宽鳍鱲个性与维持代谢、食物占有能力和日常能量消耗的关联
2022-07-08胡月唐金玉付成付世建
胡月 唐金玉 付成 付世建
(重庆师范大学进化生理与行为学实验室,重庆市动物生物学重点实验室,重庆 401331)
个性是动物在行为上表现出来的稳定的个体间差异(Consistent individual difference,CID),包括活跃性、探索性、勇敢性和社会性等个性特征[1—3]。代谢反映生命体的能量消耗水平,是衡量机体生理状态的重要指标之一[4,5],与生物的生存适合度紧密关联[6]。其中标准代谢率(Standard metabolic rate,SMR)反映动物在非活动、无食物消化吸收状态下的最低能量消耗水平,因此可用来衡量动物的维持能量消耗[5,7]。日总能量消耗(Daily energy expenditure,DEE)是另一个重要的能量学参数,常用来表征动物每天的能量消耗总和,包含维持能耗、活动能耗和摄食消化(Specific dynamic action,SDA)能耗[8]。研究者发现机体的DEE一般与SMR紧密关联[8,9]。
研究者普遍认为机体能量代谢水平与个性行为特征协同进化[2,3,10,11]。生活节奏综合症(Pace of life syndrome,POLS)假说认为动物的代谢、行为、激素和免疫特征等在种内和种间水平上均存在一系列的关联[2]。积极个性的个体(如更为勇敢和活跃)可能具备较高的SMR、DEE及食物占有能力[2,3]。有关个性与SMR的关联已有较多报道,例如,大西洋鲑(Salmo salar)的SMR与攻击性呈正相关[12];欧洲海鲈(Dicentrarchus labrax)的SMR与活跃性和勇敢性呈正相关[13]。但是由于包括鱼类在内的水生生物在自由状态下的DEE难以测定,个性与DEE之间的关联尚未被证明。近年来,随着动物运动轨迹精细测定技术的发展,通过运动轨迹衡量能量代谢进而考察水生生物DEE与个性的关系成为可能。另外,个性与食物占有能力即摄食率(Feeding rate,FR)是否存在关联也鲜有报道。最近有关异育银鲫(Carassius auratus gibelio)的研究发现,单养时FR和个性特征在特定条件下存在关联[14]。另有研究发现棘鱼(Gasterosteus aculeatus)的勇敢性与FR呈正相关[15]。但以上研究均在单养的环境下实现,较高的FR可能更多地受食欲和消化道体积大小影响,个性本身所带来的种内竞争能力的差异对食物占有能力的影响无从体现。在自然界约50%已知鱼类种类至少在生活史的特定生活阶段具有集群生活的生态习性[16]。因此在群体生活状态下衡量的FR尽管在技术层面上较为复杂,但能够充分考虑个体食物占有能力的因素,具有单养时测定FR无法比拟的生态学意义。值得一提的是,最新研究发现鱼类在独处状态和在群体中对外界刺激产生截然不同的行为反应[17,18]。因此,本研究旨在考查个性与SMR、DEE和食物占有能力的关联。
凝聚力和协调性常作为研究鱼类集群运动特征最主要的指标[1]。凝聚力通常以群体中个体间距离和个体距群体质心的距离等来衡量,而协调性主要通过不同个体之间或个体与群体质心之间移动速度的同步性、移动方向的一致性(如夹角等)等来衡量[19,20]。个体在群体中表现出来的上述群体特征可能给其带来特定的生态收益和代价,例如距群体质心距离不同可能会直接影响食物获取概率。因此,本研究旨在考查不同个性鱼类个体的集群运动特征与能量代谢消耗和食物占有能力的关联。为完成研究目标,本研究选择宽鳍鱲(Zacco platypus)雌鱼为研究对象(宽鳍鱲雄鱼在野外常处于独居状态而雌鱼集群生活)。宽鳍鱲为一种广泛分布的小型鲤科鱼类,在自然界中通常形成大小不一的群体,近期我们对该种鱼的个性行为的重复性和指标间关联进行了先期考查[21]。本研究首先在单尾鱼条件下测定了实验鱼的SMR和个性特征(即勇敢性、探索性、活跃性和社会性),随后在集群条件下(6尾鱼)测定了每尾鱼的活动轨迹和摄食状况。实验鱼在集群条件下的轨迹还用于衡量活动能量消耗及集群状态下每尾鱼的运动特征(包括活跃程度、凝聚力和协调性)。
1 材料与方法
1.1 实验鱼来源及驯养
实验用宽鳍鱲雌鱼(5.81—6.87 g)采集自乌江重庆武隆段(29°24′37″N,107°31′55″E)。实验鱼运回实验室后在循环水养殖系统(1.2 m×0.55 m×0.55 m)中进行4周的驯养。驯养期间水温维持在 (25±1)℃,水体溶氧维持在7.0 mg/L溶氧以上。实验鱼每日上午10:00以解冻后的赤虫(中国滨海骏业生物公司)饱足投喂1次,投喂30min后使用虹吸管清理粪便残渣。每日换水1次,日换水量约为总水量的10%。光照周期为 12h∶12h。
1.2 实验方案
在驯养完成后,将麻醉(MS-222,80 mg/L)后的实验鱼经腹腔注射 PIT标签(广州市洪腾条码技术有限公司),方便实验鱼的识别。随后进行2周的恢复。2周后再次在实验鱼背部区域进行体外颜色标记,用于集群过程实验鱼的识别。1周后挑选身体健康、大小接近并且背部颜色标记清晰的80尾宽鳍鱲(6.42±0.14) g用于SMR、个性和集群行为的测定。实验鱼首先进行SMR和个性特征测定(测定顺序为:勇敢性、探索性、活跃性和社会性)。为确定4种个性行为是否稳定,所有的个性行为在第1天测定完成后均在第2天进行重复测定,以考查个性指标的重复性。最终的个性指标在数值上取2d的平均值。在个性测定完成后,从80尾实验鱼中挑选60尾,随机分为10组,每组6尾鱼,用于集群行为的拍摄。通过拍摄和分析6尾鱼构成群体的运动轨迹,获取每尾实验鱼集群运动时的各个行为特征,并计算DEE。此外通过记录每尾实验鱼摄入的食物量(赤虫的条数),计算FR,用以评价实验鱼的食物占有能力。
1.3 实验设备
个性中活跃性、探索性和勇敢性的测定采用实验室自制的鱼类个性测定仪(具体结构参见文献[1])来完成。该测定装置主体部分为一长方形水槽(60 cm×20 cm×20 cm),通过可拆卸的有机玻璃隔板(不透明)将上述水槽分为开放区(40 cm×20 cm×20 cm)和隐蔽区(20 cm×20 cm×20 cm),隐蔽区放置有一株塑料水草,可为实验鱼提供隐蔽场所。开放区和隐蔽区之间的隔板上安装有可打开、关闭的小门,小门打开时,实验鱼可以在开放区与隐蔽区之间来回穿梭。水深维持在10 cm以降低实验鱼的垂直移动。实验鱼的行为通过与远端监视器相连的摄像头(Logitech Pro 9000;罗技公司,中国苏州;15帧/s)获取。设备周围以白色不透明帆布隔开以避免外界干扰。
社会性由实验室自制的二元行为选择装置(具体结构参见文献[22])完成测定。装置的主体部分为一长方形水槽(70 cm×35 cm×35 cm)。水槽由两个透明有机玻璃隔板分隔为中间的选择区(40 cm×35 cm×35 cm)和两边的两个刺激区(15 cm×35 cm×35 cm)。水深10 cm,水槽四周贴以白色不透明广告纸,以降低外界干扰。社会性视频的获取方式与上述活跃性等相同。
集群运动的测定采用实验室自制的集群行为观察装置(图 1)完成。该装置由内部的近椭圆形的实验水槽A(120 cm×60 cm×10 cm)和外部的圆形大水槽B(直径150 cm,高30 cm)构成,外部的圆形水槽主要用于提供水浴环境,方便实验水槽A内的水温控制。实验水槽A由白色PVC板制成,利于减少四周环境干扰和增强实验鱼与背景的反差,方便后续视频分析。在实验水槽A的左右两端和中部分别开有1个小孔,3个小孔分别连接3根塑料管(C、D和E),用作食物投喂通道。水槽上方固定有摄像头用于集群视频的采集。水深控制为10 cm。

图1 鱼类集群行为测定装置结构示意图(俯视)Fig.1 The structure of the device used to determine schooling in fish species (top view)
1.4 参数测定方法
SMR使用实验室自制的流水式呼吸代谢仪(具体原理和结构参见文献[23])测定SMR[1]。将单尾宽鳍鱲禁食24h后转移至流水式呼吸代谢仪,进行恢复和适应。实验鱼在代谢仪中适应1d后采用溶氧仪(HQ30d,美国哈希公司)测定每个呼吸室水流出口处的水体溶氧值,并通过容量瓶(50 mL)和计时器计算流经每个呼吸室的水流速度,以此计算实验鱼的耗氧率。每尾实验鱼的耗氧率自09:00到17:00每1h测定一次,连续测定2d。一共获得18个耗氧值,实验鱼耗氧率(mg O2/h)的计算公式如下:
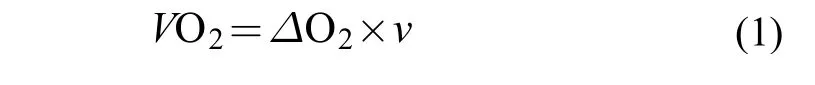
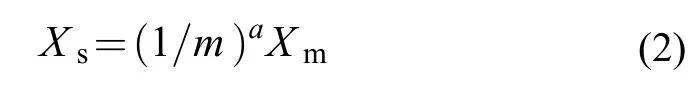
式中,Xs是标准耗氧率[mg O2/(kg·h)],Xm为公式(1)中计算出的单尾鱼耗氧率,a为体重系数,m为单尾鱼体重(kg)。最后以使用公式(2)进行体重矫正后即可得到每尾实验鱼的 SMR。
勇敢性将单尾实验鱼转移至鱼类个性测定仪的隐蔽区适应30min,随后将有机玻璃隔板上的小门轻轻打开,对实验鱼进行持续30min的行为拍摄。拍摄好的视频使用行为学软件idTracker进行后续分析。实验鱼首次进入开放区的时间(Latency to emerge,s)和进入开放区侦察的次数(Number of inspection)作为勇敢性的指标[35]。
探索性待勇敢性测定完毕,将实验鱼轻轻赶至开放区,并关闭隔板上的小门。随后在小门对面距离边缘15 cm处放置一个实验鱼从未见过的物体作为“新奇物体”(Novel object,蓝色有机玻璃柱;重复测定时换成褐色有机玻璃柱,直径和高度均为2 cm)。“新奇物体”放置完成后立刻开始探索性行为的拍摄,持续30min。拍摄好的视频使用行为学软件idTracker进行后续分析。实验鱼首次达到新奇物体的时间(Time to first approach object,s;距离物体表面3 cm以内则定义为到达)和距离物体中心的平均距离(Distance to object,cm)作为探索性的指标[35]。
活跃性在探索性测定完毕后,移除“新奇物体”,待实验鱼恢复10min后进行持续10min的活跃性拍摄,拍摄好的视频使用行为学软件idTracker进行后续分析。实验鱼的运动时间比(Time spent moving,%)和10min的总移动位移(Distance moved in 10min,cm)作为活跃性的指标[35]。
社会性实验鱼的社会性使用二元行为选择装置进行测定。首先在随机一侧的刺激区放置6尾实验鱼熟悉的同类作为刺激鱼群,另一侧空白。30min后把待测实验鱼转移至中部的选择区。在2min适应后,对实验鱼进行持续30min的行为拍摄。拍摄好的视频使用行为学软件idTracker进行后续分析。实验鱼位于偏好区的时间比(Percent time with shoal,%;距离刺激鱼群11 cm以内,即实验鱼3—4倍体长[24])及距离刺激鱼群的平均距离(Distance to shoal,cm)作为社会性的指标。社会性重复测定时刺激鱼群置于另一侧的刺激区[35]。
DEE与集群行为测定集群行为的测定在图 1所示装置中完成。将6尾实验鱼同时转移至装置中的实验水槽中。在8:30—9:00、10:30—11:00、12:30—13:00、14:30—15:00、16:30—17:00共5个时间段内进行视频拍摄用来衡量实验鱼在白天的活动轨迹和/或摄食量。在8:30和14:30时通过远端注射装置随机从3根塑料管投放解冻后的赤虫。投放时选择大小相似的赤虫,并记录称重。通过对实验鱼背部标记的肉眼观察可以对每尾鱼进行识别,并记录每尾鱼摄取的赤虫数量据此可计算每尾实验鱼的FR。其中一半实验鱼在18:00—6:00进行了测定,用以观察是否存在昼夜节律和白天夜晚的数据是否存在关联。宽鳍鱲个体活跃性等数据在白天、晚上存在关联(18:00—6:00的数据在光照条件下测定,故未纳入计算),因此以白天的集群和摄食情况衡量实验鱼的DEE不会影响本实验的主要结论(即关联)。每尾实验鱼在1d中的维持代谢消耗可通过SMR获得;摄食耗能可通过每尾鱼摄取赤虫的条数计算摄食量,再根据SDA系数获得单尾鱼的SDA耗能[25];运动耗能依靠集群视频获得每尾实验鱼在1d中的运动时长和平均速度,再根据宽鳍鱲运动速度与代谢率的关系计算相应的耗能水平[26]。维持消耗加上摄食耗能和运动耗能即为实验鱼的DEE。
集群行为指标采用idTracker进行分析[27]。通过该软件识别群体中的每尾鱼并给出每尾鱼实时的坐标点。随后再通过坐标点计算每尾实验鱼的运动时间比(Percent time moving,%)、平均游泳速度(Average swimming speed,cm/s)、速度同步性(Synchronization of speed)、距质心距离(Distance to centroid,cm)、游泳方向夹角(Separate swimming angle,rad)共5个集群参数。其中游泳速度的同步性由每尾鱼质心和鱼群质心移动速度计算得到[20]:
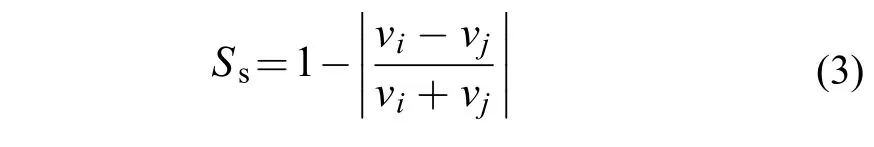
式中,Ss为速度同步性;vi和vj分别是特定实验鱼和鱼群的游泳速度。距离质心的距离用来衡量实验鱼的凝聚力,由以下公式计算[20]:

式中,Dij为特定实验鱼距鱼群质心的距离;xi,yi和xj,yj分别是特定实验鱼和鱼群质心每帧的坐标。每尾鱼和鱼群质心的游泳方向夹角由以下公式计算得到[20]:

式中,SSA为特定实验鱼与鱼群的游泳方向夹角(rad),xt、yt和mt、nt分别为单尾实验鱼和鱼群质心在第t(s)时的坐标,xt–1、yt–1和mt–1、nt–1分别为单尾实验鱼和鱼群质心在第t–0.5(s)时的坐标。
1.5 统计与分析
实验数据经Excel 2019进行常规计算后,采用SPSS 17进行统计分析。个性特征及集群特征与能量消耗和FR之间的关联采用Pearson相关性分析。所有数据采用平均值±标准误表示,显著性为P<0.05。
2 结果
2.1 个性特征与SMR、DEE和FR的关联
宽鳍鱲在个性行为、SMR和FR等方面均表现出较大的个体差异。耗氧率与体重的关系可拟合为y=0.3308x0.7847(R=0.507,P<0.001),SMR使用体重系数0.7847进行矫正后为(76.83±2.21) mg O2/(kg·h)。每天的维持代谢消耗、消化导致的能量消耗、运动导致的能量消耗和总代谢能量消耗分别为(34.19±1.32)、(45.11±3.81)、(74.50±3.27)和(153.80±4.84) kJ/kg。SMR、DEE和FR与个性(勇敢性、探索性、活跃性和社会性)的所有指标均不关联(表 1)。

表1 个性特征与标准代谢率、摄食率和日总耗能的关联Tab.1 The correlation between personality traits and standard metabolic rate,feeding rate and total daily energy expenditure
2.2 集群运动参数与SMR、DEE和FR的关联
宽鳍鱲DEE与集群运动参数中的运动时间比(R=0.524,P<0.001;图 2a)、平均游泳速度(R=0.571,P<0.001;图 2b)和距质心距离(R=0.284,P=0.031;图 2c)均呈正相关(表 2)。此外,DEE与集群过程中的FR也呈正相关(R=0.742,P<0.001;图 2d和表 2)。
表2 集群运动参数与标准代谢率、摄食率和日总耗能的关联Tab.2 The correlation between swimming character during shoaling and standard metabolic rate,feeding rate and total daily energy expenditure
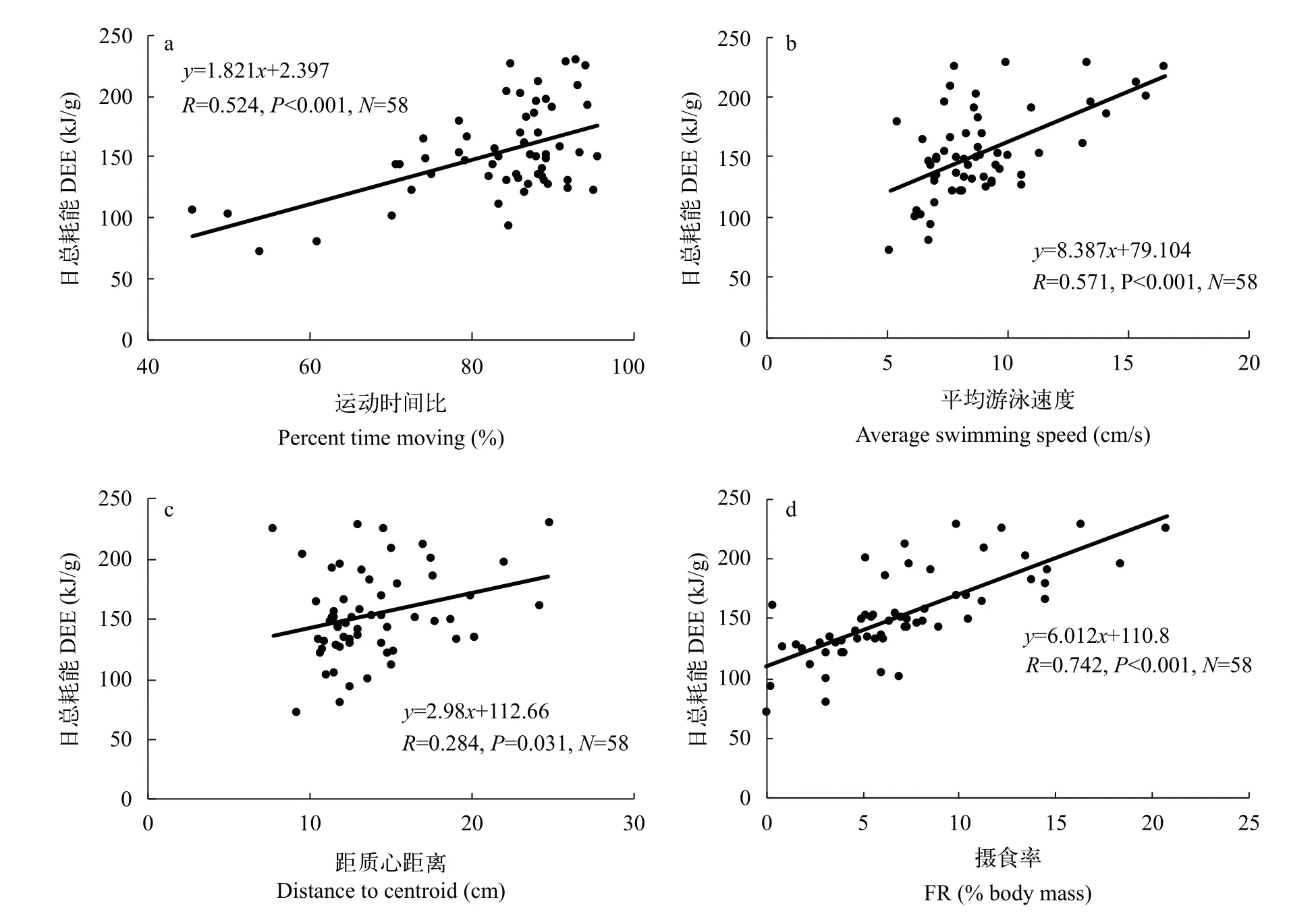
图2 日总耗能与集群运动参数(运动时间比、平均游泳速度和距质心距离)和摄食率的关联Fig.2 The correlation between total daily energy expenditure and swimming character during shoaling and feeding rate
3 讨论
3.1 个性与食物占有能力及能量代谢的关联
一般认为不同个性的动物会表现为从积极到消极的连续体[10]。在行为上表现积极的个体通常具有较高的活跃性、探索性和勇敢性及较低的社会性[3,28,29]。关于异育银鲫(Carassius auratus gibelio)的研究发现个性积极的个体表现为较高的维持代谢能量消耗,即SMR[14]。而本研究中所有个性指标与SMR均没有关联,这与以往的理论不符。原因之一可能是迄今为止鱼类行为学有关的个性测定均为单尾鱼,本研究也沿袭了传统的方法。但对于宽鳍鱲这种野生的集群性鱼类而言,在实验室进行单尾鱼标准代谢或个性行为测定时,可能存在某种应激。这就导致不同个体在实验室测定时展现出与自然状态下截然不同的行为模式,干扰了代谢以及个性特征的评估。因为应激导致的代谢上升的幅度可能与代谢、个性本身是不相关甚至是负相关的。例如较勇敢的个体可能对胁迫表现较低的应激反应[30],但这部分应激导致的代谢上升显然与个性特征不关联[31]。研究发现大西洋鲑(Salmo salar)在移除环境遮盖物后SMR与活跃性的关联丧失[32]。这种现象在集群和野生鱼类显然可能表现得更为明显。最新研究表明,鱼类个体在单独存在和在群体中时针对捕食者暴露展现截然不同的行为反应[17,18]。这提示将来针对宽鳍鱲这类野生集群性鱼类的个性测定方法值得研究。
“代谢引擎”(Metabolic engine)假说认为个性与食物占有能力正相关[3],然而本研究发现宽鳍鱲个性特征与食物占有能力无关。个性与FR无关可能因为实验设定的摄食条件与野外存在很大差异,宽鳍鱲在野外主要以藻类为食[33],其食物资源在野外分布相对均匀和充足,这种个性相关的竞争能力可能更多地体现在探索和保卫领地、或繁殖竞争等,而对食物竞争并无强烈的选择压力。鱼类个性与日总耗能的关联,本研究发现二者不存在关联。因为多数个性特征,特别是与活跃性相关的特征相互妥协,进而导致相关个性在群体中丧失[34]。但是,近期有关宽鳍鱲的研究发现,社会性和勇敢性在集群时能够部分保留,表现为勇敢性与社会性分别与群体极性和协调性呈正相关[35]。因此该问题值得进一步深入研究。今后的研究可结合皮质醇测定等技术方法以排除应激对个性及个性测定的影响,同时集群性鱼类个性的单尾测定意义也值得商榷。
3.2 集群运动参数与食物占有能力及能量代谢的关联
鱼类在集群过程中需要不断调整自身的游泳速度和方向以维持和群体的同步性[15]。群体中位置差异或运动活跃性差异会影响个体在集群过程中的生态收益和代价[36,37]。在本研究中,日总耗能与集群运动参数中的运动时间比、平均游泳速度及FR均显著正相关。运动耗能占近一半的日总耗能,因此游泳时间更多、速度更快的个体日消耗总能量更高。同理,FR也与日总耗能正相关,因为消化引起的能量消耗占日总耗能的比例近30%,同时摄食过程中伴随的其他生理(如激素水平的上调)及行为的变化也可能导致额外的能量消耗的增加。因此,集群时较为活跃和摄食较多的个体具有更高的日总耗能。相比之下,维持代谢只占日总耗能的22%,对其贡献较少,与其没有显著关联。有研究发现花栗鼠(Tamias striatus)的标准代谢与日总耗能的关联仅在繁殖期存在,可能就与繁殖期花栗鼠标准代谢在日总耗能的占比大幅提高有关[38]。
群体凝聚力指标中距群体质心距离可以较好地反映特定个体对群体靠近或远离的趋势,因此一直备受关注[15,20,35]。本研究发现宽鳍鱲的距质心距离与日总耗能呈正相关,表明越靠近群体的边缘,日总耗能越高。在本研究中,与活跃性相关的运动速度、运动时间比均与距质心距离呈正相关(结果未展示),提示在群体中较为活跃的实验鱼可能更偏好群体的边缘,且日常总能量消耗更高。鱼类的个性特征会显著影响其在群体中的空间位置,例如更为勇敢或处于领导地位的个体通常具有较弱的集群倾向,更加远离群体质心[39]。近期关于宽鳍鱲个性与集群运动参数关联的研究发现个性较为积极的个体在群体中极性更强(与群体运动的方向一致性更好),但个性与群体凝聚力的关联并未发现[35]。
综上所述,宽鳍鱲的个性特征与食物占有能力和能量代谢均无关联;集群时较为活跃的成员往往处于鱼群的边缘,且食物占有能力更高,但日常能量消耗也越多。其内在机制值得进一步深入探讨。针对集群性鱼类,在群体中考察其个性和能量代谢特征及其生态收益和代价更具生态学意义。
