钦江河口潮滩红树林群落空间分布
2022-07-06王日明梁喜幸周晓妍黄鹄戴志军
王日明,梁喜幸,周晓妍,黄鹄,戴志军
1.北部湾大学钦州市环境生态修复重点实验室,钦州 535011;
2.北部湾大学广西北部湾海洋灾害研究重点实验室,钦州 535011;
3.华东师范大学河口海岸学国家重点实验室,上海 200062
1 引言
生长于热带亚热带河口潮滩的红树林在沿海城市抵御海洋灾害、维持河口生物多样性以及高存贮碳等方面具有不可或缺性。然而,因海平面上升、土地利用变化与海岸城市扩张等作用,世界较多河口红树林都出现大规模生境破坏及损失,这很可能对沿海生态安全带来风险(Lovelock 等,2015)。由于红树植被茂密且所处的狭窄潮间带出露时间短,到目前为止很难获得准确的红树林空间分布及生态学特性等资料。
基于此,较多成果从技术层面以定量潮间带红树林面积、生物量与植被结构(Rumondang等, 2022)。譬如,利用Landsat TM/ETM+/OLI 监测红树林的动态变化(Long 等,2021),或把激光雷达(Mazzini 等,2007)、多光谱(Hu 等,2020)与高光谱(Wulder 等,2012)用于红树林分布研究;将高空间分辨率及多光谱特性的多源数据融合,监测红树林类别(Simard 等,2002;李春干,2002)、估算红树林生物量(Satyanarayana 等,2011)、分层结构(Kovacs 等,2010)、群落特征参数(Shah 等,2005)、树高(Proisy 等,2007)及平均冠幅(Satyanarayana 等,2011)等生态学参数特征。同时,一些工作则基于遥感技术开展了红树林湿地动态演替及影响机制的研究等(Long 等,2021;Wang 等,2019)。也有工作将遥感影像反演以揭示红树林应对气候变化、海平面变化(Doyle 等,2009)。就北部湾沿岸红树林而言,贾明明等利用6 景Landsat 数据获取了较高精度的该区域近40年红树林分布地图(贾明明,2014;Jia等,2014)。Long 等(2022)基于长期Landsat 影像揭示了区域南流江河口红树林空间变化格局。李春干和代华兵(2015)基于航空图像、遥感影像(SPOT 1—5 图像、Landsat 5 TM图像、ALOS图像)和野外调查发现北部湾沿岸近50年红树林面积减少了22.16%,其中斑块消失、碎化和萎缩是红树林面积减少的主要途径。
毋庸置疑,遥感技术在红树林信息解译与提取方面提供实时、快捷和大范围的优势。但红树林生长于狭窄的潮间带,不同种群的红树林混杂而生,很难利用遥感技术甄别;加之当卫星过境如恰值高潮,则部分红树林被淹没亦将影响遥感影像反演红树林信息,由此导致对红树林面积及生物学习性的量化误差。显然,很有必要将遥感技术与野外调查等多技术结合以甄别红树林空间分布格局与表征红树林物种变化的各种参数特性。
与分布在一般为数百米宽的狭窄海岸潮间带红树林比较,河口生长的红树林自海向陆可从口外海滨延伸到感潮河道;同时河口水位、盐度与沉积物的变化梯度要远高于海岸潮间带。这意味着河口红树林远比海岸潮间带红树林空间演替及其变化要复杂,利用遥感技术来提取河口潮滩的红树林信息必然将出现较大误差,很难甄别河口区红树林种属。同时,当前国内外针对红树林的研究工作主要聚焦在热带与亚热带海岸,探讨河口自海向陆潮间带红树林种群空间演替过程的成果仍较缺乏(Woodroffe 等, 2016)。有必要指出的是,河口同样遭受高强度人类活动的影响,河口红树林的损失并不亚于海岸潮间带,对河口潮间带红树林群落甄别、红树林受损缘由及机理的分析应是理解河口红树林生长发育过程及恢复的关键所在。基于此,本文以北部湾钦江河口潮滩为研究区,试图结合历史遥感影像、无人机正射影像和实测水文及沉积观测资料,阐明钦江河口红树林植被的分布特性、空间变化格局及可能的影响机制,这能为热带河口潮滩红树林的研究、保护与红树林生态修复工程提供重要理论与技术借鉴。
2 研究区域与方法
2.1 研究区域概述
研究区位于茅尾海的钦江河口(图1)。钦江是广西独流入海的第二大河,全长179 km,于尖山镇横山头分为二支,分别于犁头咀、沙井注入茅尾海。该区域一般每日发生高、低潮各一次,半月周期新老潮期交替之日则发生高、低潮各两次,属不正规混合全日潮型(常涛等,2015)。涨潮潮差最大为2.21 m,平均0.97 m,落潮潮差最大为2.20 m,平均为1.01 m。钦江尖山镇河段为钦江感潮河段上限,涨潮历时最大为505 min,平均264 min;落潮历时最大为1424 min,平均1122 min。

图1 钦江河口监测站点布置Fig.1 Layout of monitoring stations at Qinjiang Estuary
受两入海河道径流与潮汐综合作用,钦江河口在大新围与坚心围沙岛外滩及河口感潮段形成红树林潮滩。该区域分布有红树植物11 科16 种,于2005年纳入茅尾海自治区级红树林自然保护区。该区域分布面积最大的红树植物为桐花树(Aegiceras corniculatum),其次是无瓣海桑(Sonneratia apetala)。同时还有部分海漆(Excoecaria agallocha)、木榄(Bruguiera gymnorhiza)、红海榄(Rhizophora stylosa)、秋茄(Kandelia candel/Kandelia obovata)、小花老鼠簕(Acanthus ebracteatus)等。感潮段河岸高潮位以上分布有半红树植物苦郎树(Clerodendrum inerme)、黄槿(Hibiscus tiliaceus)。此外,在该河段潮滩还零星分布有芦苇(Phragmites communis)、茳芏(Cyperus malaccensis)等草本植物。
2.2 实验设计与研究方法
2.2.1 无人机影像采集与影像数据处理
在2021年4月28日和2021年5月16日对钦江河口堤外潮滩与河口感潮段进行现场勘查,收集研究区资料,主要包括对各类红树植物分布极限点、典型植物群落记录与拍照,并用GPS 工具与奥维数字地图定位、标识。根据红树林空间分布信息进行航迹规划:考虑到航摄需要在低潮期间进行,但研究区范围较大且需要无人飞机多次航摄,把钦州河口分为堤外潮滩(2 个区域)与感潮河段共3 个区域,对堤外潮滩采用区域规划方式,对感潮河段采用航带规划方式(图2)。

图2 无人机航拍航线规划Fig.2 The UAV aerial photography route planning
2021年5月,通过卫星影像和实地调研的方式对研究区域进行综合评估,结合飞行安全、飞行效率等因素确定无人机起降点备降点、盘旋点等信息。根据研究目标和无人机载荷的相关参数确定无人机飞行高度、航飞路线。结合飞行窗口期的风力、能见度、太阳光线照射角度等信息对无人机进行详细的地面调试、检测准备,利用地面站控制系统控制无人机垂直起飞,到达预定高度后切换成为固定翼巡航模式,本次航摄采用复合翼垂直起降无人机,搭载sony5100 可见光摄像头对研究区进行航摄,获取研究区的正射影像数据,影像数据包含R、G、B 这3 个波段。飞行高度250 m,航向重叠率80%,旁向重叠率70%,地面分辨率0.05 m。共飞行7 个架次,每个架次飞行时间45 min 左右,全部航拍任务历时325 min,共获取5692张有效像片,覆盖面积11.52 km2。
对采集的无人机影像使用Pix4D 软件进行几何纠正与拼接,利用“Grographic Ortho-Metric”功能生成正射影像图;随后利用“htitchMe”功能镶嵌正射影像;再利用行正射“station operator”功能评定精度以及纠正、修改在镶嵌阶段产生的误差;最终得到研究区拼接后的影像,生成点云数据和数字表面模型DSM(Digital Surface Model)影像数据。
2.2.2 无人机影像与卫星遥感影像红树林数据提取
本研究结合红树林生长特点和所获取的无人机影像与2016年卫星遥感影像(数据来自图新地球4.0 版2016年12月28日分辨率为0.27 m 的谷歌历史影像)的参数信息,根据不同物种的光谱特征、纹理特征、几何形状和拓扑特征等(冯家莉等,2015),采用目视判读法,结合现场勘查校核,确定桐花树、无瓣海桑解译标志(图3)并识别,提取钦江河口桐花树、无瓣海桑等红树林空间分布信息。因其他红树植物个体体量小、分布范围窄且大多分布于桐花树与无瓣海桑林下,或与这两种红树植物混生在一起,所提取数据难以在图上清晰表达,故本研究仅提取2016年与2021年桐花树、无瓣海桑等红树植物在钦江河口堤外潮滩的空间分布。结合现场校核,在图上仅表达各红树植物沿钦江河口的分布极限位置。

图3 无瓣海桑与桐花树解译识别Fig.3 Interpretation and identification of S.apetala and A.corniculatum
采用解译结果到现场校核与现场植株定位到解译数据确认的双向验证法对无人机图像目视解译精度进行验证。第1 步,从解译结果到现场验证,主要验证解译结果的正确性:根据无瓣海桑解译标志,对无人机影像目视解译结果中,在研究区内按分布均匀原则随机选取识别的无瓣海桑40 棵,记录其分布位置后再去现场勘查确认,确认结果所选40棵中有39为无瓣海桑,1棵为海漆。第2步,从现场植株定位到解译数据校核,主要校核是否漏译:在钦江河口堤外红树林潮滩,现场选取树龄5年生左右的中小型无瓣海桑植株40 棵,用RTK 标定其位置,再对根据无人机影像目视解译数据进行校核,查证所有无瓣海桑植株是否被解译。验证结果为所选取的40 棵无瓣海桑中有39 棵被正确识别并在解译结果中标识,有一棵漏译(图4)。综上,对无人机图像目视解译所抽取总样本为80 棵,其中解译正确的为78 棵,则目视解译的精度为97.5%。

图4 无人机影像无瓣海桑目视解译精度验证Fig.4 Validation of visual interpretation accuracy of Sonneratia apetala images from UAV
2.2.3 实验站点布置
结合无人飞机航摄进行沿程校核的同时,专门布设定点观测红树林植物群落结构,并进行相应水体盐度测定与潮滩沉积物粒度分析。考虑到犁头咀入海河段采用人工硬质驳岸,近岸几乎没有潮滩,河段内大规模生蚝养殖、渔民赶海等人工干扰导致该河段到外滩极少有红树林生长,故在该河段没有布置监测和校核站点。而在沙井入海河段,部分岸堤采用人工硬质驳岸,但全线较为平缓,岸外有宽窄不一的潮滩。同时,该河段弯道较多,沿线内湾分布有大小不一潮滩。沿该河段上溯,感潮段潮滩有较多红树植物分布。故本研究沿沙井入海河段,以HS00(21°51′21.54″N,108°35′54.06″E)为河口基点,上溯到感潮段最上限尖山镇HE00 点(21°55′42.19″N,108°37′05.02″E)进行现场布点(图1)。沿感潮段在河道转点及河口红树林潮滩边缘依次布置10 个监测点(QJ01—QJ10),在HS00 向海段布置3 个监测点(HO01—HO03),同时在大新围外滩离堤约20 m 的红树林滩地设置5个监测点(DW01—DW05)(图1)。
2.2.4 沉积物及水样样品采集测定
(1)2021年3月28日在外滩与河道现场校核监测点(图1)的红树林片区采集2 cm 厚表层土,带回实验室干燥后,用筛分法分析各沉积物粒径组成。
(2)堤外潮滩水样采集与盐度测定:于2020年11月8日区域涨潮接近高程位开始,分别在外滩站点(图1)每隔30 min 取表水采样,每个点共采10 个水样带回实验室测定盐度,取其平均值为该站点的表水盐度值。
(3)感潮河段水样采集与盐度测定:在涨潮接近高潮时,从口外站点HO03 出发,驾船按5 km/h 时速逆流上溯至HE00 站点,随后从HE00站点驾船按5 km/h 时速顺流至口外站点HO03(图1)。途中每隔5 min 采一次表层水样,带回实验室测定水体盐度。根据各取样点与布设站点的距离,采用内插值法,计算出各站点的盐度。以顺、逆流各站点两次盐度的平均值为该站点的盐度值。
2.2.5 钦江河口潮滩红树植物分布现场勘查
(1)堤外潮滩红树植物分布现场勘查。沿大新围与坚心围向海岸线,勘查红树植物空间分布,并分析记录群落结构。对有代表性的红树植物种类与典型群落结构拍照,并用GPS 工具与奥维数字地图定位、标识,以便后期影像数据处理时核验。沿堤外潮滩红树林向海林缘线,探查向海分布的红树植物种类,观察记录不同红树植物二年生与当年生幼苗生长极限位置。当年生红树植物为红树向海生长的极限位置(王日明等,2020)。随后,获取当地海平面所在位置高程点,利用中海达V90 GNSS RTK 进行区域横向断面的测量,作业过程中,测定点在固定解状态下,有效卫星个数应多于5 个,PDOP 值小于6,高程测量精度约为0.02 m。根据海事资料,钦州湾平均海平面比1985 黄海基面高0.36—0.38 m,此处取0.37 m。则所测定的高程数据减去0.37 m,即为该点以当地海平面为基准的高程值,其中一年生红树林所在位置的高程即为极限位置。
(2)感潮河段红树植物分布现场勘查。沿钦江河堤上溯,以及驾船沿河段内林缘线上溯,勘查红树植物空间分布。随后记录各红树植物沿江分布状况与典型群落结构、拍照定位。各红树植物沿江分布距HE00 站点最远点即为该类植物向陆分布上限。半红树植物与非红树植物沿江分布距HE00 站点最近点即为该类植物分布下限,最远点为分布上限。
3 结果分析
3.1 钦江河口红树林群落分布特征
钦江河口潮滩红树林群落分布表现为红树植物种类与数量自海向陆逐减的特点。红树植物向陆分布最远为距离HE00站点11.00 km处的QJ09河段,植物种类为桐花树;红树植物向陆成林分布最远为距离HE00 站点9.55 km 处的QJ07 河段,植物种类为无瓣海桑(图5)。
3.1.1 钦江河口潮滩植物分布特征
钦江河口潮滩分布的红树植物种类主要有桐花树、无瓣海桑、海漆、木榄、红海榄、秋茄、小花老鼠簕及部分半红树植物黄槿、苦郎,非红树草本植物芦苇、茳芏。按其数量与分布范围,以桐花树为主,数量多,分布范围广。钦江河口潮滩桐花树面积约400.00 hm2,其向海分布自堤外潮滩红树林林缘线外0.67 km 处(108°34′28.31″E,21°50′35.96″N),所处位置高程(以当地平均海平面为准,下同)为-0.37 m 有当年生桐花树小苗生长(图5)。其向陆分布自HS00 站点沿钦江上溯8.91 km 处潮滩有约20 m2小片林,11.00 km 处有当年生桐花树小苗生长(图5、图6)。

图5 钦江河口潮滩植物分布特征Fig.5 Distribution characteristics of plant species in tidal flat of Qinjiang Estuary

图6 钦江河口感潮段植物分布极限Fig.6 Distribution limit of plants in tidal section of the Qinjiang Estuary
2002年钦州市引种无瓣海桑20000 株种植于康熙岭镇标准海堤外滩,面积约15 hm2。2011年入侵钦江河口潮滩后,得到快速发展。2016年钦江河口潮滩无瓣海桑面积发展到0.30 hm2,至2021年已达5.62 hm2,平均年增长79.6%,并呈现继续扩张趋势。同期,钦江河口红树林总面积也在增长,但平均年增长仅46.0%。可见,外来物种无瓣海桑在钦江河口适应能力强,发展速度快。无瓣海桑主要集中分布于堤外潮滩,首先以单株分散入侵桐花树林后,再逐渐发育成片林。无瓣海桑片林下的桐花树等灌木逐渐衰老死亡。无瓣海桑向陆分布到HS00 站点沿钦江上溯9.55 km处(图6)。
小花老鼠簕、秋茄、海漆、木榄、红海榄等主要散布于堤外潮滩桐花树片林中,数量少,分布面积小。小花老鼠簕沿江分布上限为距HS00 站点5.90 km的潮滩(图6)。
3.1.2 钦江河口堤外潮滩植物群落格局
钦江河口大新围与坚心围堤外海滩形成以桐花树为主,少量无瓣海桑、海漆、木榄与红海榄,极少数小花老鼠簕与秋茄组成的红树林群落复合体(图7)。在堤外潮滩,向海滩地局部形成以桐花树为主的“无瓣海桑—桐花树”群落,部分无瓣海桑密集区域形成“无瓣海桑纯林”。近堤岸区域,局部形成“无瓣海桑+海漆—桐花树+秋茄+卤蕨+小花老鼠簕—苦郎”、“黄槿+海漆—桐花树+小花老鼠簕”、“海漆—桐花树”群落(图7)。其中,黄槿、苦郎等半红树植物仅分布于堤岸高高潮位以上的岸堤,小花老鼠簕主要分布于近岸高潮位潮滩。


图7 钦江河口潮滩植物群落格局Fig.7 Plant community patterns in tidal flats of the Qinjiang Estuary
3.1.3 钦江河口感潮段潮滩植物群落格局
钦江河口感潮段潮滩红树林分布表现为向海河口段红树林植物种类较丰富、分布面积较广,以“无瓣海桑—桐花树”纯红树植物群落为主。沿河上溯种类与数量逐渐减少(图5、图7),表现出红树植物与半红树植物、非红树植物混生的特点。HS00 至QJ04河段是海陆过渡带,河段两侧有较宽潮滩,河中间有江心洲潮滩,红树林面积53.60 hm2。该区域形成以桐花树为主的“无瓣海桑—桐花树”群落。
QJ04 至QJ07 河段受潮汐作用逐渐减弱,河段沿线潮滩分布的植物群落结构表现为:“无瓣海桑—桐花树”群落、“芦苇+桐花树”或“桐花树+茳芏”混生群落;中高潮滩林丛边缘有小量小花老鼠簕分布,近高潮带河堤中上部有部分苦郎树分布。QJ05 至QJ07 河段潮滩遍布芦苇,在芦苇丛中杂生有部分桐花树,有少量3—5年生的无瓣海桑分布,形成以芦苇为主的“无瓣海桑—芦苇+桐花树”混生群落结构。
QJ07 至QJ10 河段受潮汐作用进一步减弱,沿线潮滩少,是红树植物与非红树植物混生区。该区域群落结构表现为“芦苇+苦郎树”或“芦苇+茳芏+苦郎树”的混合植物群落。
3.2 钦江河口盐度梯度变化特征
在大潮高潮涨平期间测得的钦江河口盐度表明,自口外站点沿河道向陆逐渐降低。大潮高潮时感潮河段自HO03站点平均盐度14.2‰,至HS00站点平均盐度13.8‰,盐度变化较平缓。自HS00站点沿江上溯,至距HS00站点5.90 km处的QJ05—QJ06 河段,盐度从11.3‰快速降低至3.7‰。随后QJ07 站点盐度小幅度上升后又缓慢降低,至距HS00 站点13.20 km 的HE00 处降到0.0‰(10 次测量结果有两次盐度分别为0.02‰与0.01‰,其他显示为0.00‰)(图8)。
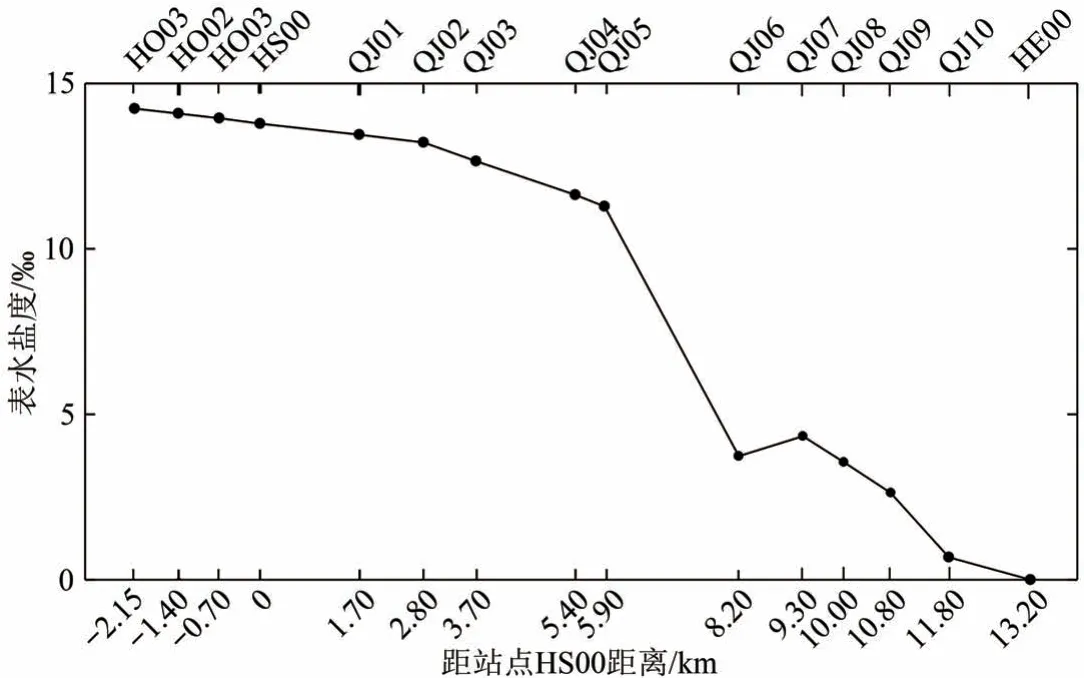
图8 钦江口感潮段河道表层水体盐度特征Fig.8 Characteristics of surface water salinity along the tidal reach of Qinjiang River
自HS00 沿河道上溯,距离越远,水体盐度越低。感潮段最远距离为距河口13.20 km 的HE00 站点,该点是钦江河口感潮段最远极限点。
3.3 钦江河口红树林潮滩沉积物粒度特征
钦江河口感潮段沉积物属沙质土,土壤颗粒较粗。其中以距HS00 站点4.00—8.00 km 河段的沉积物粒径最粗,粒径大于0.125 mm 的含量达67%—79%,粒径在0.063—0.125 mm 的含量达20%—30%(图9(a))。堤外潮滩近岸红树林丛内土壤颗粒相对较细,粒径大于0.125 mm 的含量为59%—71%,粒径在0.063—0.125 mm 的含量为27%—33%(图9(b))。
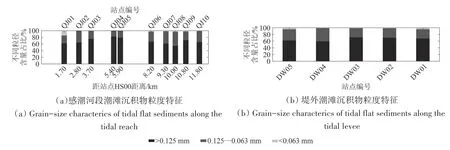
图9 钦江河口潮滩沉积物粒度特征Fig.9 Grain-size characteristics in tidal flat sediments of the Qinjiang Estuary
4 讨论
4.1 钦江口红树林分布格局
影响红树林在河口分布格局的主要因素有水体盐度、潮位高低、土壤粒径等,受控于河口复杂的水文与地貌条件,河口红树林自海向陆都展现出距离河口越远,红树群落趋于简单和长势较差的格局(王日明等,2020)。通过对钦江河口红树林分布格局研究,其结论与上述基本吻合。钦江河口潮滩红树林群落可以归纳为4种类型。
第1种为“纯红树植物”群落。大新围与坚心围外滩遍布桐花树,无瓣海桑分散于桐花树林丛中,形成以桐花树为绝对优势种的“无瓣海桑—桐花树”群落。外滩近岸区域,还有部分海漆、红海榄、小花老鼠簕、卤蕨等的分布(图7)。于HS00—QJ05 河段,仍以桐花树为优势种,无瓣海桑散布于桐花树林丛中,形成以“无瓣海桑—桐花树”群落为主的群落结构。部分中高潮滩桐花树林缘有小花老鼠簕分布,局部形成“无瓣海桑—桐花树+小花老鼠簕”群落(图7)。
第2种类型为“红树植物—半红树植物”混交群落。该类型主要位于河口堤外潮滩高高潮位与QJ01—QJ03 河段河堤高高潮位。分布于高高潮位及以上的苦郎与黄槿,与分布于高高潮位的海漆、桐花树、小花老鼠簕、卤蕨等构成“黄槿+海漆—桐花树+小花老鼠簕+卤蕨”混交群落。
第3种类型是“红树植物—半红树植物—非红树植物”混交群落,主要位于QJ05—QJ08 河段的中高潮滩。在该区域开始出现大面积的芦苇分布,桐花树苗混生于芦苇丛中,少量小花老鼠簕杂生于芦苇丛或林缘,在高潮位以上的堤岸有部分苦郎分布,该河段潮滩形成了“无瓣海桑—桐花树+芦苇+小花老鼠簕—苦郎”或“无瓣海桑—芦苇+桐花树+小花老鼠簕—苦郎”混交群落(图7)。
第4 种类型为QJ08-HE00 河段的“半红树植物—非红树植物”群落。自QJ08 往上河段,红树植物分布极少,在高潮位以上的堤岸分布有部分苦郎与黄槿等半红树植物,潮滩以芦苇为主,有少量茳芏分布,形成“非红树植物+苦郎—芦苇/茳芏”混交林(图7)。
4.2 盐度对植物群落的影响
盐度是制约红树林分布的关键因子之一,不同红树植物种类所能适应的盐度范围不同(王日明,2020)。钦江河口几种主要的红树植物耐盐性如表1所示。

表1 钦江河口红树植物耐盐特性Table 1 Salt tolerance characteristics of mangrove plants in Qinjiang Estuary
钦江河口大新围外滩盐度高,感潮河段离海距离越远,盐度越低。相应地,盐度较高的外滩向海区域群落结构表现为无瓣海桑散布于大面积桐花树林丛(图5),且伴生少量秋茄;地势较高的滩地在桐花树林丛偶分布红海榄与木榄,近岸桐花树林缘分布有部分小花老鼠簕。从HS00 至HE00 河段,平均盐度从11.2‰逐渐下降至0.0‰,潮滩红树林数量与种类也逐渐减少,由“纯红树植物”群落向“红树植物—半红树植物”群落、“红树植物—半红树植物—非红树植物”混交群落、“半红树植物—非红树植物”群落转变,群落中红树植物所占比例逐渐降低。
4.3 潮滩沉积物粒度变化对河口红树林群落发育的影响
土壤为植物提供了耐以生存的立地条件,土壤颗粒组成决定土壤的通气透水性与保水保肥性(姜芃,2011)。粒径小的粘质土壤保水保肥性好,有利于植物地上部分生长;而粒径粗的砂质土壤透水透气性强(李百凤等,2007),有利于植物根系发育。生长于潮间带的红树植物由于长期交替浸淹水,对立地的通气透水性要求更高。尤其是根系尚不发达的一二年生幼苗更是要求其立地土壤排水良好。
钦江河口堤外近岸潮滩沉积物粒径小,距HS00 站点4.00—8.00 km 的QJ03—QJ07 段潮滩粒径较大(图9(a))。相对应地,堤外潮滩红树林成年树片林较集中分布于近岸区域,生长发育良好,种类品种多。受红树林片林影响,涨潮时自林外缘向岸水动力逐渐减弱,落潮水流缓慢。粗颗粒自外向岸逐渐下沉,至近岸滩地水动力达到最弱,沉积物粒径最小。因此,堤外潮滩由岸向海沉积物粒径由细变粗,红树林分布也由岸向海扩张,由分布密集变为稀疏。桐花树二年生苗向海分布极限距林缘线0.43 km 与当年生苗向海分布极限距林缘线0.67 km,也正从另一侧面表明了植物分布与土壤粒径的相关性。
钦江河口感潮河段,自HS00向QJ03河段受潮汐与径流水动力综合影响,在该区域易形成紊流区,更有利于来自海水的较细颗粒的下沉。而在QJ03—QJ07 河段,是潮水与径流的交锋区,尤其是QJ05—QJ06 河段,受另一入海河道涨落潮水流的作用,该区域潮汐与钦江径流作用频繁,水流紊乱。因此,水体持沙力较强,沉积物粒径相对较粗。相应地,HS00—QJ03 河段潮滩面积大,分布的红树植物较多且生长良好;QJ05—QJ06 河段横向剖面较陡,潮滩窄,红树植物少,仅沿河岸呈窄长带状分布(图5),林缘线一二年生幼苗较多。QJ06 上溯河段受涨落潮作用越来越弱,潮滩面积逐渐减少,红树植物种类与数量也相应减少,至距HE00 站点11.00 km 处当年生桐花树幼苗,是钦江沿江上溯红树林分布极限(图6、图7)。
5 结论
受盐度、潮汐与土壤结构等因子作用,北部湾北部钦江河口潮滩的红树植物群落自海向陆呈梯度变化,主要结论包括:
(1)自海向陆红树林种类与数量逐渐减少,群落结构由“纯红树植物”、“红树植物—半红树植物”、“红树植物—半红树植物—非红树植物”、“半红树植物—非红树植物”群落变化。
(2)潮位与盐度是影响红树植物在钦江河口潮滩分布范围的关键要素。桐花树是钦江河口潮滩红树林群落的绝对优势种,其向海分布极限为距林缘线0.67 km 的当地平均海平面以下0.37 m 潮滩。向陆分布极限为距河口HS00 站点10.18 km 感潮段潮滩。红树林片林向陆分布极限为距HS00 站点9.55 km 处潮滩,该位点所测盐度值为4.30‰,主要植物种类为无瓣海桑。
(3)沉积物粒径影响到红树林胚胎发育与成年树的生长。堤外潮滩向海潮滩沉积物粒径较大,距HS00 站点4.00—8.00 km 河段潮滩沉积物粒径粗,落潮后有利于胚胎的着陆与根系发育。河口堤外潮滩近岸较细粒径的土壤结构具有较好的持有养分能力,能促进红树林成年树生长。
