三江源区高寒草地植物多样性与土壤因子的耦合关系
2022-07-04杨冲王文颖周华坤毛旭锋马华清
杨冲,王文颖,周华坤,毛旭锋,马华清
(1.青海师范大学地理科学学院,青海 西宁 810008;2.青海师范大学生命科学学院,青海 西宁 810008;3.中国科学院西北高原生物研究所,青海 西宁 810008)
生物多样性是描述生物及其与环境彼此影响下所产生的生态系统和各种生态过程,是反映生物种群特征及状况的重要测度,也是生态系统的服务功能的重要指标[1]。在生物多样性的不同研究层次中,群落物种多样性是种群生态学研究的重要内容和方向。物种多样性是指物种在时空格局上的丰富性及排列特点[2],通过物种多样性指数可以了解生物群落的组织结构、稳定程度、发展阶段和功能效果[3],是生物多样性的具体表达和重要的组成部分。
土壤作为草地生态系统内物质交换和能量守恒的载体,是植物生长发育的物质基础[4],其理化性质能够控制和调节植物生长的生态过程,并对植物群落特征产生多尺度和多维度的影响[5]。土壤中营养元素的缺失会对植物的生长产生一定的消极作用,具体表现为对植物种子萌发、根系分布和微量元素的摄取有一定的抑制作用,进而成为草地植物群落更新与物种多样性的限制条件[6]。同时,植物群落的组成、密度及多样性的变化又会对土壤养分的有效性以及土壤结构的时空变异产生影响。不同的植物种类对土壤中元素的利用率不同,从而影响土壤的稳定性,不同的植物盖度以及凋落物成分的变化,也会对土壤的理化性质产生影响。植被与土壤系统是不可分割的整体,土壤因子与草地植物的群落组成、物种多样性格局之间的耦合关系以及互馈机制,一直是生态学研究的热门问题[7-9]。
三江源区位于青藏高原腹地,素有“中华水塔”之称,是我国重要的天然草场,也是生物多样性的资源宝库,具有重要的经济价值和生态功能[10-11]。近几十年来由于全球气候变化以及人类活动的影响,叠加三江源区高寒生态系统的脆弱性和难恢复性,形成大面积的次生裸地,造成表层土壤养分的大量流失,而土壤因子的变化又反过来影响该区高寒草地群落的物种组成和多样性[12]。目前,关于三江源区高寒草地植物物种多样性和土壤因子关系的研究,常采用定量与定性分析、相关性分析、多元线性回归等方法探究植物多样性与土壤因子之间的相互关系,这类方法的缺陷在于未能明确多种因子间相互作用程度,难以揭示植物群落正向演替的限制因子,也难以反映植物多样性与土壤因子间的耦合协调程度。鉴于此,本研究以三江源区不同类型高寒草地为试验对象,研究了不同类型高寒草地植物多样性特征和土壤理化性质状况,利用灰色关联度模型分析植物多样性与土壤因子之间的关联程度,并引入耦合度模型揭示植物多样性与土壤因子间的耦合协调程度,试图探究不同类型高寒草地的植物群落组成特征、植物群落多样性特征、土壤理化性质状况、影响植物多样性的主要土壤因子及植物多样性与土壤因子间的耦合协调程度,以期为该区草地资源的管理利用,退化草地的人工恢复提供科学依据。
1 材料与方法
1.1 研究区域概况
研究地点位于青海省三江源国家自然保护区,E 89°45'~102°23′,N 31°39'~36°12′,属于青藏高原腹地,平均海拔3 500~4 800 m,年温差小,日温差大,日照时间长,平均年日照射时数达到2 500 h以上,居全国前列,该区气候寒冷,一年只有冷暖两季,没有四季之分,冷季持续时间长达7~8个月,全年无绝对无霜期,属高寒大陆性气候;年降水量262.7~772.8 mm,多集中在5~10月,区内河流密布,湖泊、沼泽众多,是世界上海拔最高、面积最大、湿地类型最丰富的地区;草地类型主要以高寒草原、高寒草甸和高寒湿地为主,局部地区有人工草地分布,畜牧业为当地主要的生产方式,放牧家畜主要为藏羊和牦牛[11-13]。

图1 研究区示意图
1.2 样地设置
本研究中,共设置高寒草原(G1)、高寒草甸(G2)、沼泽草甸(G3)、人工草地(G4)4种不同类型的高寒草地(表1)。其中人工草地是高寒草甸极度退化后,人工翻耕种植垂穗披碱草的人工草地。播种时用磷酸二胺和羊板粪作基肥,磷酸二胺施用量150~300 kg/hm2,后期无施肥措施。人工草地建植后第1年到第2年的返青期绝对禁牧,之后作为冬季牧场,冬季牧场通常在当年10月至次年5月放牧,放牧家畜为牦牛和藏系绵羊。每个草地类型包括3个10 m×10 m的重复样地,重复样地彼此间尽可能使它们有相近的地形、植被和土壤类型(样地之间空间距离1~3 km,避免假重复),这样共计12个样地。G1草地在禁牧区(禁牧年限为7 a),无放牧。G2、G3、G4草地为冬季牧场。G1草地在玛多县,G2、G3、G4草地在玛沁县。

表1 样地详细信息
1.3 样品采集与处理
2017年8月中旬,在每个类型样地采用随机步程法设置3个样方[13],样方大小为1 m×1 m,对每个样方用全球定位系统(GPS)进行定位,并用植物样方调查表对植物群落进行调查,统计样方中的植物群落盖度、高度、出现的物种数、所有物种的分盖度、株高(每种植物随机测定5~10株的自然高度,然后取平均值)。然后将样方中的植被按禾本科、莎草科、豆科、杂类草分类齐地面剪下,用牛皮纸信封包装,带至实验室用超纯水清洗,去除泥沙粉尘等杂质,在60 ℃下烘干至恒质量后称量其地上生物量。在剪完植被的样方上用土钻采集土壤样品,采样深度为20 cm,将3个重复样品混合成一个样,用自封袋封装带回实验室,用孔径2 mm土筛剔除植物根系和石砾等杂物,将筛分出来的土样风干后分别过1 mm和0.15 mm土筛,用于土壤理化性质的测定。
1.4 测定项目与方法
土壤含水量和容重用烘干法测定,土壤pH用酸度计法测定,土壤全氮、全碳、有机碳(酸熏处理)通过Elementar Vario ELⅢ元素分析仪测定,土壤氨氮、硝氮、速效磷、全磷通过CleverChem Anna全自动间断化学分析仪测定,土壤速效钾、全钾通过岛津A6300原子吸收仪测定。
1.5 数据处理和统计分析
植物多样性指数[14]的计算包括:
(1)重要值
相对高度=样方内某一物种高度/样方内所有物种高度总和
相对盖度=样方内某一种物种盖度/样方内所有物种盖度总和
重要值=(相对高度+相对盖度)/2
(2)物种丰富度指数:R=S
(3)Shannon-Weaver指数:
(4)Simpson指数:
(5)Pielou均匀度指数(E):
E=H′/lnS
式中:Pi为群落中物种i的重要值,S为群落中的物种总数。
利用分析系统多因素交互作用的灰色关联度模型[15]计算关联系数,揭示植物多样性指数与土壤因子之间的耦合关系和协调程度。首先对数据进行区间标准化法无量纲化处理,然后计算关联系数。
(6)关联系数计算:
ξi(j,k)=
式中:ξi(j,k)为不同类型高寒草地第k个样本点的土壤指标i和植物群落多样性j的关联系数,ZiL(k)为土壤指标i的标准化值,ZjI(k)为植物多样性指标j的标准化值,ρ为分辨系数,在[0,1]内取值,通常取0.5[16]。
将关联系数按样本数量求得平均值,可以得到一个m×n的关联度矩阵γ,它能够从整体上反映单个土壤因子指标和单个植物多样性指标之间的关联程度。
(7)关联度计算:
式中:γ(i,j)为关联度,m为土壤因子指标数,n为多样性指标数,关联度评价标准见表2。
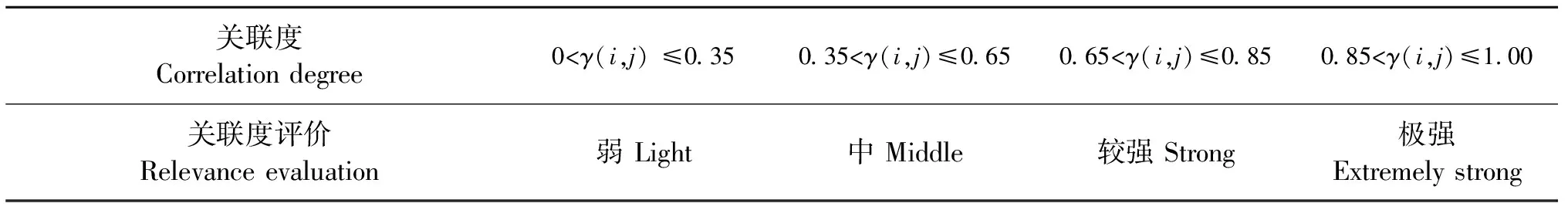
表2 关联强度标准
Extremely strong为从整体上定量评价不同类型高寒草地群落多样性与土壤因子的耦合协调发展程度,继续构建两者之间的耦合模型[17]。系统耦合协调程度评价标准见表3。

表3 系统耦合协调度标准
(8)耦合度(C)计算:
数据记录用Excel 2007完成,作图用ArcGIS、GraphPad Prism8完成,数据统计分析用SPSS 21完成。数据方差分析前均进行正态性和方差齐性检验,用方差分析(ANOVA,LSD)比较不同处理间差异显著性。用平均值和标准差表示测定结果,用标准差与原始数据平均数的比值计算变异系数,用MATLAB 2017计算关联度和耦合度。
2 结果与分析
2.1 不同高寒草地群落物种组成特征
由表4和表5可知,在4个草地中共有植物47种,分为15科34属,禾本科和菊科物种数相同,均为6属7种,莎草科为3属7种,毛茛科为4属6种,龙胆科有3属6种,玄参科为2属3种,唇形科为2属2种,豆科为1属2种,蔷薇科、报春花科、虎耳草科、堇菜科、伞形科、百合科、泥炭藓科均为1属1种。G1草地由4科5属共5种植物组成,占研究区物种数的10.6%,紫花针茅和垂穗披碱草为主要优势种群,重要值分别为44.49和40.32;G2草地由11科14属共22种植物组成,占研究区物种数的46.8%,矮嵩草和线叶嵩草为主要优势种群,重要值分别为15.4和14.72;G3草地由13科17属共20种植物组成,占研究区物种数的42.6%,藏嵩草和华扁穗草为主要优势种群,重要值分别为29.78和9.06;G4草地由11科24属共29种植物组成,占研究区物种数的61.7%,短芒洽草和甘肃马先蒿为主要优势种群,重要值分别为13.82和12.73。

表4 群落物种组成及重要值
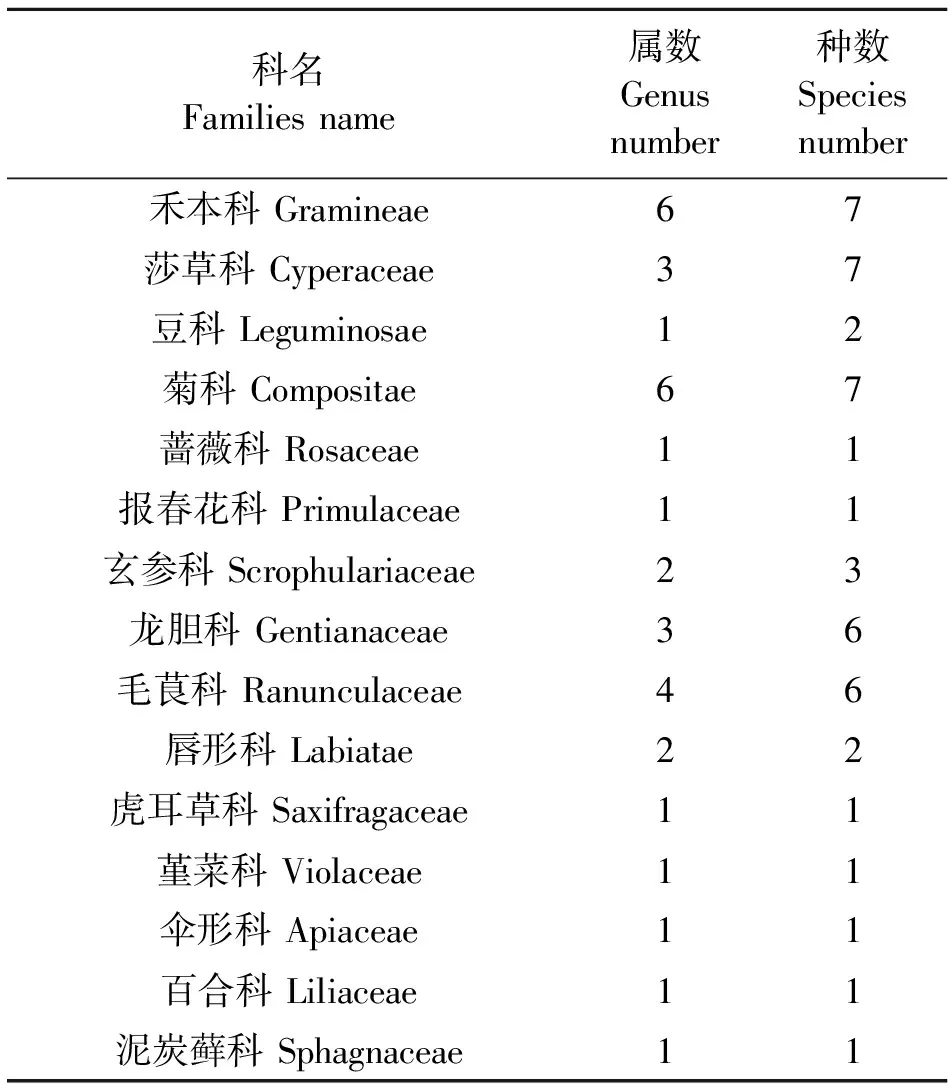
表5 调查草地植物种类组成
2.2 不同高寒草地地上生物量及功能群变化特征
由表6可知,不同功能类群植物的地上生物量在各草地之间有较大差异,其中,禾本科植物地上生物量范围为27.25~205.93 g/m2,G1草地禾本科植物生物量最高,与其他草地差异显著(P<0.05),G4草地禾本科植物生物量较高,与其他草地差异显著(P<0.05),G3与G4草地禾本科植物生物量最低,相互之间差异不显著。莎草科植物地上生物量范围为0.69~475.25 g/m2,G3草地莎草科植物地上生物量最高,且与其他草地差异显著(P<0.05),G3草地莎草科植物地上生物量较高,且与其他草地差异显著(P<0.05),G1与G4草地之间差异不显著,且显著低于G2与G3草地。豆科植物地上生物量在各草地普遍较低,变化范围为0.15~15.96 g/m2,各草地彼此之间差异不显著。杂类草地上生物量为6.39~178.67 g/m2,G4草地杂类草地上生物量最高,与G2草地差异不显著,与G1和G3草地差异显著(P<0.05),G1与G2草地之间差异显著(P<0.05),G2与G3草地之间差异不显著,G1与G3草地之间差异不显著。各草地总的地上生物量范围为213.16~581.11 g/m2,G3与G2草地地上生物量较高,两者差异不显著,但均显著高于G1草地,G1与G4草地地上生物量较低,且两者差异不显著,各草地地上生物量高低为G3>G2>G4>G1。

表6 各功能群及总的地上生物量
2.3 不同高寒草地植物群落结构与物种多样性特征
2.3.1 群落高度和盖度对不同高寒草地的响应 由图2可知,不同高寒草地植物群落的高度、盖度外貌特征存在差异,群落高度变化范围为8.5~25.4 cm,G4草地群落高度最高,与G3草地差异不显著,G1与G2草地群落高度较低且两者差异不显著,G3、G4草地分别与G1、G2草地差异显著(P<0.05)。群落盖度的变化范围为66.7%~94.3%,G2草地群落盖度最高,G2、G3、G4草地彼此之间差异不显著,G1草地群落盖度最低,与其他草地之间差异显著(P<0.05)。总体上来讲,研究区不同高寒草地的群落高度差异较大,群落盖度的差异较小,G2草地群落高度较低,盖度最高,植被多呈现垫状或匍匐类型。

图2 群落高度和盖度变化
2.3.2 物种多样性特征对不同高寒草地的响应 生物多样性是群落的重要特征,是生态系统功能维持的生物基础。本研究以Shannon-Weaver指数、Simpson指数、物种丰富度指数、Pielou均匀度指数来研究植物群落物种多样性,物种丰富度指数为每块草地中的物种数。由图3可知,植物群落物种数变化范围为5~29种,均值为19种,变异系数为0.53,G4草地物种数最多为29种,G2草地次之,G1草地物种数最少仅5种;Shannon-Weaver指数变化范围为1.15~2.82,均值为2.29,变异系数为0.34,G4草地Shannon-Weaver指数最高,G2、G3草地Shannon-Weaver指数较高,G1草地Shannon-Weaver指数最低;Simpson指数变化范围为0.63~0.92,均值为0.83,变异系数为0.17,G4草地Simpson指数最高,G2、G3草地Simpson指数较高,G1草地Simpson指数最低;Pielou均匀度指数变化范围为0.71~0.88,均值为0.81,变异系数为0.09,G2草地Pielou均匀度指数最高,G3和G4草地Pielou均匀度指数较高,G1草地Pielou均匀度指数最低。
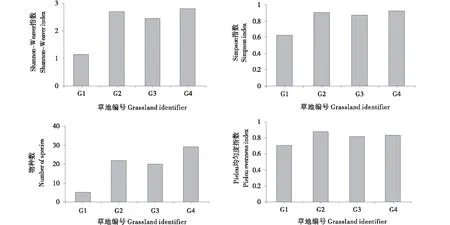
图3 物种多样性特征
2.4 不同高寒草地土壤理化性质特征
为了进一步进行耦合度计算,求取各草地0~20 cm土层3次重复后不同理化指标的均值,结果如表7所示。

表7 不同高寒草地土壤理化性质特征
2.5 不同高寒草地植物多样性与土壤因子的灰色关联度分析
将12个土壤因子(pH、容重、含水量、有机碳、全碳、氨氮、硝氮、全氮、速效磷、全磷、速效钾、全钾)共线检验后作为子序列,将3个植物物种多样性指数作为母序列,与4个不同类型高寒草地进行灰色关联度分析,分辨系数取0.50,结合关联系数计算公式计算出关联系数值。由表8可知,各指标的关联度在0.758~0.895,说明植物多样性与土壤因子之间存在较强的耦合作用。为进一步分析植物多样性对土壤因子的反馈作用以及土壤因子中对植物多样性的主要影响因子,将不同植物多样性指数对土壤因子的关联度值进行简单平均,得到各个土壤因子与植物多样性指数间的关联度值。由表9可知,Shannon-Weaver指数、Simpson指数和Pielou均匀度指数与土壤因子的关联度分别为0.825、0.817、0.818,说明土壤因子对Shannon-Weaver指数的影响较大,对Simpson指数的影响略小。在12个土壤因子中pH、容重、硝氮、全氮、速效磷、全磷对植物多样性影响较大,且关联度均值都超过0.9,是影响植物生长与分布的关键因子。

表8 不同高寒草地植物物种多样性指数与土壤因子关联度矩阵

表9 植物群落物种多样性指数与土壤因子耦合矩阵
2.6 不同高寒草地植物多样性与土壤因子耦合协调度分析
由表10可知,不同类型高寒草地植物群落多样性与土壤因子的耦合度范围为0.167~0.762,平均耦合度为0.553,G1草地耦合度为0.167,处于严重不协调状态,G2草地耦合度为0.683,处于弱协调状态,G3草地耦合度为0.762,处于中度协调状态,G4草地耦合度为0.599,处于轻度不协调状态。

表10 不同类型高寒草地植被与土壤系统耦合协调状况评判结果
3 讨论
3.1 不同类型高寒草地植物群落组成和地上生物量特征
每种植物都有其独特的结构和功能,群落中的种类组成是形成群落的结构基础和群落的重要特征,对生态系统的稳定性、生产力和营养动态等功能有着重要意义[18]。本研究中,三江源区不同类型高寒草地植物群落物种构成表现为“多数种属于少数科、少数种属于多数科”,这与张建贵等[19]在祁连山的研究结果相一致。三江源区高寒草原相对于其他高寒草地由于生境恶劣,特别是土壤养分和土壤水分的限制,造成种间竞争激烈,可能是物种数较少的原因,这与杜岩功等[20]在玛多县的研究结果一致。
植物群落生产力水平是生态系统结构和功能的重要表现形式,是植物生态学特性和外界环境条件共同作用的产物,地上生物量是衡量草地植物生长发育、生产力大小和生态系统服务价值的重要指标,代表了生态系统利用物质和能量的能力[21]。本研究中,禾本科、莎草科、豆科、杂类草4种高寒草地植物功能群,在不同的高寒草地中地上生物量大小存在一定差异,在沼泽草甸和高寒草甸中,群落地上生物量主要取决于莎草科地上生物量,在人工草地和高寒草原中,群落地上生物量主要取决于禾本科地上生物量,豆科在各高寒草地中的生物量均较小,另外,人工草地建植5 a后,优良禾本科牧草以及总的地上生物量都大幅降低,杂类草比例明显较高,这与王彦龙等[22]在果洛的研究结果相一致,因此,作为牧用的人工草地必须进行人工调控管理。
3.2 不同类型高寒草地植物群落物种多样性特征
生物多样性是生态系统的基本特征,也是草地生态系统健康诊断的主要依据,物种多样性指数是物种水平上多样性和异质性程度的度量,能综合反映群落物种丰富度和均匀度的总和。生物多样性本身不是一个独立变量,其维持受到多种因素的影响[23]。本研究中,沼泽草甸相较于高寒草甸、人工草地来说,其Shannon-Weaver指数、Simpson指数、Pielou均匀度指数都较低,可能的原因是沼泽草甸处于湿水环境且盖度较高根系致密,并能发育出草毡表层,外来物种很难入侵和生存,植物群落的优势种藏嵩草种群在空间上占有明显的生态位优势,使群落物种丰富度指数及均匀度指数下降,从而形成生物多样性指数较低的格局;人工草地的物种丰富度指数和Shannon-Weaver指数最高,但是其Pielou均匀度指数却低于高寒草甸和退化高寒草甸,显示出群落物种的丰富度、Shannon-Weaver指数与Pielou均匀度指数并不一致,这与刘忠宽等[24]在锡林河的研究结果相一致,可能是因为群落物种的丰富度和多样性程度越大,物种对生境的分割程度越高,因而导致群落物种均匀度的降低。
3.3 不同类型高寒草地植物多样性与土壤因子的耦合关系
植物多样性对于三江源区高寒草地的保护和恢复有重要意义,不同植物种适应高寒环境和脆弱生态系统的能力不同,群落中多样化的植被对于环境条件的动态变化和人为干扰有着较强的适应能力[25]。植被特征、土壤理化性状是决定不同类型高寒草地植被土壤系统稳定性的主要影响因素,生态恢复过程中植被群落与土壤系统的耦合效应研究己经成为目前生态恢复评价的一个重要研究方向。植被群落多样性与土壤因子的关联机制复杂,对于影响植物多样性的土壤因子,不同的学者研究结果有所不同。有学者[26-28]认为,土壤有机碳和全氮与植物多性指数会表现出较高的关联性,是影响群落多样性的重要因子,也有学者[29-30]认为土壤有机碳对群落多样性的影响程度较低。有学者[31-32]认为,土壤全钾和速效钾与群落多样性无关,也有学者[33-35]认为土壤全钾和速效钾是影响植物多样性的主导因素。本研究中,土壤因子与Shannon-Weaver指数、Simpson指数和Pielou均匀度指数关联度分别为0.825、0.817、0.818,属于较强关联,这说明土壤理化性质的改善对于植被的恢复有重要意义。土壤硝氮、pH、速效磷、全氮、容重、全磷对植物多样性影响较大,影响程度依次增高,关联度均值都超过0.9,是影响植物生长与分布的关键因子。
耦合度是指两个或两个以上的系统或要素之间通过自身和外界的各种相互作用而彼此影响的现象,是评价协调状况好坏的定量指标。将耦合的概念引入到生态学研究领域具有重要的创新意义,它强调的是一种深层次的因子之间的关系及过程的研究,为揭示生态系统的协同、约束关系提供了理论基础[36]。本研究中,耦合协调度的结果表明,4类高寒草地植物多样性与土壤因子的耦合协调度总体上处于较低水平。其中,高寒草原处于严重不协调状态,人工草地处于轻度不协调状态,高寒草甸处于弱协调状态,沼泽草甸处于中度协调状态。高寒草原海拔相对较高,生境严酷,土壤有沙化趋势,植物受土壤养分和水分的双重限制,植被稀疏低矮,生态系统异常脆弱,可能是导致植物多样性与土壤因子的耦合协调度处于严重不协调状态的原因。人工草地处于轻度不协调状态,可能是因为单播人工草地很容易受到外来草种的入侵,导致高寒牧区多年生人工草地普遍存在建植3~5 a后急速退化的现象。
4 结论
研究区高寒草地植物种类组成简单且数量稀少,共有草本植物15科34属47种,以禾本科、菊科、莎草科、毛茛科、龙胆科为主,各高寒草地地上生物量大小具体表现为沼泽草甸>高寒草甸>人工草地>高寒草原。
物种丰富度指数、Shannon-Weaver指数和Simpson指数的具体表现为人工草地>高寒草甸>沼泽草甸>高寒草原,Pielou均匀度指数的具体表现为高寒草甸>人工草地>沼泽草甸>高寒草原。
植物多样性与土壤因子关联度较强,土壤pH、容重、硝氮、全氮、速效磷、全磷是影响植物生长与分布的关键因子,4类高寒草地植物群落多样性与土壤因子的耦合协调度整体上处于较低水平。
