BnaTT2基因缺失黄籽甘蓝型油菜材料的创制
2022-07-12张彦锋陈娜娜韦世豪朱彦涛穆建新
安 然,张彦锋,陈娜娜,韦世豪,朱彦涛,穆建新
(陕西省杂交油菜研究中心,陕西 杨凌 712100)
油菜是我国第一大油料作物,也是多种轮作倒茬作物模式中不可缺少的作物,对我国的食用油供给安全和可持续发展具有重要意义[1]。甘蓝型油菜是我国油菜的主要栽培品种,对其品质进行改良,是促进我国油菜产业发展的重要环节。从营养价值来看,不同种皮颜色的甘蓝型油菜籽营养成分存在差异。有研究表明,在相同的遗传背景条件下,与黑籽油菜品种相比,黄籽油菜的含油量、蛋白质含量较高[2],硫代、芥酸等抗营养物质以及木质素和多酚的含量较低[3-4]。从生产方面来看,黄籽油菜的皮壳率低,出油率高,油色清亮,并且对菜籽油的产量提升也有一定的促进作用[5-7]。因此,黄籽油菜品种的创制对油菜生产和育种有着重要意义[8-9]。
黄籽甘蓝型油菜种质资源多样,但不存在天然的黄籽油菜材料,所以其遗传机理解析相对复杂[8]。1960年首个黄籽甘蓝型油菜被发现,之后研究者发现油菜种皮黑色素积累与类黄酮变化存在一定的相关性[10-15]。而作为类黄酮代谢途径中重要分支的原花色素,也被证实是调控种皮颜色的重要物质[16-17]。根据已知的十字花科类黄酮代谢途径,BnaTT2基因调控的4-二氢黄酮醇还原酶(DFR)、TT8基因调控的花色素还原酶等都是类黄酮合成途径中重要的转录因子[18-19]。因此,对这些基因的研究和利用是黄籽油菜品种选育的关键所在。
DFR能够使二氢榭皮素生成无色花色素,进而产生色素积累,形成深色油菜籽粒[20]。因此抑制DFR基因的表达是种皮颜色显黄的关键。甘蓝型油菜共有2个BnaTT2基因,分别位于甘蓝型油菜的A08和C08两条染色体上,均含有3个外显子和2个内含子。相关研究证明,BnaTT2基因能够有效调控DFR基因的表达[18]。因此,对BnaTT2基因进行研究与靶向敲除,是获得黄籽油菜材料的重要环节。
CRISPR/Cas9(clustered regularly interspaced short palindromic repeats/CRIS-PR associated nuclease 9)基因编辑技术是一种新兴的基因编辑技术,可以实现对靶向基因的精准敲除、序列替换和定点突变,快速创制目标突变体,已经逐步成为作物种质资源创制的重要手段之一,该技术在基因功能研究方面已逐渐成熟并成功应用于多种动植物。目前,植物CRISPR/Cas9系统已日臻完善,在拟南芥以及烟草、水稻、高粱、小麦、玉米等农作物中都已成功应用[21-25]。
本研究利用CRISPR/Cas9基因编辑技术及油菜下胚轴遗传转化等技术,对甘蓝型油菜类黄酮合成途径的关键基因BnaTT2进行靶向敲除,以创制BnaTT2基因突变体材料,并最终获得了黄籽的甘蓝型油菜材料,可为甘蓝型油菜育种提供新的种质资源。
1 材料与方法
1.1 材料与试剂
甘蓝型油菜(BrassicanapusL.,2n=38,AACC)受体材料K407、大肠杆菌Trans1-T1感受态细胞、根癌农杆菌GV3101感受态细胞等,均由陕西省杂交油菜研究中心油菜分子设计育种实验室提供;CRISPR/Cas9基因编辑载体pHSE401,由中国农业大学陈其军教授提供。引物合成及DNA测序均由北京奥科鼎盛生物科技有限公司完成。
1.2 方 法
1.2.1BnaTT2基因敲除靶位点序列设计 利用CRISPR/Cas9 在线靶位点设计网站(CRISPR RGET Tool)、油菜数据库(http://brassicadb.org/brad/index.php)以及NCBI提供的序列信息,对甘蓝型油菜基因BnaTT2进行序列比对,在不同拷贝的BnaTT2同源编码区上寻找包含CRISPR/Cas9系统可识别的共同靶位点核苷酸序列,设计靶位点,合成靶位点序列用于基因编辑,靶位点引物序列为17-TT2-P1(5′-ATTGTGTTCCACGGCGAAGGA-AAA-3′)和17-TT2-P2(5′-AAACTTTTCCTTCG-CCGTGGAACA-3′)。
1.2.2 载体的构建 构建载体参考Wang等[26]的方法。以甘蓝型油菜DNA为模板,利用引物17-TT2-P1/17-TT2-P2进行PCR扩增。PCR扩增程序为:95 ℃ 5 min;95 ℃ 30 s, 60 ℃ 30 s,72 ℃ 40 s,32个循环;72 ℃ 10 min。反应体系20 μL:DNA模板2 μL,引物各0.5 μL,Mix 10 μL,ddH2O 7 μL。将扩增片段连入CRISPR/Cas9载体pHSE401上,命名为pHSE401-TT2。测序检测后,利用电击法将pHSE401-TT2转入根癌农杆菌GV3101感受态细胞中,测序检测后将其命名为GV3101-TT2。
1.2.3 甘蓝型油菜的遗传转化 利用农杆菌介导法对甘蓝型油菜下胚轴进行遗传转化[27]。甘蓝型油菜种子经体积分数75%乙醇消毒1 min后,再用体积分数15%次氯酸钠(含吐温200 μL)消毒15 min,并用ddH2O漂洗。点播于1/2 MS培养基上,暗培养7 d。切取油菜下胚轴约10 mm作为外植体,使用含重组质粒的农杆菌 GV3101-TT2菌液(OD为0.4~0.6)侵染10 min,再用M1培养基(MS培养基+1 mg/mL 2,4-D溶液1 mL)共培养2 d,然后依次转入M2培养基(M1培养基+50 mg/mL潮霉素溶液50 μL)、M3培养基(M2培养基+0.1 mg/mL吲哚乙酸溶液250 μL)中继续培养,待产生幼芽后,切取芽体转入生根培养基M4(M1培养基+0.1 mg/mL α-萘乙酸溶液100 μL)中生根,生根后移栽至含基质的土壤中炼苗,最后转至人工气候室继续培养,直至收获种子。
1.2.4 阳性植株鉴定 将遗传转化后的幼苗(T0代)移栽至土壤中,用碱裂解法(SDS法)提取幼嫩叶片DNA,用载体骨架序列特异性扩增引物U6-F/CRIS-R(表1)进行靶位点PCR检测。PCR反应程序为:95 ℃ 5 min;95 ℃ 30 s,57 ℃ 30 s,72 ℃ 40 s,32个循环;72 ℃ 10 min。反应体系20 μL:DNA模板2 μL,引物各0.5 μL,Mix 10 μL,ddH2O 7 μL。连接、转化后挑取单克隆送检测序,筛选出含有载体骨架序列的植株。
1.2.5 阳性株系变异位点检测 根据油菜数据库提供的基因序列信息,针对甘蓝型油菜BnaTT2基因在所检测到的靶位点侧翼设计特异引物F2/R2(表1),对含有载体骨架序列的植株进行靶位点序列扩增,PCR反应程序为:95 ℃ 5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃40 s,30个循环;72 ℃ 10 min。反应体系20 μL:DNA模板2 μL,引物各0.5 μL,Mix 10 μL,ddH2O 7 μL。对目标片段进行克隆和测序分析,解析BnaTT2基因的突变位点。
1.2.6 T1和T2代植株靶位点的检测 将利用CRI-SPR/Cas9获得的BnaTT2基因突变体植株(T0代)自交套袋后收获种子(T1代),从中选取黄色籽粒T1代种子进行种植。在苗期提取幼嫩叶片DNA,用引物F2/R2(表1)进行靶位点PCR检测,PCR扩增程序和体系与1.2.5节同。对发生靶位点突变的T1代植株套袋自交并收获种子(T2代),选择黄色籽粒的T2代种子继续种植。苗期提取叶片DNA后用引物F2/R2(表1)进行靶位点PCR检测,PCR扩增程序和体系与1.2.5节同,验证靶位点突变情况。

表1 CRISPR/Cas9敲除基因位点检测所用的引物
1.2.7 油菜籽粒生物学特性分析 利用SLY-C微电脑自动数粒仪和LT1002B电子天平测定油菜籽粒的千粒质量,测3次后取平均值。黄籽粒色观测按照便于量化的原则,采用李加纳等[28]的6 级标准,肉眼观察确定种子颜色等级(color index,CI)。单颗籽粒中,黄色部分占总籽粒面积≥80%为5级,≥60%~<80%为4级,≥40%~<60%为3级,≥20%~<40%为2级,<20%为1级,完全褐籽或黑籽为0级。利用公式CI=∑(等级数×该等级籽粒数占所有等级籽粒数的百分比)计算单株的平均颜色等级。利用近红外光谱分析仪(BURKER MATRIX-I)测定T2代黄色油菜籽粒的脂肪酸成分和含油量,计算各组分相对含量,以K407材料为对照,利用SPSS Statistics 26进行t检验比较二者的差异。
2 结果与分析
2.1 甘蓝型油菜目的基因靶标片段的选择与构建
从供试甘蓝型油菜中扩增BnaTT2基因序列,与NCBI数据库中的同序列进行比对,发现BnaTT2基因在甘蓝型油菜中有2个拷贝。利用在线设计软件(CRISPR RGET Tool),在BnaTT2基因第1个外显子上选择2个拷贝的共有序列为靶位点序列,构建BnaTT2基因编辑载体,具体靶位点序列见图1。

图1 甘蓝型油菜BnaTT2基因的靶位点序列
2.2 甘蓝型油菜基因编辑植株的鉴定与分析
使用农杆菌介导法对甘蓝型油菜下胚轴进行遗传转化,共获得T0代植株16株,提取对照材料K407和T0代植株的DNA,利用载体引物U6-F/CRIS-R和靶位点侧翼设计特异引物F2/R2对其进行PCR检测,有4株T0代植株靶位点发生了突变。对4株T0代植株套袋自交后收获油菜籽粒(T1代)。T1代黄色籽粒油菜植株种植后利用引物F2/R2进行PCR检测发现, 有6株植株存在位点突变(存在杂合体植株),套袋自交收获T2代油菜籽粒。T2代黄色籽粒种子种植后进行PCR验证,最终确定有2株能够稳定遗传的纯合体BnaTT2基因突变体植株(TT2-Mutant-1和TT2-Mutant-2),其碱基突变位点见图2。图2表明,TT2-Mutant-1突变体在靶位点存在单碱基A的插入,TT2-Mutant-2突变体则是在靶位点存在单碱基A的缺失。

□.碱基插入位点;│.碱基缺失位点.CK.对照材料K407;TT2-Mutant.BnaTT2基因缺失T2代材料;图3同
2.3 油菜籽粒的生物学特性
2.3.1 千粒质量及粒色 经测定,TT2-Mutant和K407的千粒质量分别为4.74和3.62 g,TT2-Mutant的千粒质量较对照K407增加了1.12 g。黄色油菜籽粒的黄色度等级为4.8。籽粒颜色和籽粒大小对比如图3所示,每行40粒种子,基因编辑材料TT2-Mutant油菜籽粒与对照K407相比明显变大,种皮颜色由黑色(K407)转变为黄色,这表明BnaTT2基因的靶向敲除能够引起油菜籽粒颜色的改变。

图3 甘蓝型油菜突变体籽粒颜色和籽粒大小的变化
2.3.2 脂肪酸成分 T2代油菜籽粒的脂肪酸成分测定结果(图4)显示,TT2-Mutant材料各组分相对含量分别为:油酸(56.08±0.17)%,亚油酸(21.49±0.31)%,亚麻酸(12.73±0.17)%,芥酸(0.92±0.23)%;含油量(47.25±0.16)%。相比对照K407,黄籽油菜籽粒的含油量、亚油酸和亚麻酸相对含量分别提升6.10%,3.13%和4.99%;而芥酸及油酸相对含量则分别下降1.53%和1.01%。t检验结果显示,与对照材料K407相比,黄籽油菜籽粒油酸相对含量存在显著差异(P<0.05),而亚油酸、亚麻酸、芥酸相对含量和含油量存在极显著差异(P<0.001),表明对BnaTT2基因的靶向敲除一定程度上影响了油菜籽粒的脂肪酸组分。
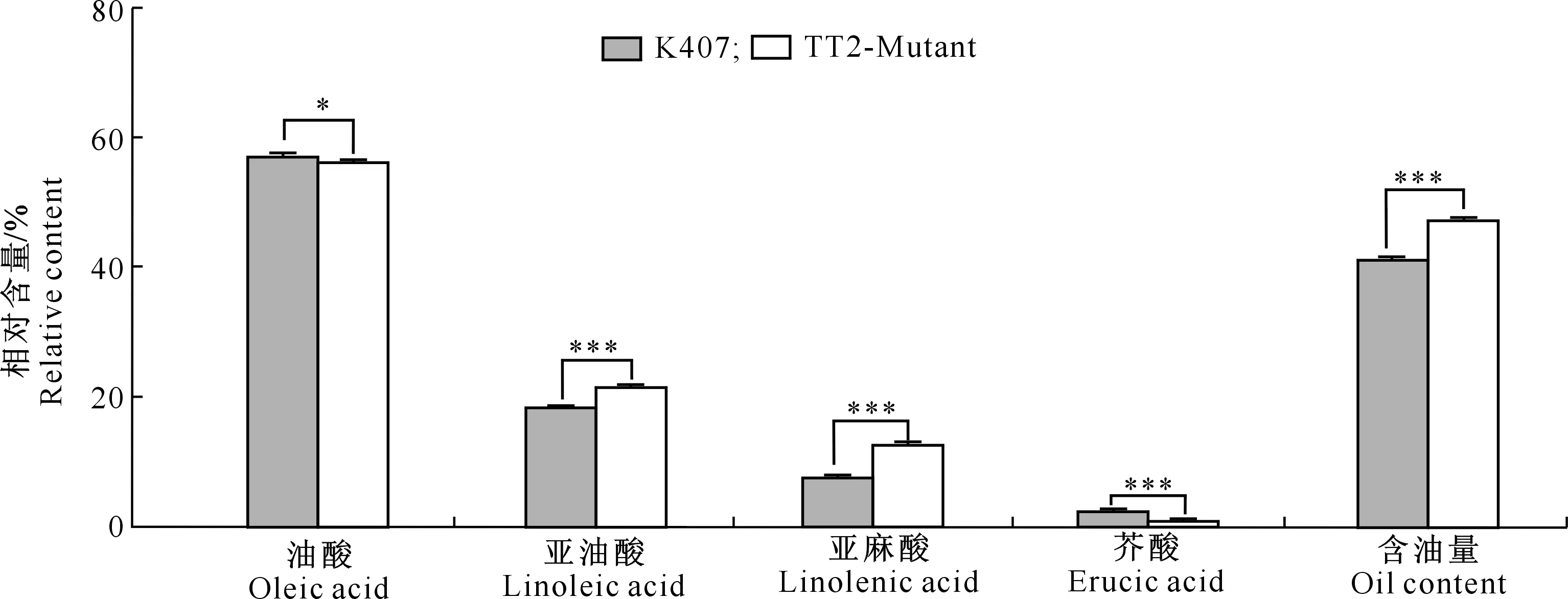
*.P<0.05;***.P<0.001;TT2-Mutant均为2株突变体(TT2-Mutant-1和TT2-Mutant-2)的平均值
3 讨论与结论
油菜作为我国主要的油料作物,其含油量高低、油分品质优劣对我国的油菜产业发展有至关重要的作用。较黑籽油菜而言,黄籽油菜有着高含油量、高蛋白含量、低硫苷、低芥酸等优点,因此对其相关功能基因进行研究,对油菜品质改良具有重要的理论与实践意义。
基因敲除技术的广泛应用,使得基因功能研究简单易行。而CRISPR/Cas9介导的基因编辑技术能够诱导基因组双键断裂,并通过细胞自身的非同源末端重组修复途径造成若干碱基的插入、缺失和替换,从而使基因发生移码突变,起到基因敲除的作用。本研究利用CRISPR/Cas9基因编辑系统对甘蓝型油菜的BnaTT2基因进行靶向敲除,利用农杆菌介导法成功获得2株黄籽油菜籽粒材料(TT2-Mutant-1和TT2-Mutant-2),对其BnaTT2基因靶位点进行测序分析显示,TT2-Mutant-1和TT2-Mutant-2在BnaTT2基因的2个拷贝上都存在单碱基插入或缺失,其中TT2-Mutant-1突变体在BnaTT2基因靶位点上存在单碱基A的插入,TT2-Mutant-2突变体则是单碱基A缺失。以上结果表明,本试验所构建的基因编辑载体能够准确地对BnaTT2基因进行靶向编辑,使该基因相关位点发生碱基突变;同时该碱基位点的突变能够影响BnaTT2基因的功能表达,使油菜籽粒的种皮颜色由黑色(基因编辑前)转变为黄色(基因编辑后),说明BnaTT2基因与油菜籽粒的种皮颜色密切相关。此外,BnaTT2基因靶向敲除材料TT2-Mutant的千粒质量相较对照增加了1.12 g,千粒质量的变化可能与BnaTT2基因的靶向敲除有关,也可能由其他原因引起,该部分变化原因有待进一步分析验证。
本研究发现,与对照K407相比,TT2-Mutant的脂肪酸组分相对含量发生了很大变化,其中含油量极显著提升了6.10%,表明BnaTT2基因功能缺失能够在一定程度上影响油菜籽粒的油脂相对含量,进而影响油菜的出油量;同时,BnaTT2基因的功能缺失还导致TT2-Mutant中的芥酸相对含量极显著下降1.53%,而亚油酸及亚麻酸相对含量分别极显著提升3.13%和4.99%,这表明BnaTT2基因功能缺失能够影响籽粒中脂肪酸的组成,可以有效提高对人体有益的脂肪酸含量,降低有害物质含量,可为优质精品食用油的创制提供新途径,为甘蓝型油菜育种提供新的种质资源。
