粗胫翠尺蛾视蛋白基因的克隆及灯光对其表达量的影响
2022-07-08全林发董易之李文景池艳艳陈炳旭
姚 琼,全林发,徐 淑,董易之,李文景,池艳艳,陈炳旭
(广东省农业科学院植物保护研究所/广东省植物保护新技术重点试验室,广州 510640)
0 引言
荔枝、龙眼是中国南方优势名贵水果,主要分布于广东、广西、福建、海南、四川及云南等省(自治区),是“南国四大果品”之首[1]。2019年全国荔枝和龙眼栽培面积分别高达54.1万和27.6万hm2,在农业经济中占有重要地位[2]。以尺蛾为主要类群的食叶害虫在荔枝、龙眼梢期所造成的叶片损失在病虫害造成的叶片损失中占比超过80%,对荔枝、龙眼高产稳产造成巨大威胁[1,3]。危害广东荔枝、龙眼的尺蛾种类主要为粗胫翠尺蛾(Thalassodes immissaria),该虫每年可发生7~8代,世代重叠,8—11月发生为害最严重[4-5]。粗胫翠尺蛾多于夜间羽化,羽化当晚交尾,交尾后第2天即可产卵,雌性成虫单雌产卵总量可高达180余粒,是荔枝、龙眼梢期的重点防控对象[3]。
视觉是昆虫感知外界信息的最重要途径之一,对昆虫取食、躲避天敌、求偶、产卵等行为起关键作用[6]。昆虫通过复眼上的光感受器将光信号转化成生物电位信号,再通过一系列级联反应,将其传递到中枢神经系统引起视觉反应[7]。其中,光感受器的感光色素因子的核心成分是视蛋白[7-8]。视蛋白是一种具有7个跨膜结构的G蛋白偶联受体,分子量约为30~50 kDa[9]。根据其对不同波长敏感性可分为紫外光视蛋白(ultraviolet wavelength sensitive opsin,UV),蓝光视蛋白(blue wave length sensitive opsin,BL)和长波长视蛋白(long wave length sensitive opsin,LW)3类,3种视蛋白在应对不同光源中发挥不同的作用,其对应的不同波长分别为300~400、400~500、500~600 nm[8-11]。
灯光防控技术主要以昆虫趋光性或负趋光性为理论基础开发,对果园夜行性蛾类进行诱捕、驱避或干扰以达到绿色防控目的[12-14]。目前,该项技术在中国主要荔枝和龙眼产区被推广应用,其相关研究主要针对荔枝、龙眼果期头号害虫——荔枝蒂蛀虫,对于梢期头号害虫尺蛾的影响却无研究报道[13-16]。笔者以粗胫翠尺蛾为试验对象,在该虫转录组数据结果基础上鉴定2个视蛋白基因,并对其全长进行克隆;在对其序列特征进行生物信息学分析后,通过RT-qPCR技术测定2个视蛋白基因(TiLW和TiUV基因)的发育模式以及在光照处理后的表达变化,以期为阐明视蛋白在粗胫翠尺蛾应激反应中的生理功能研究提供线索。
1 材料与方法
1.1 试验材料与试剂
粗胫翠尺蛾采自广东省农业科学院白云试验基地荔枝园,置于植物保护研究所实验室内长期饲养。饲养条件:温度(25±1)℃,相对湿度(60±5)%,光周期14:10(L:D)。幼虫以荔枝嫰梢喂养,成虫以10%的蜂蜜水为食。
1.2 总RNA的提取与反转录
利用Trizol试剂分别提取粗胫翠尺蛾成虫头部的总RNA,采用2%的琼脂糖凝胶电泳进行检测,再采用PrimeScript RT Reagent试剂盒(Takara,Tokyo,Japan)反转录合成cDNA第一链,于-20℃中保存备用。
1.3 序列克隆与分析
以粗胫翠尺蛾转录组(数据待发表)测序结果为基础,从成功注释的unigene中搜索视蛋白(opsin)基因序列,将其对应的蛋白序列与NCBI中的nr数据库进行BLASTP,验证是否属于视蛋白。利用Primer 5.0软件根据目的基因片段设计引物,用于获得的粗胫翠尺蛾TiUV、TiLW视蛋白基因全长序列(表1)。分别以1对特异性引物进行5’和3’RACE扩增。5’RACE反应程序为:94℃ 3 min;94℃ 30 s,65℃ 30 s,72℃ 1 min,32个循环;72℃10 min;4℃保存。3’RACE反应程序为:94℃ 3 min;94℃ 30 s,58℃ 30 s,72℃ 1 min,32个循环;72℃10 min;4℃保存。最后用DNAstar软件将所得序列片段拼接为完整的序列。
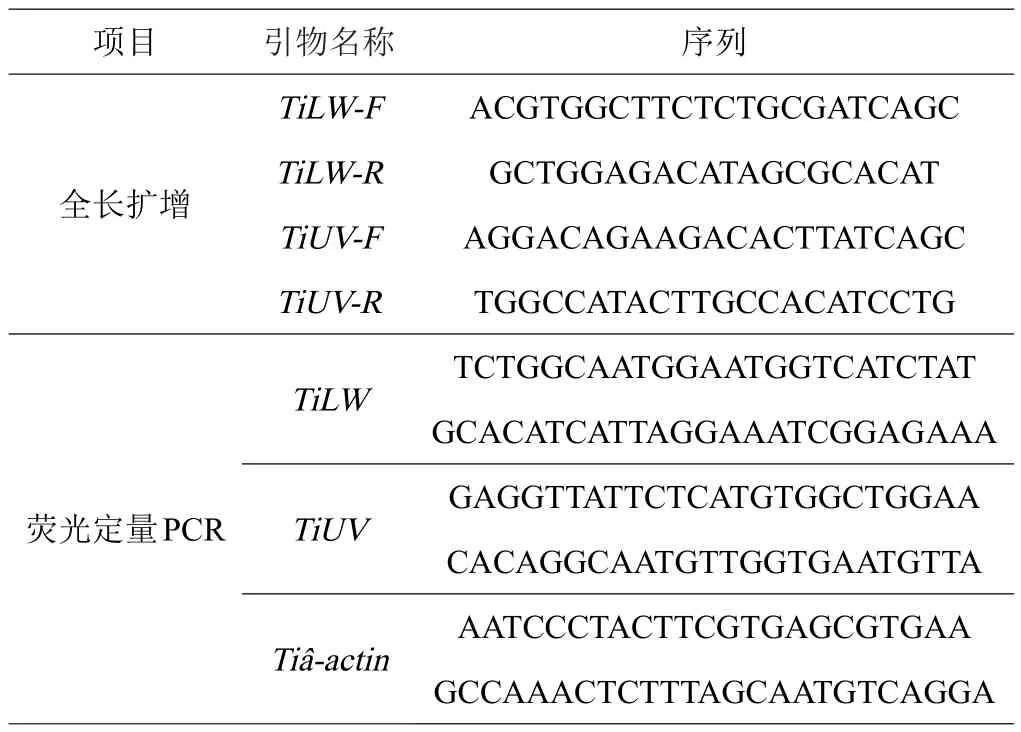
表1 试验所用的引物
使用NCBI中的BLAST工具进行核酸序列的同源性分析,与不同物种进行比对,构建系统发育树。使用在线软件ORF Finder查找序列的开放阅读框,使用DNAstar软件预测其编码氨基酸序列。利用ProtParam软件预测蛋白的分子量、等电点等理化性质;ProtScale软件进行疏水性分析;TMHMM 2.0软件分析跨膜结构;SOPMA在线软件预测视蛋白二级结构,Prosite Scan和Motif Scan在线软件分析其氨基酸序列的功能位点。采用MEGA 4.1软件中的邻位相连法(neighbor-joining,NJ)构建系统进化树,Bootrap为1000次。
1.4 光照处理
在不同人工气候箱中(GXZ型,宁波江南仪器厂)分别设置24L:0D(光照处理组)和14L:10D(对照组),将当天羽化的粗胫翠尺蛾成虫共8头(♀:♂=1:1)置于饲养盒中,将饲养盒置于不同处理的人工气候箱中,用浸润10%蜂蜜水的棉球进行饲养,分别于处理后24 h和48 h取样。
1.5 荧光定量检测
利用Primer 5.0软件设计粗胫翠尺蛾的TiUV和TiLW基因的qPCR引物(表1)。采用GoTaq qPCR Master Mix试剂盒(promega,麦迪逊,美国)进行RT-qPCR。其反应体系包括:10 μL GoTaq qPCR Master Mix,1 μL cDNA模板,0.5 μL正向引物,0.5 μL反向引物,8 μL DEPC水。反应程序:95℃预变性2 min,95℃变性15 s,60℃退火60 s,40个循环。以Tiβ-actin基因为内参基因计算相对表达量,数据分析使用比较比较CT值方法分析[17]。每次RT-qPCR需要3次技术重复,并且每个样本进行3个生物学重复。
2 结果与分析
2.1 粗胫翠尺蛾视蛋白基因的克隆与序列分析
以粗胫翠尺蛾成虫头部cDNA为模板进行PCR扩增,得到TiLW和TiUV视蛋白基因的ORF长度为1504 bp和1762 bp;分别编码为374个和378个氨基酸,蛋白分子量分另为41.04 kDa和41.52 kDa,等电点为7.01和5.69。序列二级结构分析结果表明,TiLW和TiUV蛋白序列主要由α-螺旋和随机卷曲组成。其中,TiLW氨基酸序列中α-螺旋比例占45.01%,β转角比例占2.70%,随机卷曲比例占33.15%,延伸链比例占19.14%;TiUV的α-螺旋比例占2.93%,其序列β转角、随机卷曲和延伸链比例均高于TiLW(表2)。与其他已报道昆虫视蛋白一样,TiLW和TiUV均具有7个跨膜螺旋区。TiLW包含4个N-糖基化位点,4个酪蛋白激酶II磷酸化位点,4个N-豆蔻酰化位点,2个蛋白激酶C磷酸化位点,1个G蛋白耦合受体家庭标签,1个视网膜结合位点;TiUV包含1个N-糖基化位点,9个酪蛋白激酶II磷酸化位点,8个N-豆蔻酰化位点,1个蛋白激酶C磷酸化位点(图1)。
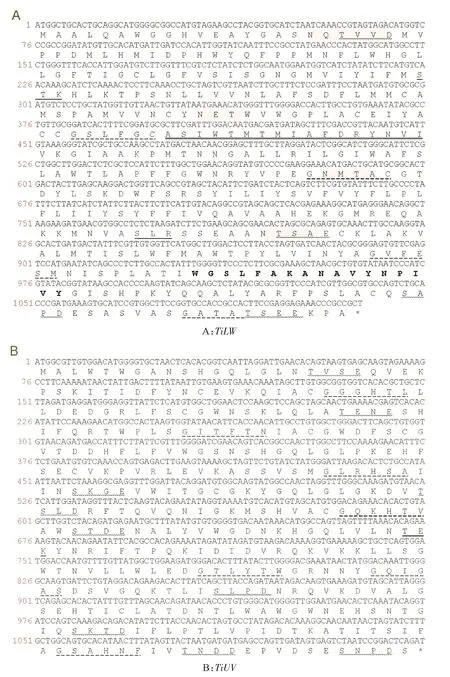
图1 粗胫翠尺蛾TiLW和TiUV视蛋白氨基酸序列
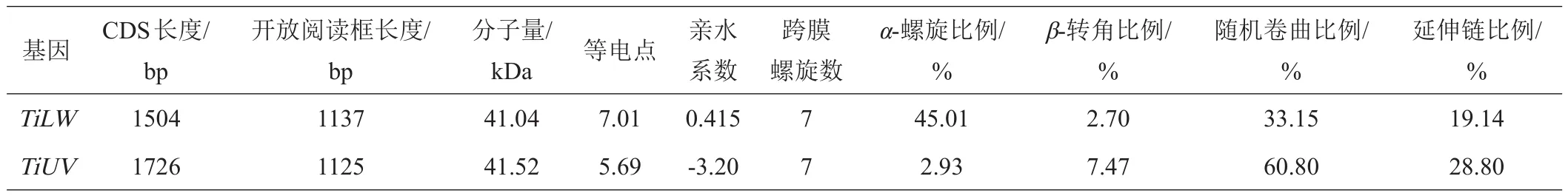
表2 粗胫翠尺蛾2个视蛋白基因及其编码氨基酸的序列特征
2.2 基于UV和LW视蛋白氨基酸序列进化树的构建与同源性分析
通过与其他昆虫的同类蛋白氨基酸序列进行比对,粗胫翠尺蛾TiLW和TiUV与其他物种的长波长视蛋白LW和紫外光视蛋白UV具有较高同源性。使用MEGA 5软件邻接法进行1000次重复计算后构建进化树(图2)。进化关系分析表明,克隆得到的粗胫翠尺蛾视蛋白分属于形成LW和UV 2个明显的分支。其中,粗胫翠尺蛾TiLW同源性最高为桦尺蛾(Biston betularia)和松白条尺蠖(Operophtera brumata)同类蛋白,与TiUV同源性最高的有大蜡螟(Gallearia mellonella)、亚洲玉米螟(Ostrinia fumacalls)和柑橘凤蝶(Paplllo xuthus)等多种鳞翅目昆虫UV蛋白。
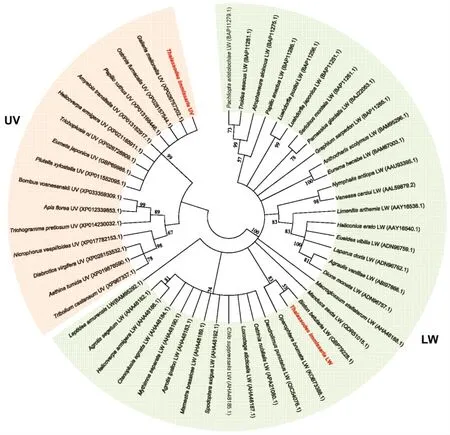
图2 粗胫翠尺蛾与其他昆虫的视蛋白系统进化树分析
2.3 TiLW和TiUV在粗胫翠尺蛾不同发育阶段及雌雄成虫期表达量分析
对TiLW和TiUV在粗胫翠尺蛾不同发育阶段表达量进行分析。结果表明,TiLW和TiUV在粗胫翠尺蛾不同发育阶段均有所表达(图3)。TiLW在3日龄雌成虫有最高量表达,其次为2日龄雌成虫;TiUV同样在2、3日龄雌、雄成虫体内有高量表达,其次为卵期。TiLW和TiUV视蛋白基因在粗胫翠尺蛾成虫期表达量普遍高于其他发育阶段,而且从1日龄到3日龄雌雄成虫体内的基因表达量呈现逐步升高趋势。比较而言,TiUV在卵期高于TiLW;在幼虫阶段低于TiLW,仅在4、5龄期幼虫体内表达与TiLW同期表达无显著差异;在蛹期和雌性成虫体内,TiUV表达量远低于TiLW,但在雄性成虫体内两者表达量无显著性差异。推测TiUV在虫体卵期扮演重要角色,TiLW在幼虫和成虫期有更重要的生理功能,TiLW在雌性成虫视觉功能中居主导地位。
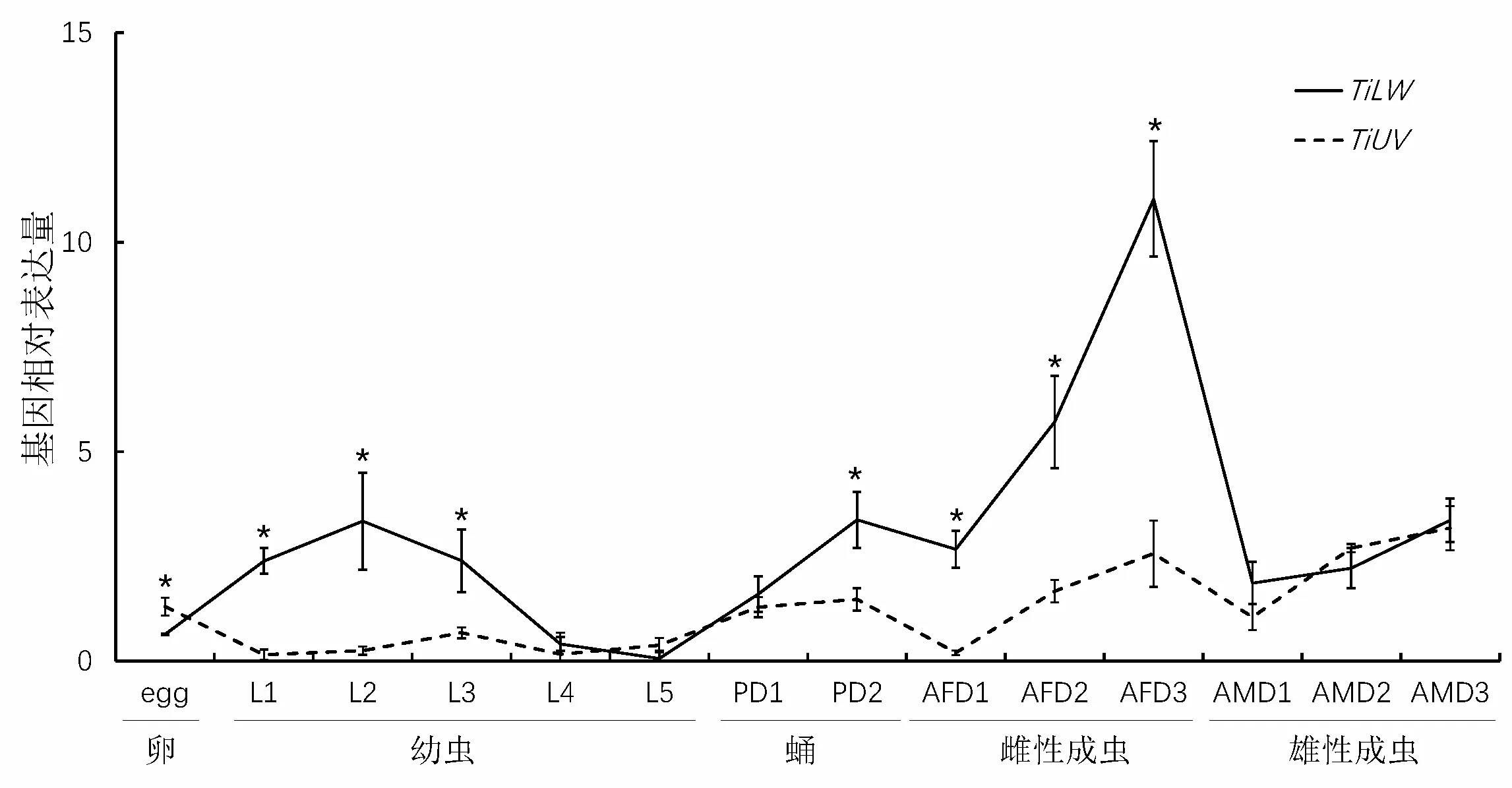
图3 TiLW 和TiUV 视蛋白基因在粗胫翠尺蛾雌雄成虫期的表达量
2.4 光照处理对粗胫翠尺蛾TiLW和TiUV视蛋白基因表达量影响
对光照处理组和对照组粗胫翠尺蛾头部TiLW和TiUV视蛋白基因表达量变化进行RT-qPCR检测。图4结果显示,成虫头部TiLW和TiUV的表达量在光照处理后均受到不同程度的影响。与对照组相比,光照处理后24 h,TiLW在雌、雄虫头部分别上升12.96倍和降低8.61倍,而TiUV仅在雄虫头部降低3.10倍;光照处理48 h后,仅TiLW在雌虫头部表达量与对照组无差异,在雄虫头部表达量下降3倍,而TiUV在雌虫头部下降1.84倍,在雄虫头部表达无差异。整体而言,光照处理24 h和48 h对粗胫翠尺蛾TiLW的表达影响要大于TiUV基因。

图4 光照处理24 h与48 h对TiLW和TiUV视蛋白基因表达量的影响
3 讨论
灯光干扰技术是一项重要的荔枝龙眼害虫绿色防控措施,具有高效环保、无农药残留危害、对害虫不产生抗药性等特点[14-16]。光对昆虫影响有多种,包括趋光性、光适应、干扰昼夜节节律、影响生长发育等。灯光干扰技术则以上述靶标害虫的影响为基础开发[12-13]。目前,该项技术在广西和广东无公害荔枝龙眼种植园广泛推广应用,室内试验结果表明,24L:0D光周期处理后,荔枝蒂蛀虫单雌产卵量由136.33粒降低至1粒;田间试验验证夜晚光照下荔枝蒂蛀虫蛀果率可控制在4.00%以下[16]。由此可见,该项技术在荔枝龙眼农药减量化种植中有重要推广意义。但是该项技术对于荔枝、龙眼梢期头号害虫——粗胫翠尺蛾的影响却并无研究报道。本研究首次从粗胫翠尺蛾中克隆得到了2种视蛋白基因(TiLW和TiUV),在对其序列特征进行生物信息学分析后,通过RT-qPCR技术测定了2个视蛋白基因的发育模式以及在光照处理后的表达变化,以期为阐明灯光干扰技术对非靶标害虫影响提供线索,同时为视蛋白在粗胫翠尺蛾应激反应中的生理功能研究提供参考。
鳞翅目昆虫视觉系统高度发达,其视觉主要依赖于对不同光谱有吸收性的光感细胞,光感细胞对不同光谱的吸收主要由视蛋白和发色团结合而成的视色素决定[11,18]。光周期、日龄、性别等因素都对昆虫视蛋白基因的表达有一定影响,而且不同的视蛋白基因表达的影响因素不尽相同[9]。本研究所获得的粗胫翠尺蛾TiLW和TiUV视蛋白,其编码氨基酸经序列分析显示,两者具有昆虫同类蛋白所具有的多个保守结构域,包括N-糖基化位点、酪蛋白激酶II磷酸化位点、N-豆蔻酰化位点和蛋白激酶C磷酸化位点等,但是仅在TiLW中找到了G蛋白耦合受体家庭标签和视网膜结合位点。这些保守位点对于视蛋白结构形成及与发色团结合具有重要作用[9,18-19]。此外,基因表达时期和组织特点在一定程度上反映基因功能。对马尾松毛虫(Dendrolimus punctatus)的研究表明,LW和BL视蛋白基因在成虫期高量表达,UV则在卵和幼虫期高量表达[20];柑橘木虱(Asian citrus)雌性成虫中LW表达量显著高于UV,而雄性成虫中LW表达量略低于UV[21];棉铃虫(Helicoverpa armigera)雌、雄性成虫中LW表达量均高于UV[22]。UV和LW视蛋白在不同昆虫以及昆虫不同发育时期的视觉刺激感知过程中的生理作用均有所区别。本研究中粗胫翠尺蛾TiUV在卵和成虫期有较高量表达,而TiLW在幼虫期和成虫期有较高量表达,TiLW在雌性成虫的表达量远高于雄性成虫,推测TiUV可能与卵孵化相关,TiLW可能与幼虫觅食和雌性成虫产卵地点选择更为相关。
昆虫头部和复眼是昆虫感知光刺激的关键部位,其头部视蛋白基因的表达与昆虫生存环境息息相关[23-25]。因此通过克隆并研究视蛋白结构的组成,进一步了解视蛋白如何参与调控昆虫感知周围环境的光信号,对害虫视蛋白与环境相适应的机制研究非常重要。在本研究中,为研究光照对粗胫翠尺蛾视蛋白基因表达量的影响,使用成虫头部组织作为试验样品。前期研究发现在持续黑暗条件下棉铃虫3个视蛋白基因的相对表达量显著性上调[22],不同(颜色或强度)光源下,粘虫(Mythimna separata)、赤拟谷盗(Tribolium castanem)和韭菜迟眼蕈蚊(Bradysia odoriphaga)等视蛋白基因的表达均受到不同程度的影响[26-28]。本研究发现,持续光照处理后TiLW的表达变化要大过TiUV,在光照处理24 h后,TiLW在雌雄成虫体中分别显著上升12.96倍和下降8.61倍,TiUV仅在雄性成虫中下降3.10倍;而在光照处理后48 h,粗胫翠尺蛾TiLW和TiUV表达量与对照组差别不大。结果表明参与粗胫翠尺蛾在光照刺激下的应激过程,光刺激可引起粗胫翠尺蛾2个视蛋白基因表达变化,但在处理一定时间后该虫可适应光刺激,即灯光干扰技术对粗胫翠尺蛾视蛋白表达影响并不大。粗胫翠尺蛾依靠视觉和嗅觉信号寻找寄语、配偶、栖息地和产卵场所,但光感受器和嗅觉感受器如何协调作用调节这些生物行为的机理尚不清楚。粗胫翠尺蛾如何通过调节视蛋白基因的表达来感知和适应其周围环境、是否参与调控该虫的其他生物习性、光干扰技术对粗胫翠尺蛾繁殖力影响等,均值得进一步深入研究。
