苦荞漆酶基因的克隆与生物信息学分析
2022-07-06杨晓琳蔡苏云贺润丽尹桂芳王艳青卢文洁孙道旺王莉花
杨晓琳 段 迎 蔡苏云 贺润丽 尹桂芳王艳青 卢文洁 孙道旺 王莉花
(1山西中医药大学中药与食品工程学院,030619,山西太原;2云南省农业科学院生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业农村部西南作物基因资源与种质创制重点实验室,650221,云南昆明)
苦荞[Fagopyrum tataricum(L.)Gaertn]为双子叶蓼科一年生草本植物,作为药食兼用作物,富含抗性淀粉、高活性蛋白和膳食纤维等营养成分,以及芦丁和槲皮素等黄酮类生物活性成分,具有降压、降脂和降胆固醇等功效[1-4]。由于苦荞米具有极高的营养药用价值,其产品也在不断被研发,开发方向日益广泛,使得人们对苦荞米的需求量不断提高。根据果壳厚度,苦荞可分为厚壳型和薄壳型2类[5]。普通栽培苦荞由于种皮厚、难脱壳形成整粒生荞米,降低了苦荞的营养活性成分和加工利用价值;薄果壳突变体由于易脱壳成整粒生荞米,但适应性差和产量低等导致其难以推广种植,所以薄果壳苦荞成为研究和育种的热点。
漆酶(laccase)是一种含铜的多酚氧化酶,广泛存在于真菌、细菌和植物中,在高等植物中通过氧化还原反应催化各种底物,参与到植物的各种生理反应中。研究[6-8]发现拟南芥共有17条漆酶基因(AtLAC1~AtLAC17),根据功能分为6类,其中AtLAC4和AtLAC17参与木质素合成,既引导又指导细胞壁的木质化。已有证据[9-10]表明漆酶能够参与细胞壁形成,参与调控木质素单体的聚合过程,在植物木质素合成中起着重要的作用。在杨树中抑制漆酶基因的表达,木质素总含量组成都没有发生变化,但导致杨树木质部纤维细胞壁的变形脱落,表明漆酶对木质部纤维及细胞壁结构完整性有重要影响[11]。
吴朝昕[12]对苦荞果壳成分进行研究,发现厚果壳苦荞果壳木质素含量高于薄果壳苦荞。本课题组前期研究以云荞1号与小米荞杂交F2为遗传材料,利用转录组测序技术,分析比较苦荞薄果壳和厚果壳中差异表达基因,发现木质素生物合成途径大部分基因在苦荞厚果壳的表达量高于薄果壳,推测苦荞薄果壳性状与木质素成分合成和积累量低有关。对转录组数据中木质素合成途径差异基因分析结果表明,漆酶基因在薄果壳苦荞中的表达量显著低于厚果壳苦荞。基于转录组测序结果,本试验以苦荞为材料,运用RT-PCR技术对木质素生物合成途径的关键基因漆酶基因进行克隆,并对其进行序列、蛋白理化性质、系统进化树和实时荧光定量PCR表达分析,为阐明苦荞薄果壳形成的分子机制提供理论依据。
1 材料与方法
1.1 试验材料
分别采用云荞1号(云南省农业科学院生物技术与种质资源研究所选育的厚果壳品种)和小米荞(云南地方薄果壳品种)不同发育期果实的混合材料进行基因克隆。选用云荞1号和小米荞结实期不同部位组织材料(即结实期的叶、花、茎、果壳和种子)进行荧光定量PCR分析。
1.2 试验方法
1.2.1 引物设计与合成 根据转录组测序序列及漆酶中4个铜原子附近的氨基酸保守序列,分别设计了8条引物用于RT-PCR基因克隆(表1),由生工生物工程(上海)股份有限公司合成。
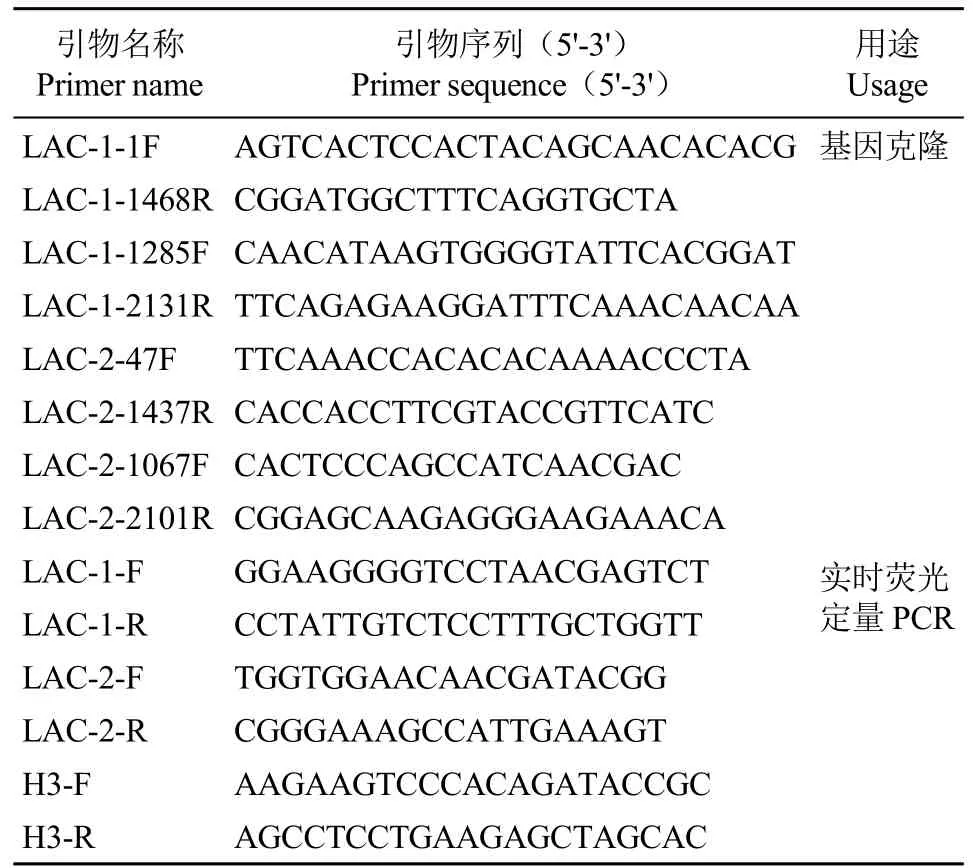
表1 引物序列及用途Table 1 Sequences of primers and their usage
1.2.2 RNA提取与cDNA第1链合成 根据Trizol提取试剂盒,由生工生物工程(上海)股份有限公司提供,操作说明分别对苦荞厚果壳和薄果壳混合材料进行总RNA提取。使用1.5%琼脂糖凝胶电泳检验提取结果,选择完整性较好、条带清晰且无降解的总RNA进行反转录。
在0.2mL的PCR管中加入总RNA 5μL、随机引物 1μL和 ddH2O 1μL,70℃温浴 5min,冰浴2min;离心加入试剂 5× First-Strand Buffer 2.0μL、10mmol/L dNTP 0.5μL、Rnase inhibitor 0.25μL 和Reverse Transcriptase 0.25μL,总体积 10.0μL,42℃温浴60min,72℃温浴10min。
1.2.3 RT-PCR基因克隆 以苦荞总RNA反转录合成的cDNA第1链为模板,采用25μL的PCR反应体系进行PCR扩增,包括2×GC Buffer I 12.5µL、上游引物(10µmol/L)0.5µL、下游引物(10µmol/L)0.5µL、dNTP(10mmol/L)0.2µL、ddH2O 10.1µL、cDNA模板1µL和Taq酶(5U/μL)0.2µL。PCR反应扩增程序为95°C预变性3min;94°C变性30s,58°C 退火 30s,72°C 延伸 90s,循环 33 次;72°C修复延伸7min。PCR扩增产物经1%琼脂糖凝胶电泳检测,用试剂盒回收纯化目的条带,由生工生物工程(上海)股份有限公司进行测序。
1.2.4 漆酶基因及其编码蛋白序列分析 将克隆所得基因序列在NCBI(http://www.ncbi.nim.nih.gov/blast/Blast.cgi)进行 Blast比对,在 ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)中查找基因的ORF,翻译获取氨基酸序列。使用SMART(http://smart.embl-heidelberg.de/)工具在线对苦荞漆酶蛋白结构域进行预测分析;用在线软件EX Pasy(https://web.expasy.org/protparam/)分析编码蛋白基本理化特性,用protscale(https:/web.expasy.org/protscale/)工具对其亲、疏水性进行预测;利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)在线分析苦荞漆酶蛋白二级结构;应用SWISS-MODEL(http://swiss model.expasy.org/)模拟构建FtLAC-1和FtLAC-2蛋白的三级结构模型。在NCBI上的Blastp同源比对,将高分值序列在DNAMAN进行序列比对,并用MEGA 6.0软件构建苦荞漆酶与其他植物漆酶NJ系统进化树[13-15]。
1.2.5 漆酶基因荧光定量PCR(qRT-PCR)分析用Primer Premier 5.0软件设计目的基因及内参基因引物序列(表1)。qRT-PCR扩增程序:95℃预变性30s,随后进行45个循环:95℃变性5s,60℃退火30s。以苦荞FtH3(JF769134.1)为内参基因,利用2-ΔΔCt法计算出基因的相对表达量。
2 结果与分析
2.1 苦荞RNA提取及漆酶基因的克隆结果
RT-PCR扩增产物电泳结果如图1所示,在800和2500bp左右有明显的扩增条带。PCR产物回收测序,将测序序列进行拼接组装,得到最终序列。经过序列比对发现克隆到的2条laccase基因在云荞1号(厚果壳)和小米荞(薄果壳)中的序列一致性为100%,且与转录组测序序列高度一致,可见LAC-1和LAC-2基因在薄壳苦荞和厚壳苦荞中没有突变和分化,所以命名为FtLAC-1和FtLAC-2。FtLAC-1和FtLAC-2基因开放阅读框长度分别为1695和1707bp,各编码564和568个氨基酸。
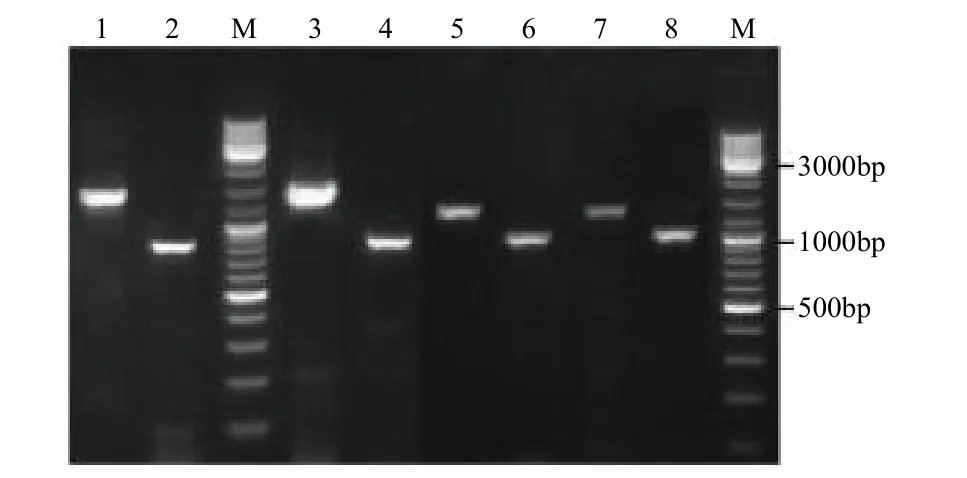
图1 苦荞LAC-1和LAC-2克隆基因电泳图Fig.1 Amplification of the LAC-1 and LAC-2 genes from tartary buckwheat
2.2 苦荞漆酶基因生物信息学分析
2.2.1 漆酶蛋白理化性质分析 使用Pfam对漆酶蛋白的结构域进行预测,结果(图2)显示,2个基因编码蛋白均含有3个经典铜离子结构域,即Cu-oxidase-3、Cu-oxidase和Cuoxidase-2结构域,说明FtLAC-1和FtLAC-2均可能具有漆酶蛋白的基本功能。利用EX PASy工具进行预测,进一步了解苦荞漆酶蛋白的理化特性以及潜在功能,结果显示,FtLAC-1和FtLAC-2蛋白分子量分别为61.41和62.47kD,理论等电点为9.45和9.41,FtLAC-1蛋白分子式为C2796H4336N760O771S15,带负电氨基酸35个,带正电氨基酸53个,不稳定系数为35.29;FtLAC-2蛋白分子式为C2837H4329N765O804S14,带负电氨基酸27个,带正电氨基酸41个,不稳定系数为28.46。
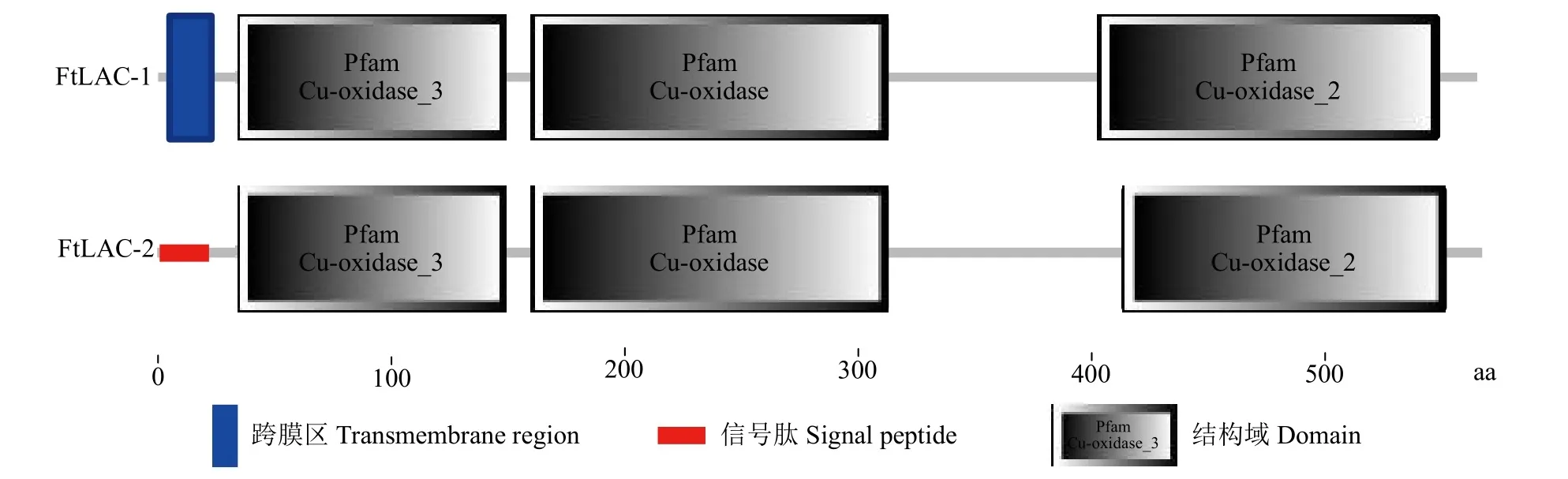
图2 FtLAC-1和FtLAC-2蛋白结构域Fig.2 Conserved domains of gene-encoded proteins of FtLAC-1 and FtLAC-2
对疏水/亲水性进行预测,结果见图 3,FtLAC-1和FtLAC-2平均亲水性分别为-0.030和-0.121,可知这2个氨基酸序列编码蛋白均为亲水性蛋白。
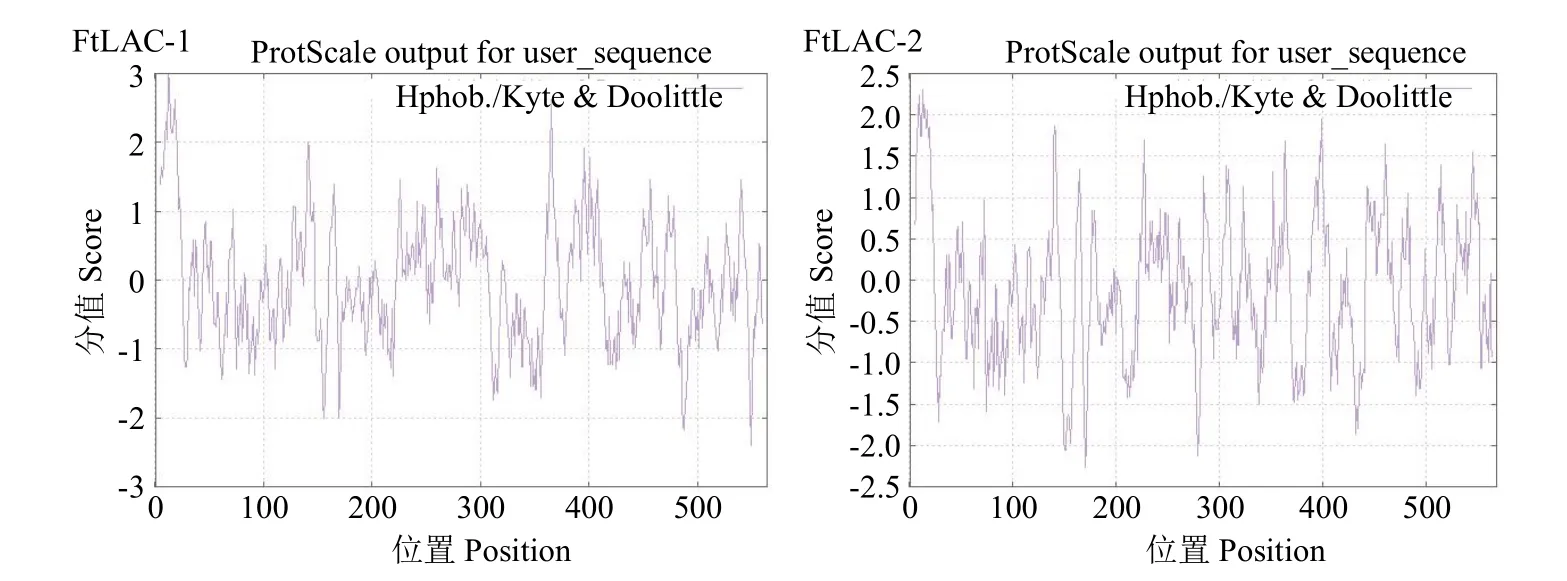
图3 苦荞漆酶亲疏水性分析Fig.3 Analysis of hydrophobicity of laccase proteins
2.2.2 漆酶蛋白结构分析 利用SOPMA在线对苦荞漆酶蛋白进行二级结构结构预测,表明FtLAC-1蛋白二级结构中α-螺旋占15.07%、无规则卷曲占50.89%、延伸链占27.48%和β-转角占6.56%;FtLAC-2蛋白的二级结构由12.68%的α-螺旋、6.34%的β-转角、52.29%的无规则卷曲和28.70%的延伸链构成。利用SWISS-MODEL对蛋白质的三级结构进行预测,可能的构型如图4所示。
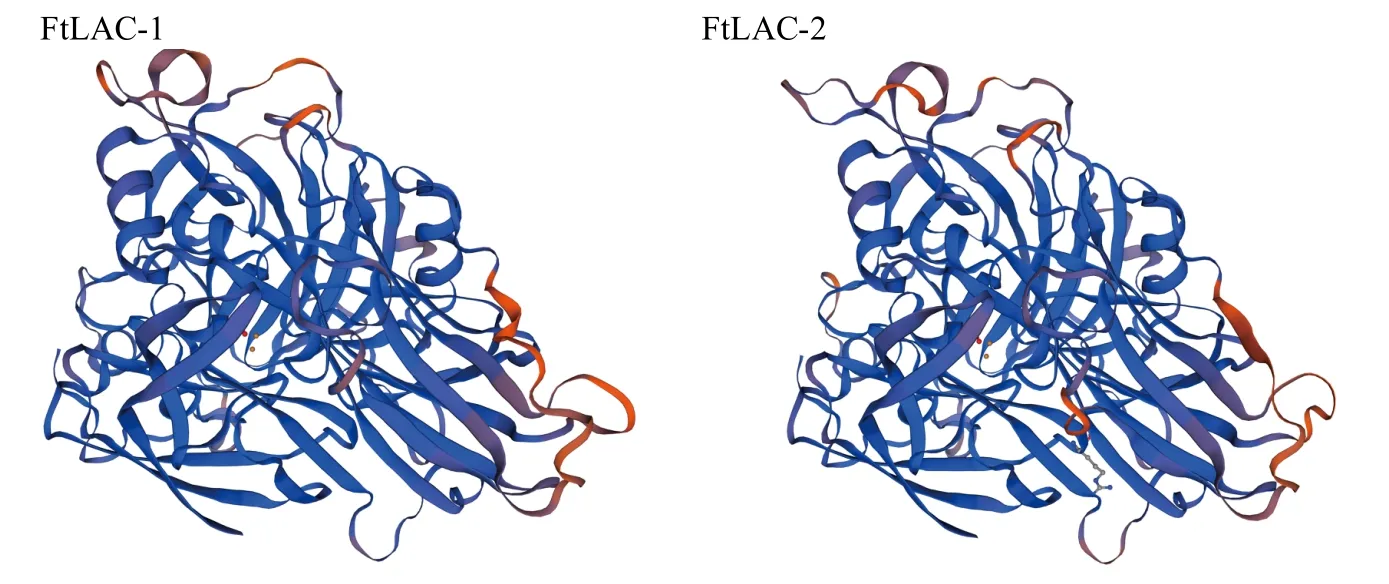
图4 苦荞漆酶蛋白三级结构预测Fig.4 The prediction of tertiary structural model of laccase proteins
2.2.3 漆酶蛋白同源比对及进化树分析 在NCBI中对FtLAC-1和FtLAC-2进行比对,结果显示2个氨基酸序列相似性并不高。经BLAST序列分析得到FtLAC-1与橡胶树Hevea brasiliensis(Willd.ex A.Juss.)Muell.Arg.(XP 021662642.1)和阿月浑子Pistacia vera L.(XP 031276156.1)的序列一致性分别高达82.62%和80.72%,FtLAC-2与甜菜亚种Beta vulgaris subsp.vulgaris(XP 010685719.1)和林烟草 Nicotiana sylvestris Speg.(XP 009796024.1)序列一致性分别为72.17%和72.06%。选取BLAST比对一致性较高的序列与之进行多序列比对,结果见图5。

图5 漆酶氨基酸序列比对Fig.5 Amino acid sequence alignment of different plant laccase proteins
为了分析苦荞漆酶蛋白基因FtLAC-1和FtLAC-2的亲缘进化关系,采用MEGA 6.0对苦荞漆酶基因编码的氨基酸序列同来自NCBI数据库中其他植物漆酶氨基酸序列进行系统进化树的构建,结果如图6所示,从进化树中可以看出FtLAC-1和FtLAC-2分别与不同植物分为两大类,FtLAC-2与甜菜单独聚在一个小分支,说明二者具有较近的亲缘关系;FtLAC-1与拟南芥AtLAC4聚在一大簇,FtLAC-2与拟南芥AtLAC17聚在一大簇,已有研究[6-7]证实拟南芥AtLAC4和AtLAC17参与木质素合成,推测FtLAC-1和FtLAC-2基因也可能参与木质素的合成。

图6 不同植物漆酶蛋白序列系统进化树Fig.6 Phylogenetic tree analysis of laccase proteins from different species
2.3 实时荧光定量PCR(qRT-PCR)表达分析
应用qRT-PCR技术分析FtLAC-1和FtLAC-2基因在厚果壳和薄果壳苦荞不同组织器官的表达量,结果见图7。从不同组织器官来看,漆酶基因FtLAC-1和FtLAC-2在苦荞不同组织的表达量不同,在厚果壳与薄果壳苦荞中均表现为在种子中的表达量高于其他组织器官,推测材料选取时期正处于苦荞果实生长膨大期,相关基因活跃,表达量较其他部位高。FtLAC-1在厚果壳苦荞果壳中的表达量约是薄果壳的9倍,在厚果壳苦荞花的表达量远远高于薄果壳;FtLAC-2在苦荞厚、薄果壳中表达量基本一样,但在厚果壳苦荞茎、花和叶中的表达量高于薄果壳苦荞。总体来看,FtLAC-1和FtLAC-2基因在厚果壳苦荞不同组织器官的表达量均高于薄果壳苦荞。

图7 FtLAC-1和FtLAC-2基因在各器官的相对表达量Fig.7 Relative expression of FtLAC-1 and FtLAC-2 genes in various organs
3 讨论
随着我国“三高”人数越来越多,苦荞显著的药理作用及营养价值引来食品和药品等行业越来越多的关注,所以开展易于加工且高产质优的薄果壳苦荞品种选育研究是医疗保健行业、农业生产和加工企业的共同需求[16]。
吴朝昕[12]研究发现薄壳苦荞果壳纤维素和木质素含量显著低于厚壳苦荞。本课题组前期研究转录组测序,分析发现木质素合成途径基因大多在薄果壳苦荞的表达量低于厚果壳,为了深入验证苦荞薄果壳形成的分子机制,本试验运用RT-PCR技术从云荞1号和小米荞的不同发育时期混合材料中成功克隆获得2条苦荞漆酶基因FtLAC-1和FtLAC-2,开放阅读框长度分别为1695和1800bp,编码564和568个氨基酸,经预测2个蛋白均为亲水性蛋白,此外,苦荞漆酶蛋白FtLAC-1和FtLAC-2都具有3个典型的铜区保守域Cu-oxidase-3、Cu-oxidase和Cu-oxidase-2,表明其与其他植物漆酶蛋白具有相似的功能。但2个蛋白的二级和三级结构特征存在差异,2个基因序列不具有较高相似性,其氨基酸序列同源性低。
研究[17]表明,植物漆酶参与调控单体聚合,在木质部或正在木质化的组织中表达量高,参与调控木质素的积累过程。陈亮[18]研究发现,在丹参中过表达漆酶基因促进发根的生长,使得根系加粗,用显微镜观察丹参发根的横截面,看到木质部范围变大,染色后凯式带的紫色加深,说明漆酶加强了其木质化程度。漆酶基因在高等植物木质素合成中起着重要作用,其表达差异也会引起植物木质化结构表现差异。有研究[19-23]发现,当漆酶表达受到抑制时会减少木质素的积累,而木质素含量降低会影响植物木质化结构的表达,例如果核的残核和软核,利用该表达特点可进一步研究漆酶表达调控机制。本研究采用qRT-PCR技术分析基因表达模式,结果显示FtLAC-1和FtLAC-2在厚、薄果壳型苦荞不同组织器官中表达量不同,其中在种子的表达量最高。FtLAC-1和FtLAC-2在厚果壳苦荞不同组织器官的表达量均高于薄果壳苦荞,推测厚果壳木质素积累多与漆酶基因的高表达有一定的关系。经课题组前期转录组差异表达基因分析,薄果壳苦荞中与木质素生物合成途径相关的大部分基因表达下调,且在厚果壳中的表达量高于薄果壳苦荞,与本研究漆酶基因表达趋势相同,但有研究[24-25]发现在软籽石榴种皮的硬度发育中,漆酶基因会促进木质素合成积累,可以推断漆酶基因与苦荞果壳木质素合成代谢有紧密联系。但FtLAC-1和FtLAC-2基因在果壳不同组织器官表达模式不同,其在果壳木质素合成和积累中的具体功能还需进一步验证。
4 结论
从苦荞中克隆出2条漆酶基因FtLAC-1和FtLAC-2,生物信息学分析结果推测二者具有其他植物漆酶蛋白相似功能。qRT-PCR基因表达分析结果显示,FtLAC-1和FtLAC-2在厚果壳苦荞中的表达量高于薄果壳苦荞,推测其在薄果壳苦荞中低表达与薄果壳的木质素合成积累低有关,但具体功能还有待进一步验证。
