亚洲树轮缺轮率的空间分布特征及其影响因素
2022-06-30王春松周非飞郑壮鹏董志鹏邴晓瑾
王春松,周非飞,郑壮鹏,董志鹏,邴晓瑾
(福建师范大学 a.地理科学学院,b.湿润亚热带山地生态国家重点实验室培育基地,福州 350007)
0 引言
亚洲地形起伏大、地貌结构复杂,形成了多样的气候类型,巨大的季风-干旱气候系统深刻影响着地理环境[1]。树木生长受环境影响,树轮宽窄变化能反映环境变化特征,树轮年代学正是基于此点经交叉定年而发展成为一门学科[2]。树木年轮因其具有定年精确、时间分辨率高、空间分布广等特点而广泛运用于生态学与古气候学领域[3]。树轮是形成层细胞生长与分化的结果,一般形成层对外界环境较敏感,当环境突变或发生异常气候事件,树木形成层生长异常而形成诸如伪轮、霜轮、缺轮等异常轮现象[4-5]。基于异常轮的研究,可以重建历史上的干旱、火山喷发、虫灾等自然灾害事件[6-7]。
树木整个生长季气候如果都很恶劣,就可能在某一年或数年不产生年轮,即缺轮[8]。树木在寒冷、干旱区更易发生缺轮[6,9],但在温暖湿润区缺轮现象亦有出现。这说明缺轮不仅受气候因素调控,树种、树龄等生理因素对缺轮也有重要影响。邵雪梅等研究表明华山地区降水与树木径向生长有显著的正相关关系[9],陈诗音等研究表明福建地区不同树龄马尾松径向生长存在差异[10],Liu等研究表明在降水较少的低海拔地区黄山松(Pinustaiwanensis)径向生长受季节性干旱影响更大[11]。这些研究为缺轮记录重建干旱事件提供了生理上的解释。Liang等分析祁连圆柏(Juniperusprzewalskii)缺轮记录重建了中国中西部大面积干旱事件[6],将缺轮记录重建干旱事件的方法运用于实践。不同树龄间树木缺轮现象差异明显,Novak等研究表明地中海松(Pinushalepensis)缺轮率随树龄的增加而逐渐上升[13]。树木存在一定的生态弹性去抵抗环境胁迫并从中恢复,树种间生态弹性差异导致其应对极端气候事件的策略存在不同。Fang等指出青藏高原地区的西藏冷杉(Abiesspectabilis)和大果圆柏(Juniperustibetica)对干旱事件的响应策略不同是导致两者生长差异的原因[12]。
当前研究在空间上仅考虑小生境或局域尺度,研究对象也仅限于单个或几个树种,针对大空间尺度的多树种缺轮现象的研究存有空白。本研究使用国际树轮库中亚洲树轮缺轮记录,分析气候因子与生理因子对树木缺轮的影响,试探讨不同区域树木缺轮现象对不同要素的响应,以阐明亚洲缺轮记录的空间分布特征,为之后开展大规模基于缺轮的重建工作提供数据支持与科学依据。
1 数据与方法
1.1 数据来源
1.1.1 树轮数据
树轮宽度序列在树轮研究中运用最久、发展最完善,且树轮宽度相较于密度、同位素等指标,获取简单、空间分布广且密集,为更好地反映空间上的分布,本研究采用树轮宽度原始序列数据[14]。在国际树轮数据库中下载亚洲地区589个点的树轮宽度数据,并记录经纬度、树种等基本信息。每棵树木的树龄无法精准确定,但年表时间跨度基本可替代树龄,此方式会高估树木年龄,但并未对研究结果产生实质影响,因此选用年表长度代替树龄。
1.1.2 气象数据
气象数据使用Climatic Research Unit(CRU)数据集CRU TSv.4.04中全球陆地温度、降水0.5°×0.5°的月平均数据,在KNMI (https://climexp.knmi.nl/start.cgi)网站中下载。使用临近采样点最近的一个格点的1901—2000年年均温、降水序列数据来代表采样点气象数据。选取该时段温度与降水是因为多数年表在2000年左右截止,气象数据起始时间在1901年,所以该时段的温度、降水能较好地反映各树轮样本生长环境气候特征。
1.2 研究方法
缺轮率可直接在COEECHA结果中读出,具体值是年表中的总缺轮数与总轮数的比值,计算公式如下:
MRR=MR/ATR
(1)
式(1)中:MRR(Missing ring rate)代表某点年表缺轮率,MR(Missing ring)代表某点年表中总缺轮数量,ATR(All tree ring)代表年表总年轮数量。使用MATLAB 2020、SPSS 23.0进行数据处理与分析,使用Arcmap 10.2、Origin 2018绘图。
2 结果与分析
2.1 亚洲树轮缺轮空间分布特征
亚洲树轮缺轮记录在东北亚地区(151条)主要分布在俄罗斯北部(66条)、中国(50条)、蒙古(30条)等地,在中南亚地区(95条)集中分布在尼泊尔(45条)、巴基斯坦(24条)、不丹(19条)等地,东南亚地区(14条)缺轮记录较少且均匀分布在各国家,在西亚地区(34条)分布在土耳其(24条)、黎巴嫩(5条)等地(表1)。亚洲树轮缺轮记录主要分布在高纬(俄罗斯北部)、高海拔(青藏高原)低温区,中亚、中东等干旱区。但是有些缺轮也出现在东南亚与中国东南部等温度高、降水多的地区(图1)。

表1 亚洲各地区树轮及缺轮年表数Table 1 The chronology of tree ring and missing ring in Asia
2.2 气候因子对缺轮率的影响
因样本间缺轮率相差较大,以中位数为界,将样本分为高缺轮率(MRR>中位数1.64‰)、低缺轮率(MRR<中位数1.64‰)两组,探讨温度、降水对不同缺轮率段的影响。以年降水量200 mm为间隔,得出亚洲树轮缺轮率降水量的梯度区间分布。在总缺轮率、高缺轮率、低缺轮率3组中,80%的样本都在降水1 000 mm区间内,且缺轮率峰值都出现在200~400 mm年降水区间。以年降水量1 000 mm为界,总缺轮率组和高缺轮率组在小于1 000 mm区间内缺轮率随降水量增加而减少,在大于1 000 mm区间内随降水量增加而增多,在低缺轮率组中无上述现象(图2a、2c、2e)。在降水量小于1 000 mm的缺轮样本中,总缺轮率组缺轮率随降水量每增加100 mm而减少0.91‰(P<0.05)(图3a);低缺轮率组缺轮率随降水量每增加100 mm减少0.03‰(图3c);高缺轮率组缺轮率随降水量每增加100 mm减少1.22‰(P<0.05,图3e)。
以年平均气温5℃为间隔,得出亚洲树轮缺轮率的温度梯度区间分布。缺轮率的温度区间分布并未出现与降水量区间相似的分布特征,总缺轮率、高缺轮率、低缺轮率3组样本的缺轮率峰值并不一致。总缺轮率组峰值出现在-15℃以下区间,次峰值在-5~0℃区间;高缺轮率组峰值出现在0~5℃区间,次峰值在-5~0℃区间;低缺轮率组峰值出现在30℃以上区间,次峰值在0~5℃区间。以年均温0℃为界,3组样本缺轮率随温度升高而增加,大于0℃区间内3组样本缺轮率与年均温无明显趋势关系(图2b、2d、2f)。在年均温小于0℃的样本中,年均温每上升10℃,总缺轮率组缺轮率增加2.35‰(图3b),低缺轮率组增加0.36‰(P<0.05)(图3d),高缺轮率组增加3.56‰(图3f)。
2.3 生理因子对缺轮率的影响
亚洲树轮平均缺轮率为3.94‰,树种间的轮率差异较大,其中祁连圆柏、岷江柏木、西伯利亚落叶松的平均缺轮率较大,分别为22.51‰、8.85‰、5.99‰;而高加索冷杉、橡木、榆树的平均缺轮率较小,仅分别为0.15‰、0.2‰、0.36‰。松科与柏科的树种缺轮率较高,缺轮率大于平均缺轮率的树种分别有祁连圆柏、岷江柏木、西伯利亚落叶松、西藏白皮松、兴安落叶松。对比各树种缺轮率与年降水、年均温之间的关系,缺轮率较高的树种多在年降水量800 mm以下地区(表2)。不同树种缺轮率与降水的相关性差异明显,其中大果圆柏、云南铁杉缺轮率与年降水量显著正相关,欧洲赤松缺轮率与年降水量显著负相关(图4b)。
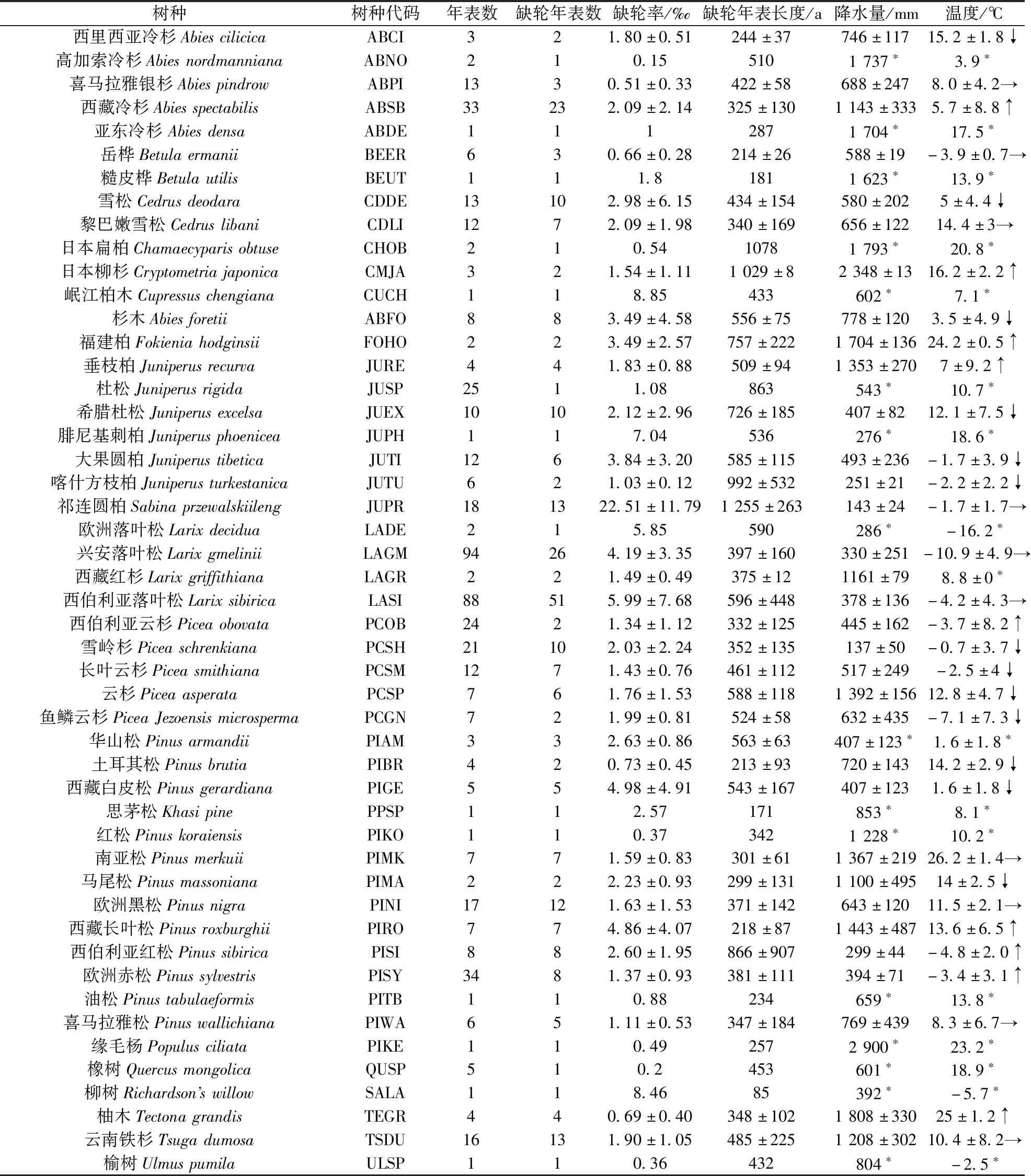
表2 亚洲缺轮树种基本信息Table 2 Basic information of species with missing ring in Asia
以100年为间隔得到缺轮率年龄段梯度分布,亚洲缺轮记录主要集中在300~600年的年龄区间。树轮缺轮率随树龄增加呈上升趋势,但100~200年树龄段的缺轮率却高于200~300年树龄段的缺轮率(图5)。通过查找各树龄段树种发现,100~200年树龄段主要是兴安落叶松、西藏长叶柏、西藏冷杉等缺轮率较高树种,而200~300年龄段主要是雪岭杉、欧洲黑松、云南铁杉、柚木等缺轮率较低的树种。亚洲多数树种缺轮率与树龄呈现正相关关系,其与中西伯利亚红松呈显著正相关关系,与兴安落叶松呈极显著的正相关关系(图5a)。
3 讨论
3.1 气候因子对缺轮率的影响分析
总体上看亚洲地区树轮在干旱区更易产生缺轮,降水对树木径向生长的限制作用较温度更为明显。干旱区气候干燥,较少的降水无法有效补给土壤水分,树木生长过程中的水分需求不能得到满足,使得降水成为干旱区树木生长的限制因素[10]。当降水量较往年减少时,树木形成层细胞生长、分裂过程受阻导致该年的树轮宽度变窄[16]。若遇上极端干旱年,树木形成层细胞停止生长而产生缺轮,所以在干旱区树木的缺轮率随降水减少而增加。而在年降水量充沛的湿润地区,树木缺轮可能是季节性的干旱事件所引起的。在形成层细胞生长、分裂初期,树木生长对水分的需求大大增加,若此时水分不足,形成层细胞生长、分裂停滞[16-17],从而造成缺轮现象。土壤水是植物生长直接可利用的水分,土壤含水量除直接受降水影响外,还受蒸发的控制。气温上升导致潜在蒸发力越强,使得土壤含水量下降,树木生长可利用的水分就会减少,进而造成树木径向生长停滞。温度通过蒸发改变土壤含水量进而影响树木缺轮率,这种间接影响可能是造成温度较降水对缺轮率影响不显著的原因。根据IPCC[18]指出,当前全球气温都有不同程度的上升,降水量呈现差异化。已有的研究表明在中亚[19]、青藏高原、中国东南地区[20]有不同程度的干旱化。结果也显示缺轮率随温度上升、降水减少有上升的趋势,故在全球暖干化下,该区域的树木径向生长更加受限[21],树木缺轮现象可能会更加频发。
3.2 生理因子对缺轮率的影响分析
缺轮率随树龄的增加而增大,该结论与前人研究较为一致[22-23]。树木随年龄的增加其高度增高、体积加大,树干内运送水分阻力不断增加,从而使得老龄树更易受到水分胁迫,导致所受到降水的限制作用增强[23]。树龄较老的树叶其光合容量较低,木质部组织生长增加其自身呼吸而消耗更多的能量,使得老龄树营养物质来源减少、能量消耗增加,导致用以树木次级生长的营养物质大大减少,从而限制树木径向生长[24]。老树相较于年轻的树可能具有更保守的生长策略,为防止不良气候条件的干扰,其形成层细胞在水热条件适宜的月份开始分裂且细胞生长速率较慢[25],在极端气候年份,这种保守型生长策略可能导致其径向生长停滞。大果圆柏与欧洲赤松其生长环境的类似,但其缺轮率与降水的相关性差异巨大。《中国植物志》中描述欧洲赤松为喜光性强、深根性树种,能适应土壤水分较少坡地,以及较干旱砂地及石砾砂土地区。在降水较多时,使得土壤水分过多,进而限制欧洲赤松根部呼吸,抑制其生长;且欧洲赤松喜阳,降水量增加导致光照减少,又进一步抑制其生长,上述影响可能是欧洲赤松缺轮率与降水量呈正相关关系的原因。缺轮现象可能是树木在极端干旱事件中的保护机制,通过减少树木组织生长所消耗的养分,使得更多营养物质分配到植物维持自身的生命活动中,各树种调节该过程能力的不同可能是缺轮率产生差异的原因[26]。Fang 等[12]对比青藏高原地区的杜松和西藏冷杉,发现相较于西藏冷杉,杜松面对干旱事件会花费较少的时间来降低水分需求,以更快地适应干旱状态。
