均不显著,仅YM-6在L(适宜水分)条件下显著高于其余处理。图4
图4 干旱胁迫下鸭茅生物量变化Fig.4 Changes of Dactylisglomerata biomass under drought stress
2.4.2 根冠比变化
研究表明,随水分胁迫程度增大,根冠比基本呈现升高的趋势,不同水分胁迫下,野生材料根冠比均大于栽培材料,YM-1在中度及重度干旱下分别达到4.16和5.09,差异显著(P<0.05)。
随水分胁迫程度增大,野生材料基本呈现出在H(重度干旱)条件下根冠比最大,均显著(P<0.05)高于L(适宜水分)条件,栽培材料随着水分胁迫的增大根冠比变化不显著。表2
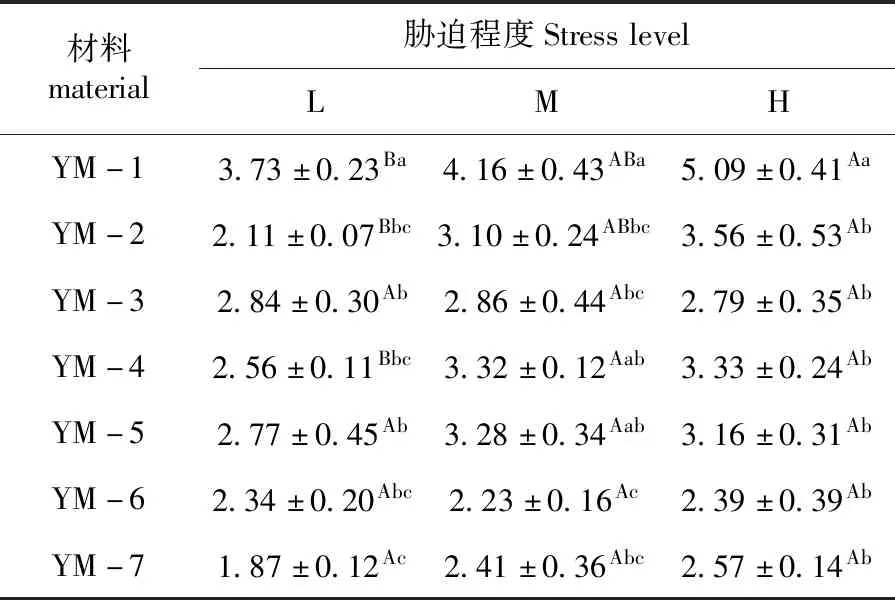
表2 干旱胁迫下鸭茅根冠比Table 2 Root-shoot ratio of Dactylisglomerataunder drought stress
2.5 干旱胁迫下鸭茅耗水变化
研究表明,随水分胁迫程度增大,周耗水量逐渐降低,各材料耗水曲线峰值出现的时间大致相同,总体表现为L(适宜水分)>M(中度干旱)>H(重度干旱)。
栽培材料和野生材料在L(适宜水分)条件下,周耗水变化趋势相似,均为先升高后降低的趋势,均在第四周达到最大值,最高耗水为32.38 g,M(中度干旱)及H(重度干旱)条件下,生长代谢受水分胁迫影响,周耗水始终保持较低水平,且变化相对平缓,3种水分胁迫下,周耗水最终趋于一致。图5
随水分胁迫程度增大,总耗水量逐渐降低,总体表现为L(适宜水分)>M(中度干旱)>H(重度干旱)。
各材料在水分充足时耗水多,而水分亏缺时耗水较少,不同水分胁迫下,各材料间总耗水差异不同,在L(适宜水分)下各材料间的总耗水差异不大,最高为146.7 g,最低为126.2 g,M(中度干旱)下YM-1总耗水最高,为102.4 g,而H(重度干旱)栽培材料总耗水最高,YM-6及YM-7总耗水分别达到63.9和73.8 g,多数野生材料保持较低总耗水。3种水分胁迫下,多数野生驯化材料总耗水差异显著P<0.05),而栽培材料均表现出在L(适宜水分)条件下总耗水显著(P<0.05)高于其他2处理。图6
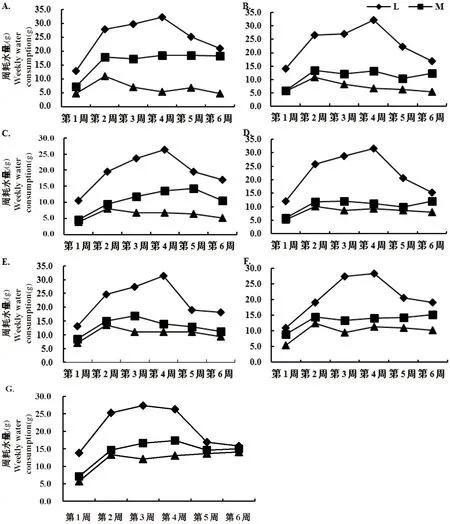
注:字母代表不同材料A.YM-1 B.YM-2 C.YM-3 D.YM-4 E.YM-5 F.YM-6 G.YM-7
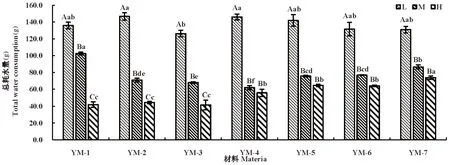
图6 干旱胁迫下鸭茅总耗水变化Fig.6 Changes in total water consumption of Dactylisglomerata under drought stress
2.6 干旱胁迫下鸭茅水分利用率变化
研究表明,7份材料在适宜水分下的生物量增量最大,而随水分胁迫加重,各材料干物质累积明显减少,但随水分条件的降低,不同材料水分利用率(WUE)呈现升高趋势,呈现出H(重度干旱)>M(中度干旱)>L(适宜水分)。
L(适宜水分)条件下,栽培品种保持较高水分利用率,YM-6及YM-7水分利用率分别达到2.66和2.99 g/kg,随着干旱加剧,野生驯化材料水分利用率逐渐升高,部分材料高于栽培品种,YM-3在重度干旱下水分利用率达5.2 g/kg。野生材料较栽培材料具有一定的耐旱性。表3
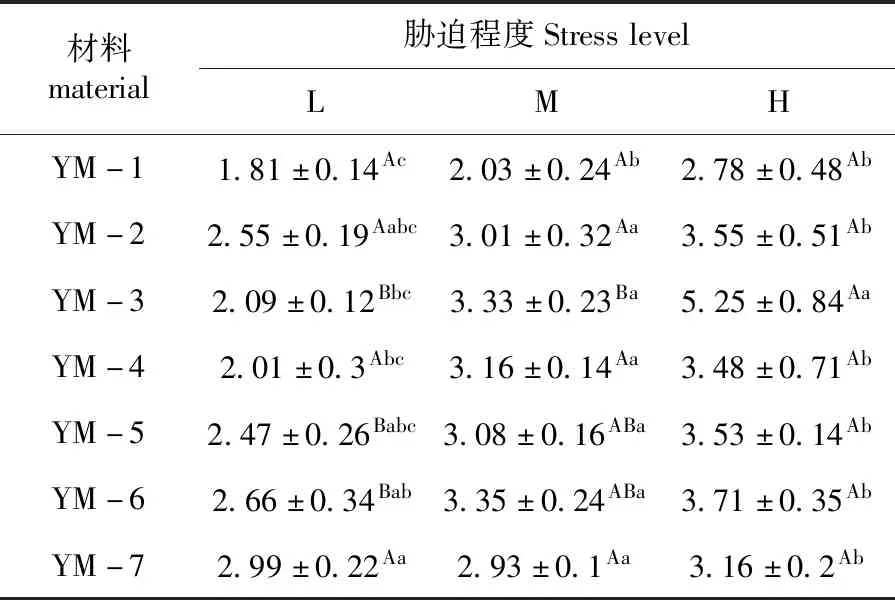
表3 干旱胁迫下鸭茅水分利用率Table 3 WUE of Dactylisglomerataunder drought stress(g/kg)
2.7 不同鸭茅材料抗旱性综合评价
研究表明,不同材料抗旱性强弱依次为YM-5>YM-7>YM-6>YM-4>YM-2>YM-1>YM-3。表4
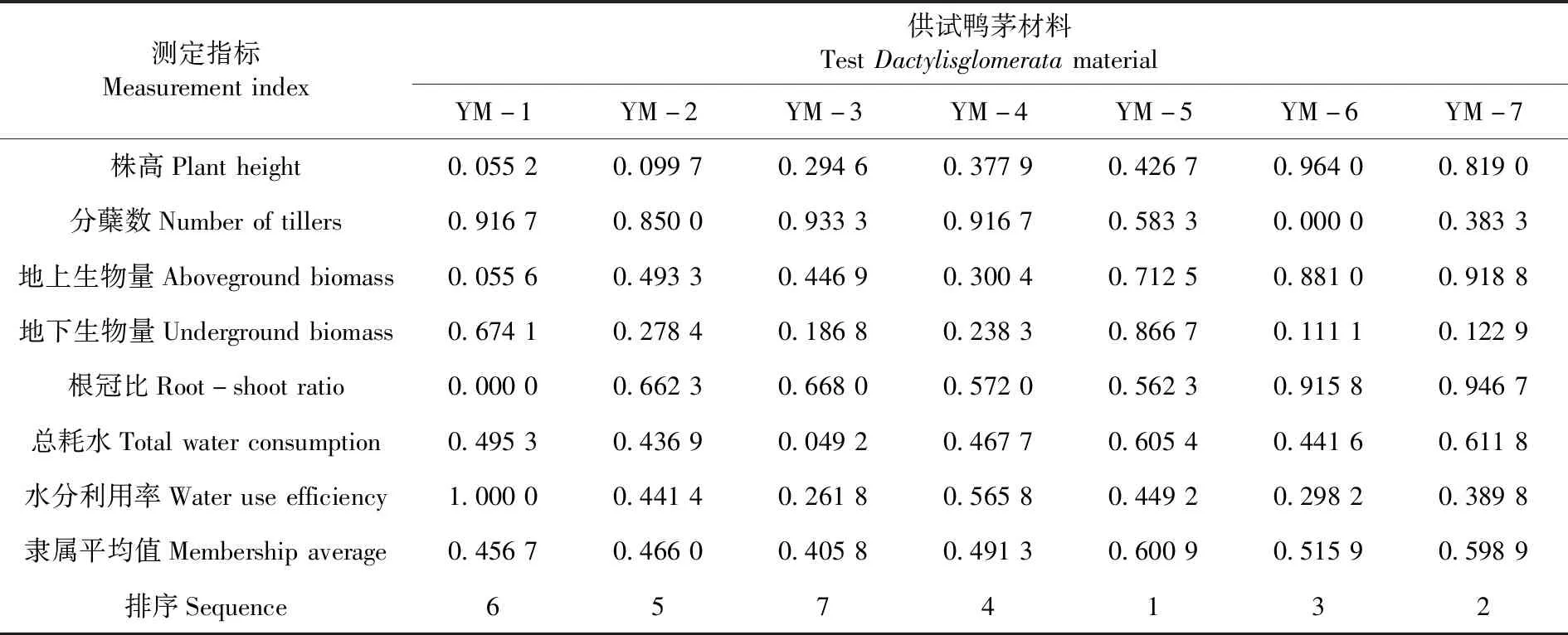
表4 不同鸭茅材料的抗旱隶属函数值Table 4 Drought resistance membership function values of different Dactylisglomerata
3 讨 论
植物根系的生长发育状况直接影响到地上部的生长,根冠比能够反映出植物的生长环境条件对根系与地上部分生长的不同影响[12]。植物生长速率是植物活力及植物对环境胁迫反应的重要指标[13]。干旱胁迫对植物的生长具有抑制作用[14]。朱铁霞等[15]研究干旱对紫花苜蓿(Medicagosativa)幼苗生长的影响,发现随着胁迫时间延长,幼苗生长速度受到严重抑制。周海军等[16]发现同一品种钝叶草(Stenotaphrumhelferi)的匍匐茎生长速度随干旱胁迫程度加重而呈现出明显下降趋势。研究结果也体现了上述结论,随着干旱时间的延长,鸭茅生长速度受水分条件的限制,生长速度逐渐减缓,呈现出先下降后升高再下降的趋势。在不同水分梯度下,适宜水分下鸭茅生长速度高于中度干旱及重度干旱条件。其次植物生长对水分的缺乏有着极高的敏感度,土壤中水分不足,导致植物营养不足,生长发育迟缓,株高降低、分蘖增多、地上生物量降低、地下生物量增大等现象。其中分蘖是植物重要的生物学特性,植物对环境的适应及群体的自动调节,很大程度上是通过分蘖消长来实现的[17],研究中,随水分胁迫程度的增大,7份鸭茅分蘖数呈现先降低后升高的趋势。水分条件还影响生物量的分配,水分胁迫下,地上生物量呈逐渐下降趋势,生物量向根部的相对分配增加。在适度水分条件下,植物会将生物量优先向根和茎部分配,但随着干旱程度增加,植物将减少地上生物量分配,集中向地下生物量分配。杨丹等[18]发现随着干旱胁迫程度的加剧,供试黑麦(Secalecereale)材料的平均相对地上生物量、地下生物量及相对分蘖数(RT)总体呈下降趋势。又如在干旱胁迫过程中,3个紫花苜蓿品种的地上生物量下降,为了缓解水分亏缺,其根冠比均增大[19]。有研究表明,耐旱性较强的棉花通过保持更好的光合性能和较大的根冠比,保持生殖器官干物质的累积以适应干旱胁迫[20]。研究中,随水分胁迫程度的增大,7份鸭茅生长受到不同程度的抑制,地下及地上生物量总体上呈逐渐降低的趋势,且根冠比呈上升趋势。
植物的蒸腾耗水量在不同的水分条件下存在着很大差异,不同水分条件对植物的生长发育具有不同程度的影响[21]。植物耗水量主要取决于灌水量,耗水量随着灌水量的增加而增大,土壤水分含量与植物的耗水量呈正比关系[22-24]。研究表明,鸭茅植株总耗水量随水分胁迫的增大呈现降低的趋势,在适宜水分条件下鸭茅周耗水呈现先增大后减少的变化,中度干旱和重度干旱条件下,鸭茅植株受水分限制,生长代谢受到影响,周耗水始终保持较低水平,呈现出适宜水分>中度干旱>重度干旱。在水分利用效率上发现,随着干旱程度增大,各材料的水分利用效率却呈现出逐渐增大的趋势,与大多数学者研究结果[22,25-26]相一致,即适当的水分胁迫,不但降低耗水量,反而在一定程度上提高水分利用效率,提高植物生物量。
研究中采用的供试材料为野生驯化材料和栽培材料,从植物生长发育速度、株高、分蘖、生物量及水分利用效率上来看,二者不尽相同。栽培材料和野生材料地上生物量差异不大,主要由于株高和分蘖两因素的消长实现,栽培材料生长速度及株高均高于野生材料,但野生材料具有较强的分蘖能力,地下生物量及根冠比远高于栽培材料,在某种程度上,野生材料适宜干旱环境的能力要高于栽培材料,且不同水分胁迫下栽培材料根冠比同样低于野生材料,尽管2种材料地上生物量没有明显差异,但野外材料应对干旱条件具有较强的适应性。综合生长特性及耗水特性,各材料各指标未呈现完全一致的变化。
4 结 论
4.17份鸭茅材料的生长发育特性存在差异,综合生长速度、株高、分蘖数、地上及地上生物量均表现出适宜水分>中度干旱>重度干旱,且随水分胁迫程度增加,7份鸭茅材料植株生长发育均受到不同程度的抑制。
4.27份鸭茅材料的耗水规律基本一致,随水分胁迫的增大,各材料耗水量逐渐降低,水分利用率逐渐增大,材料间耗水特性存在差异,中国四川宝兴和新疆尼勒克县鸭茅属于高耗水高水分利用率材料,而中国新疆乌鲁木齐市鸭茅属于低耗水高水分利用率材料。
4.3不同材料抗旱性强弱依次为中国新疆尼勒克县鸭茅>美国俄勒冈鸭茅>中国四川宝兴鸭茅>中国新疆塔城市鸭茅>中国新疆奇台县鸭茅>中国新疆阿勒泰市鸭茅>中国新疆乌鲁木齐市鸭茅。中国新疆尼勒克县鸭茅表现最优,其次为美国俄勒冈鸭茅,中国新疆乌鲁木齐市鸭茅表现较差。
