太行山东坡中段植物多样性垂直分布格局及其驱动因素*
2022-07-08梁红柱刘丽丽付同刚朱建佳刘金铜
梁红柱,刘丽丽,高 会,付同刚,朱建佳,苏 杨,刘金铜**
(1.河北师范大学生命科学学院 石家庄 050024; 2.中国科学院遗传与发育生物学研究所农业资源研究中心 石家庄 050022; 3.中国科学院大学 北京 100049; 4.河北省国土整治中心 石家庄 050031; 5.河北科技师范学院园艺科技学院 秦皇岛 066004)
山地是指具有一定海拔、坡度和相对高度的地面,反映且浓缩了水平自然带的自然地理和生态学特征,生境高度异质化。由于较少的人类干扰,山地成为陆地生物物种的栖息地与避难所,是地球上生物多样性最为丰富的单元,全球生物多样性保护的重点区域。山地对气候变化反应极为敏感,山地生态系统随全球气候变化响应的研究,已成为全球变化研究的热点领域。植被在地球上的分布呈明显的地带性变化,表现为水平地带性和垂直地带性,而垂直地带性包含了多重环境因素的梯度效应,气候、植被、土壤和水文等自然要素随海拔自下而上逐渐更替。山地景观在海拔梯度上的变化是水平梯度的1000倍,被认为是水平地带性变化的“微缩模型”。山地植物多样性的空间格局是群落生态学研究的核心内容,有助于了解生物多样性的维持机制,而山地植被的垂直变化也一直是生态学的热点领域。
对山地植被垂直带谱的研究历史已久,但较之自然地理学的其他领域仍有很大差距。研究山地植物群落物种多样性的垂直分异格局,有助于理解海拔梯度上的植被地带性变化。植物多样性随海拔梯度的变化呈现多种趋势格局。在山地群落多样性垂直格局研究中,α多样性指数、β多样性指数和γ多样性指数的应用较为广泛,而β多样性更能体现环境梯度下的群落组成和分化程度,且对生物多样性保护决策具有重要的指导意义。近年来数量分类学方法和排序法受到广泛重视,而计算机技术的快速发展,促生了一系列数据处理和分析方法的创新和推广。生物多样性与海拔梯度格局的关系,依赖于环境变量之间的协作关系,而不同地理区域的山地植物群落垂直变化格局是不同的,且形成这种格局的机制尚不明确。
太行山为我国第二、第三阶梯分界线,位于黄土高原和华北平原的过渡地带,是京津冀经济圈和华北平原的天然屏障和重要水源地。太行山的植被历来受到人为干扰,近20年以来,太行山植被状况呈好转趋势。由于地形、土壤水分、光辐射等差异,太行山东西坡植被差异较大,东坡的垂直变化更为突出。对太行山植被分布的研究,主要集中于植物群落的组成、构建与演替,地形因子、人为干扰、气候因素等对植被的影响以及太行山区生态系统服务功能格局等。依据垂直梯度上的生态要素特征,太行山地地貌类型可分为低山丘陵区、中山区和亚高山区,不同生态类型区具有特定的植被分布格局和生态系统服务功能。根据山地海拔不同,植被类型由低到高大致可分为低山落叶阔叶灌丛带、中山落叶阔叶林带和亚高山草甸带。对太行山区垂直梯度的群落构建和演替机制尚无定论,地形、气候和土壤等因素对植物群落垂直分布的综合影响,以及物种分布对气候变化的响应也缺乏深入的研究。
本文基于太行山东坡中段海拔梯度上的群落样方调查,综合各项环境因素,运用群落生态学和数量分类学的研究方法,探讨太行山东坡中段垂直梯度植物多样性的分布格局,了解群落演替的特征,分析影响物种分布的主导因素,解释植被和环境之间隐含的内在关联,旨在为山地生物多样性的保护和可持续利用提供理论支持,为脆弱山地生态系统的恢复重建提供科学依据。
1 研究区概况
研究区太行山区位于我国第二阶梯东缘,东西衔接黄土高原与华北平原,总面积为13.7万km。调查取样点位于太行山东坡的中段,地理坐标为113°41′43″~113°53′35″E,38°33′13″~38°45′48.7″N。地势由北向南倾斜,该区最高点为驼梁国家级自然保护区的主峰南驼,海拔为2281 m,为河北省第三高峰。
该区域地处暖温带,属半湿润半干旱大陆性季风气候,四季分明,雨热同季,蒸发量大,昼夜温差较大。年均温8.0 ℃,年日照率达62%,年均太阳辐射总量131~136 kJ·cm。年平均降雨量为690 mm,年均蒸发量为1882.6 mm。该区地貌类型较单一,主要土壤类型为棕壤、褐土、草甸土和亚高山草甸土。
2 研究方法
2.1 样地设置
本研究于2018年6月至2020年10月,以太行山东坡中段地区海拔梯度上的植物群落为对象,自低海拔(100 m)到高海拔(2282 m),每升高100 m设一个取样位点(图1),每一取样位点平行设置5套样方作为重复,每套样方包含6个小样方:包括乔木样方1个,投影面积为10 m × 10 m,内设5 m × 5 m的灌木样方4个,1 m × 1 m的草本样方1个,总共16个取样位点。共包含乔木样方80个,灌木样方320个,草本样方80个,共计480个样方。记录每个样方内的植物种类及株数,并用GPS定位样方的经纬度、海拔等信息。5套样方调查到的乔木、灌木和草本物种总数作为该取样位点的物种丰富度; 5套样方乔木、灌木和草本植株个体的平均密度,作为该取样位点不同类型群落的植株密度。

图1 太行山东坡中段研究区域取样位点图Fig.1 Sampling sites map of the study area in the middle section of the eastern slope of the Taihang Mountain
2.2 环境因子数据获取
气候因子:基于太行山区的气象站点近20年平均气温和降水数据,在ArcGIS中采用克里格插值法,分别给每个样方赋值气候因子数值。
地形因子:基于DEM影像,提取样地的平均坡度和坡向等数据。
土壤因子:取调查样地0~20 cm的表层土样,测定其C、N、P、K和有机质等含量,并测定土壤孔隙度等指标。
2.3 植物群落多样性测度
采取多样性测度的方法计算群落α多样性和β多样性指数,其中群落内生物多样性(α多样性)主要包括:丰富度指数、Shannon-Wiener指数; 群落间生物多样性(β多样性)主要包括:Sørensen指数、Jaccard指数和Cody指数。多样性指数计算公式如下:
丰富度指数()为样方内物种数。

式中:为物种数目,P为种的个体数在全部个体数中的比例。

式中:、为两个群落的物种数,为两个群落共有的物种数,()为沿生境梯度增加的物种数,()为沿生境梯度失去的物种数。
Sørensen指数和Jaccard指数反映群落或样方间物种的相似性,Cody指数反映样方物种组成沿环境梯度的替代速率。
2.4 排序分析
依据样方调查物种类型和物种密度数据,建立海拔梯度上植物群落优势物种丰富度矩阵,并建立对应的环境变量矩阵(包括气候因素、地形因子和土壤因子)。基于R语言的“vegan”软件包,采取RDA和CCA排序方法,生成排序图。
2.5 数据处理
本研究基于太行山区遥感影像数据,用ArcGIS 10.2提取相应环境因子,采用Origin 9.3和R4.0.3软件对数据进行处理和作图; 为了解环境因子对物种分布的影响,采取R4.0.3软件对物种和环境因子进行排序分析,探讨影响太行山中段维管植物群落构建和物种分布的主导因素。
3 结果与分析
3.1 植物多样性的垂直分异特征
3.1.1 物种丰富度垂直分异特征
太行山区中段地带性植被以暖温带落叶阔叶林为主,由于总体海拔不足够高,尽管没有形成明显的植被垂直带谱,植物群落的结构和功能在海拔梯度上也存在较大差异。不同物种及其个体多度是形成差异的基础,因此研究植物群落在海拔梯度上的结构和多样性分布具有重要意义。依据样方群落种群类别和密度,结合乔木、灌木和草本物种的层高和盖度,计算其重要值,筛选出海拔梯度上群落优势物种:胡桃()、荆条(var.)、酸枣(var.)、狭叶珍珠菜()、杨(var.)、地榆()、油松()、臭椿()、茜草()、旋覆花()、白蔹()、风毛菊()、委陵菜()、唐松草(var.)、绣线菊()和白莲蒿()。
由图2a可以看出,随山地海拔升高,物种丰富度和植株密度出现明显波动。在海拔100~900 m,物种丰富度基本呈逐渐升高趋势,海拔900 m处达到最大值; 900~1100 m物种数急剧降低,1100 m达谷底;之后到海拔1800 m之间,物种丰富度再次逐渐增加,1800 m以上物种再次减少。从物种丰富度来看,与太行山北段的植物群落单峰分布的结论不同,太行山中段群落物种多样性在海拔梯度上的分布更为复杂,但基本符合物种多样性垂直分布的“中间高度膨胀”理论。

图2 太行山东坡中段维管植物群落丰富度和种群密度(a)、α多样性(Shannon-Wiener指数)(b)、相似性指数(Sørensen和Jaccard指数)(c)和群落更替速率(Cody指数)(d)的垂直分布Fig.2 Altitudinal variations of the species richness and density (a),alpha index (Shannon-Wiener index)(b),similarity index(Sørensen and Jaccard indexes)(c)and succession rate (Cody index)(d)of vascular plant communities in the middle section fo the eastern slope of the Taihang Mountain
3.1.2 群落α多样性垂直分异特征
物种丰富度和群落α多样性(Shannon-Wiener指数)在海拔梯度上的变化趋势,常被作为物种多样性的垂直分布格局。由图2b可以看出,海拔梯度上群落α多样性与物种丰富度的分布具有相似的趋势:随海拔升高,群落α多样性先后在海拔900 m和1800 m左右出现峰值,表明植物群落在这两个区段具有较丰富的物种多样性。综合图1和图2的结果,太行山东坡中段物种多样性在海拔梯度上的分布,出现了不同于太行山北段地区的单峰格局,呈现较为复杂的两次峰值的模式。
3.1.3 植物群落β多样性垂直分异特征
植物群落的β多样性常用来描述植物群落间的演替特征和动态变化,而相似性指数(Sørensen指数和Jaccard指数)则反映了群落间物种组成的相似性。如图2c所示:随海拔升高,Sørensen指数和Jaccard指数呈现基本一致的变化规律。在海拔700~1200 m和1700~2000 m,相似性指数出现两个较低区间,表明这两个区间植物群落之间相似性较低,群落间差异显著,而反映出物种多样性更为丰富,这与图1和图2的结果相统一。
植物群落β多样性的测度也可用Cody指数来反映,通过比较群落间新增加和失去物种的数量,更为直观地了解物种更替状况,可反映群落物种组成沿环境梯度的替代速率。图2d表明,Cody指数的峰值出现在海拔700~1200 m和1700~2000 m,意味着这两个区间植物群落物种更替速率更快,群落物种组成差异增大,物种多样性程度相应升高。本结果与朱珣之等最大物种丰富度出现在海拔1500 m不同,可能与太行山东坡中段海拔梯度的水热分布有关。
3.2 植物多样性垂直分布格局与生物环境关系分析
3.2.1 植物群落多样性与生物环境因素的关系
植物群落多样性受海拔、温度、降水、植被净初级生产力(NPP)等生物与环境因素的影响。图3显示,太行山东坡中段随海拔梯度升高,温度持续降低,而降水升高。植被净初级生产力(NPP)随海拔梯度升高,出现与Shannon-Wiener指数类似的分布规律,分别在700~900 m和1700~1800 m出现两次峰值,这与太行山区产水量的垂直分布具有类似的规律。
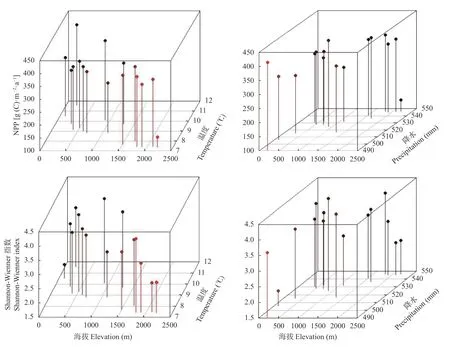
图3 太行山东坡中段植被净初级生产力(NPP)、Shannon-Wiener指数与海拔、温度和降水之间的关系Fig.3 Relationships between vegetation net primary productiviyt (NPP),Shannon-Wiener index and elevation,temperature and precipitation in the middle section of the eastern slope of the Taihang Mountain
对植物群落Shannon-Wiener指数与NPP及环境因子进行偏相关分析,发现Shannon-Wiener指数与海拔呈负相关,与NPP和温度呈正相关,但均未达到显著水平; 与降水相关性不明显(表1)。可见,群落物种多样性与植被NPP在海拔梯度上具有一致的变化趋势,二者之间具有显著相关性。

表1 太行山东坡中段植被物种丰富度、Shannon-Wiener指数与环境因子偏相关分析Table 1 Partial correlation analysis between species richness,Shannon-Wiener index of vegetagtion and environmental factors in the middle section of the eastern slope of the Taihang Mountain
3.2.2 植物多样性垂直分布格局的环境解释
基于海拔梯度上的样方调查,筛选出不同群落的优势物种,应用数量生态学方法,对太行山东坡中段物种多样性海拔梯度的分布与环境因素之间的关系进行了研究。用R-vegan程序包对物种分布数据进行排序分析,发现物种分布的“axis lengths”介于3与4之间,因此可选用线性模型或单峰模型进行排序分析。本研究选用线性模型(RDA)和单峰模型(CCA)排序方法分别对数据进行排序,结果如图4所示。
由图4可以看出,RDA排序显示影响物种垂直分布相关性较强的因素有海拔、温度、降水、土壤孔隙度等; 物种分布中,胡桃、荆条、酸枣和杨具有相似的分布环境; 绣线菊、风毛菊、唐松草、委陵菜等分布环境类似。环境因素中,海拔与物种分布最为密切,且对物种分布的解释量(影响程度)也较大; 其次,温度和降水对物种分布的解释量也较大,相关性弱于海拔的影响。CCA排序表明对物种分布影响程度较大且相关性高的环境因素包括降水、温度和海拔。土壤K含量和坡度对物种分布的影响程度最小。其中胡桃、荆条、杨等分布环境相似,风毛菊、绣线菊、唐松草等分布环境相似。CCA排序结果与RDA结果类似,印证了线性模型和单峰模型均适用于该区域物种分布的排序研究。

图4 太行山东坡中段植物群落优势物种RDA和CCA排序图Fig.4 RDA ordination and CCA ordination of the dominant plant species in the middle section of the eastern slope of the Taihang Mountain
用R软件的permutest()函数,对CCA排序结果进行蒙特卡洛置换检验(Monte Carlo permutation test),分析10个环境因子对物种分布的解释量是否具有显著性。结果显示显著性为0.016,达到显著性水平,说明排序结果可以接受多种环境因子对物种分布的综合解释量。应用envfit()函数对每种环境因子进行检验,结果如表2所示。其中CCA1和CCA2代表环境因子与排序轴夹角的余弦值,表示环境因子与排序轴的相关性。代表环境因子对物种分布的决定系数,其值越大,代表该环境因子对物种分布的影响越大。Pr表示相关性的显著性检验。

表2 太行山中段植物群落垂直分布影响因子CCA排序结果蒙特卡洛显著性检验Table 2 Monte Carlo significance test of the CCA ordination results of the influencing factors to the altitudinal distribution of plant communites in the middle section of the eastern slope of the Taihang Mountain
从检验结果看,海拔、降水和温度对物种的分布影响最为显著,土壤因子中的C、N和土壤孔隙度对物种分布的影响也较大。地形因子对太行山东坡中段群落物种分布的影响不显著。数量分类学排序结果与群落多样性分析结果一致,说明分类结果具有合理性。α多样性和β多样性分布格局与排序结果具有一致性,也印证了排序分类结果的合理性。
4 讨论
4.1 太行山东坡维管植物物种多样性垂直分布格局
根据样方调查和标本采集数据,太行山东坡中段低山丘陵区的维管植物共计54种,隶属于32科49属; 中山区103种,隶属于47科88属; 亚高山区58种,隶属于21科48属。海拔梯度上各植被带的调查取样面积一致,因此上述数据可反映物种多样性的总体大小。由调查结果可以看出,物种多样性在中海拔地区明显偏高,这与Mabessimo对太行山北段东灵山植物多样性的研究结果一致,符合山地植被垂直分布“中间高度膨胀”理论。
张金屯等和茹文明等对中条山植被垂直带的数量分类研究,认为有序样方聚类方法的结果更符合实际。本研究采取海拔梯度上有序样方调查进行数据获取,从结果来看,在低山丘陵区,随着海拔梯度上升,α多样性指数呈现持续升高的趋势。结合太行山区产水量垂直分异特征,可知在低山区海拔500 m处,产水量出现峰值,表明在此区间,生态系统结构和功能匹配较好,植物群落物种丰富度也呈现持续增加的趋势。对整个太行山区NPP垂直分异特征的研究表明,NPP年际间变化速率在海拔600 m处也出现峰值,这也印证了在此区间物种丰富度较高的结论。海拔600 m以下,物种丰富度和种群密度相对较低,而对太行山区植被指数NDVI的研究发现,太行山东部中低海拔区域的丘陵地带,NDVI出现减少趋势,可能均与该区段山地坡度较小,人类活动频繁,人为干扰较重有关。
太行山东坡中段的中山区,维管植物丰富度呈先降后升的趋势,在中部海拔达峰值,这与郑敬刚等对太行山南段王屋山的物种垂直分布特征一致。而对太行山区植被NPP和产水量垂直分异特征的研究显示,植被NPP和产水量分别在海拔1300 m和1400 m再次出现峰值,表明在此海拔区段,太行山区生态系统服务功能与植被再次达到较好的匹配,加之人为干扰程度下降,植被丰富度也随之升高而再次出现峰值。由于太行山中段地区总体海拔不高,随海拔升高,温度下降幅度不大,在产水充足的情况下,水热匹配尚能达到较好的状况。因此,随海拔升高,产水量在低山区和中山区出现两次峰值的情况下,山地植被丰富度可跨越产水峰值区,在一定程度上呈现持续增加的趋势。而到了亚高山区段,随着产水和温度的同步下降,群落丰富度再次持续降低。
亚高山草甸区段,植被以温带性质的草本植物为主,一年生草本植物比例较高,特有种较丰富,表明仍受到一定程度的人为干扰。海拔1800 m以上物种丰富度下降明显,而群落相似性指数则急剧上升,表明亚高山区维管植物群落的共有种较多,物种多样性则较低; 该区段Cody指数也明显持续降低,表明群落间物种的更替速率下降,群落维管植物物种趋于稳定。
4.2 太行山东坡海拔梯度上植物群落差异及对演替的影响
生物多样性的维持机制历来是生态学的重要研究内容,群落中物种的共存对生物多样性的维持至关重要,然而对群落的构建和演替机制尚不明确。当前,群落构建和物种共存的理论仍以生态位理论为主,而基于生态位模型(ecological niche modeling)的生物多样性维持机制研究受到了广泛关注。对群落物种间的联结关系进行分析,常被用来揭示种间关联性和了解生境因素对植被演替的影响。在群落生境的空间异质性加大时,物种多样性则趋于升高。
β多样性指不同群落间物种组成的差异,由物种更替和嵌套两个过程所决定,可较为直观地反映环境梯度上植物群落物种组成的差异和梯度变化。相比较于α多样性和γ多样性,β多样性随海拔梯度变化的研究相对较弱。在β多样性研究的过程中,众多测度方法被提出,其中加性分配法则、相似性指数和相异性指数被广泛采用。本研究主要讨论Sørensen、Jaccard和Cody指数的海拔梯度变化,发现太行山东坡中段随海拔升高,植物群落β多样性指数的垂直变化不同于常见的单峰模式,其群落物种多样性在中部海拔区间出现两次峰值,而β多样性指数也分别在峰值区相应出现低谷值,表明群落间的物种相似度较低,反映出较高的物种丰富度;Cody指数也相应出现两个峰值,表明群落间物种更替速率在此区间较快,从而使群落间物种相似程度降低。中部海拔物种丰富度出现两次峰值,表明低山区和亚高山区的物种在中部海拔地区发生交汇,物种周转较快,从而形成较丰富的物种多样性。
β多样性加性分解的应用,可以更好地揭示海拔梯度上的群落构建机制。通过对太行山东坡中段群落β多样性的分析可知,在海拔梯度上β多样性变化活跃区段,物种周转组分对形成物种丰富度的贡献更大。植物群落的演替与其生境具有协同演替效应,这与太行山南段油松群落的分布类似。综合研究结果,太行山东坡中段植物群落的垂直分布格局和物种多样性水平,是受到群落垂直演替进程、环境因素和人为干扰协同作用的结果。
4.3 植物群落垂直分布格局与环境因素的关系
区域尺度上,影响生物多样性梯度变化的因素包括:水分、光照、温度、土壤养分、土壤pH、干扰、竞争、捕食等多种因素。本研究主要分析环境因素中的气候因子、地形因子和土壤因子对海拔梯度上植物群落优势物种分布的影响。对整个太行山区的植被指数研究发现,16%的植被NPP显著升高,而气候因素对NPP起到促进作用,人为干扰表现为抑制作用。
曹杨等对太行山南段植被群落的数量生态学研究表明,群落与优势种的分布与山地坡度和坡向相关性较大,认为CCA排序方法提供了物种与环境的相关性,更适合于该区域的群落分布研究。本研究中,RDA和CCA排序结果显示,群落优势种的垂直分布与海拔和温度相关性最大,降水的影响稍弱。地形因子中坡度和坡面对该区域物种垂直分布的影响均达显著水平。土壤因子中,土壤全氮含量、全碳含量以及土壤孔隙度与物种垂直分布的相关性较大,土壤钾含量和磷含量相关性未达显著水平。坡度可增加太行山区植被带谱结构组成的复杂性,而本研究坡度对群落垂直分布格局的影响不显著。海拔、水和热是影响太行山东坡中段地区群落物种垂直分布的主要因素。
采用线性模型和单峰模型对太行山东坡中段地区群落优势种排序分析,显示了较为一致的结果。低海拔地区,酸枣、荆条、胡桃等物种分布于同一象限,表明具有相近的生存环境,且与温度呈显著正相关,而与海拔和降水呈显著负相关; 绣线菊、风毛菊、唐松草、委陵菜等物种具有相似的分布环境,与海拔、降水等环境因子呈显著正相关,而与温度呈显著负相关。由CCA显著性检验结果印证了相似的结论,即海拔和水热条件是驱动该区域物种垂直分布的主要环境因素。
5 结论
由研究结果可知,太行山东坡中段维管植物群落α多样性垂直分布出现两次峰值,符合“中部高度膨胀”理论,这一分布格局与太行山区产水量和NPP垂直格局一致; 植物群落β多样性相似性指数在物种丰富度峰值区达到最低,表明群落间差异性最大,群落间物种演替速率增大,因此群落物种多样性增加; 根据RDA和CCA排序结果可知,影响植物群落垂直分布的主要环境因素是海拔、温度和降水,而与坡度、坡面等地形因子相关性不显著。根据研究结果,可知太行山东坡中段植物垂直分布格局,是群落垂直演替进程、环境因素和人为干扰协同作用所形成的结果。
综上,当前对太行山区植被垂直梯度分布和群落演替格局的研究,多数集中于较小的区域或流域尺度,由于太行山区纬度跨度较大,北段、中段和南段的环境因素、水土资源耦合程度、山区产水、耗水状况等均有较大差异,各区域的物种垂直分布特征差异也较大。因此,结合当前新的研究视角和数据分析方法,对整个太行山区尺度的植被时空分布格局,尤其对地理位置较为特殊的太行山区在气候变化背景下的植物群落演替规律的研究仍需进一步加强,以期为脆弱山地生态系统的保护、恢复和可持续利用提供理论支持。
