西藏8种灌木光合特性研究*
2022-06-29南吉斌旦增尼玛单增罗布李炳章
南吉斌,旦增尼玛,单增罗布,李炳章
(西藏自治区林木科学研究院,西藏 拉萨 850000)
绿色植物通过光合作用产生有机物,以供植物生长发育,光合作用是植物生命过程中重要的组成部分,是研究全球碳平衡的重要方式[1]。植物光合作用能力的强弱直接影响植物的生长,而光合指标是植物光合作用的直接反应[2],其随环境的变化而变化,在一定程度上可反应植物适应环境的能力和物种的丰富度,也是植物引种栽培及选择抗旱造林树种的重要依据[3-4]。森林是国家宝贵的自然资源,对改善生态、维护环境平衡起着至关重要的作用,而灌木是森林的重要组成部分之一[5]。
西藏气候恶劣、昼夜温差大、辐射强、干旱,导致部分地区主要的自然植被类型以灌木为主,其分布面积较大[6-7]。近年来由于全球气候变暖,过渡放牧导致西藏土壤沙化越来越严重,因此筛选适应性强的乡土树种已成为西藏植被恢复工程的重点[8-9]。目前。针对灌木的研究主要集中在物种多样性[10]、群落结构[11]、森林资源[12]、碳汇量及造林配置等方面[13-14]。但就西藏乡土灌木光合特性的比较研究未见报道。本研究主要以西藏主要的8种乡土造林灌木幼苗为试验材料,利用Li-6800 便携式光合仪,通过测定其植物叶片的光合及叶绿素荧光指标,并应用叶子飘等[15]直角双曲线修正模型对光响应及叶绿素荧光响应曲线进行估算,分析西藏8种灌木光合特性差异,探讨其光响应及叶绿素荧光响应技术,为拉萨河流域抗旱造林树种选择提供技术参考。
1 材料和方法
1.1 试验地概况及材料来源
试验地位于西藏自治区林木科学研究院苗木基地,地处拉萨河流域,温暖干燥,太阳辐射强,空气稀薄,昼夜温差较大,属于高原温带半干旱季风气候,年平均气温为9.8 ℃,年平均降水量468.1 mm,降水一般集中在6—9月,年日照时数长,全年无霜期100~120 d。
试验材料为2 a生实生幼苗,于2021年6月中旬,选取长势良好、高度基本一致,健康无病虫害的西藏8种灌木幼苗(表1),统一定植在规格为15 cm×21 cm的营养钵中,基质以腐殖质、珍珠岩、沙黏土按1︰1︰3的体积比配置,每钵种植1株幼苗,每个树种分别种植在10盆营养钵中,统一放在西藏自治区林木科学研究院炼苗棚中,进行统一的水分、养分管理。

表1 8种灌木种类
1.2 指标测定
1.2.1 生长指标测定
每个种选取生长健康的3株幼苗,共计24株苗木,于2021年7月初开始测量苗高、地径。苗高用钢卷尺测量,精确到 0.01 cm。地径用游标卡尺测量,精确到 0.001 mm。
1.2.2 SPAD值测定
采用SPAD-520(柯尼卡美能达公司,日本)叶绿素计进行测定。每个种随机选择健康无病虫害的3株幼苗,每株选择方向朝南的上中下各3片叶子,每个叶片测量3个点取平均数。
1.2.3 光合及叶绿素荧光指标测定
(1)光响应曲线和叶绿素荧光曲线测定 用Li-6800便携式光合仪(6800-01A,Li-Cor公司,美国)多相闪光 Multiphase FlashTM 光合荧光有叶室,选择晴朗的天气,于每天9:00—12:00进行测定。开始测前设定诱导光强(PAR)为1 000 μmol/(m2·s),流速500 m/s,温度20 ℃,CO2浓度400 μmol/mol,光合有效辐射梯度2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、80、50、0 μmol/(m2·s),重复6次。
(2)各光合和叶绿素荧光参数 用 Li-6800便携式光合仪于每天9:00~12:00进行测定。开始测前设定流速500 m/s,温度20 ℃,CO2浓度400 μmol/mol,光合有效辐射(PAR)为1 000 μmol/(m2·s),等待稳定后记点,测定指标有净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)、电子传递速率(ETR)、实际光化学速率(YpsⅡ),重复6次。
(3)Fo、Fm、Fv/Fm测定 用Li-6800便携式光合仪测定,测定前对叶片进行暗反应30 min,测定暗反应下最小初始荧光(Fo)、最大荧光(Fm)和暗反应下光系统Ⅱ最大光化学效率(Fv/Fm),重复6次。
1.3 数据统计与处理
1.3.1 光合参数计算
用叶子飘等[15]的光合计算软件 4.1.1 中的直角双曲线修正模型对光响应和叶绿素荧光响应曲线进行拟合,并计算表观量子效率(α)、最大净光合速率 (Pnmax)、饱和光强(Isat)、光补偿点(Ic)、 暗呼吸速率(Rd)、最大电子传递速率(Jmax)等,方程如下 :
(1)光响应[15]
式中:Pn为净光合速率,α为表观量子效率,I为光量子通量密度;β、γ为系数,Rd为暗呼吸速率。
(2)叶绿素荧光响应[16]
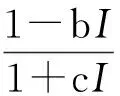
式中,J为电子传递效率,A为叶绿素荧光曲线的初始斜率,I为光量子通量密度,b、c为系数。
1.3.2 数据处理
利用 Excel 2010进行数据整理,用Origin 2017作图,利用SPASS.21 软件进行单因素方差分析和多重比较。
2 结果与分析
2.1 生长指标及SPAD值比较
西藏8种灌木苗高、地径和SPAD值见表2。由表2可知,8种灌木苗高和SPAD值P值分别是0.001、0.000,均小于0.05,说明8种灌木苗高、SPAD值之间差异显著,8种灌木苗高范围为26.410~30.047 cm,地径的范围为2.787~4.957 mm,SPAD值范围为32.467~49.900之间。8种灌木中,金露梅(Potentillafruticosa)苗高最高(30.047 cm),江孜沙棘(Hippophaerhamnoidessubsp.gyantsensis)次之(28.553 cm),皱叶醉鱼草(Buddlejacrispa)苗最小(26.410 cm);金露梅地径最高(4.957 mm),钝叶栒子(Cotoneasterhebephyllus)次之(4.660 mm),拉萨小檗(Berberishemsleyana)苗最小(2.787 mm);钝叶栒子SPAD值最大(49.900),拉萨黄花木次之(45.367)皱叶醉鱼草最小(32.467)。
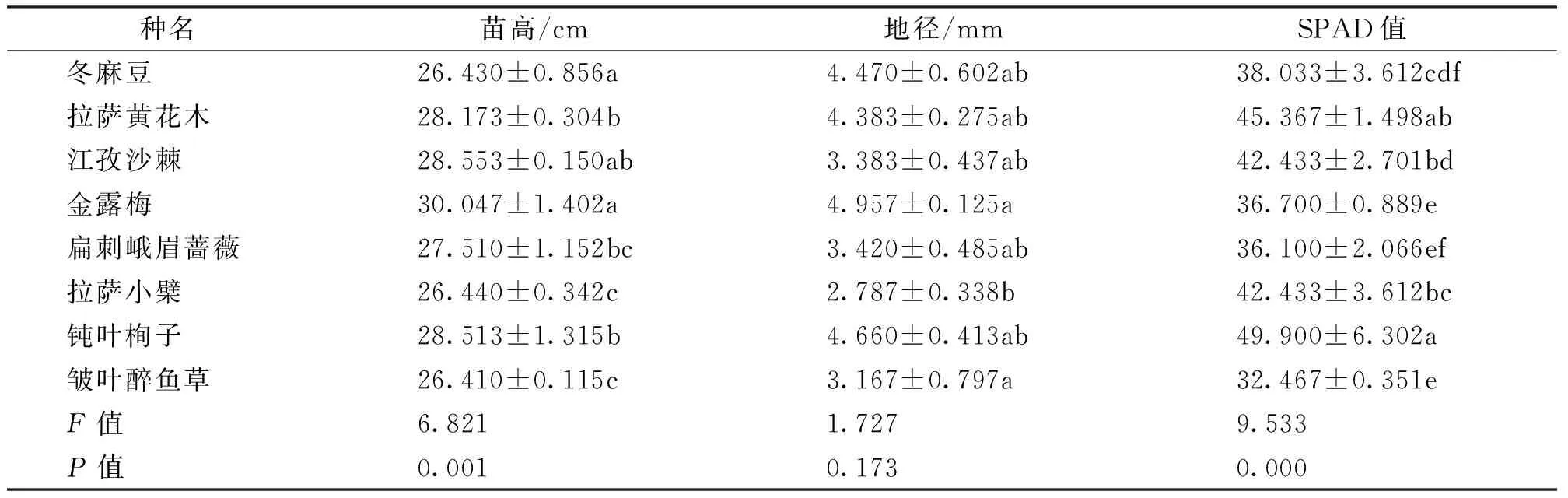
表2 8种灌木生长指标及SPAD值
2.2 净光合速率-光合有效辐射曲线及光合拟参数比较
8种灌木Pn-PAR曲线(图1)。由图中可知,8种灌木Pn-PAR曲线变化趋势一致,总体分4个阶段,第一阶段是随着PAR的增加,Pn呈直线上升趋势;第二阶段是当PAR继续增加时,Pn呈上升幅度逐渐减小;第三阶段是当PAR增加到光饱和点时,随着PAR的增加呈趋于稳定状态;第四阶段是随着PAR增大,Pn逐渐呈下降趋势,出现光抑制现象。
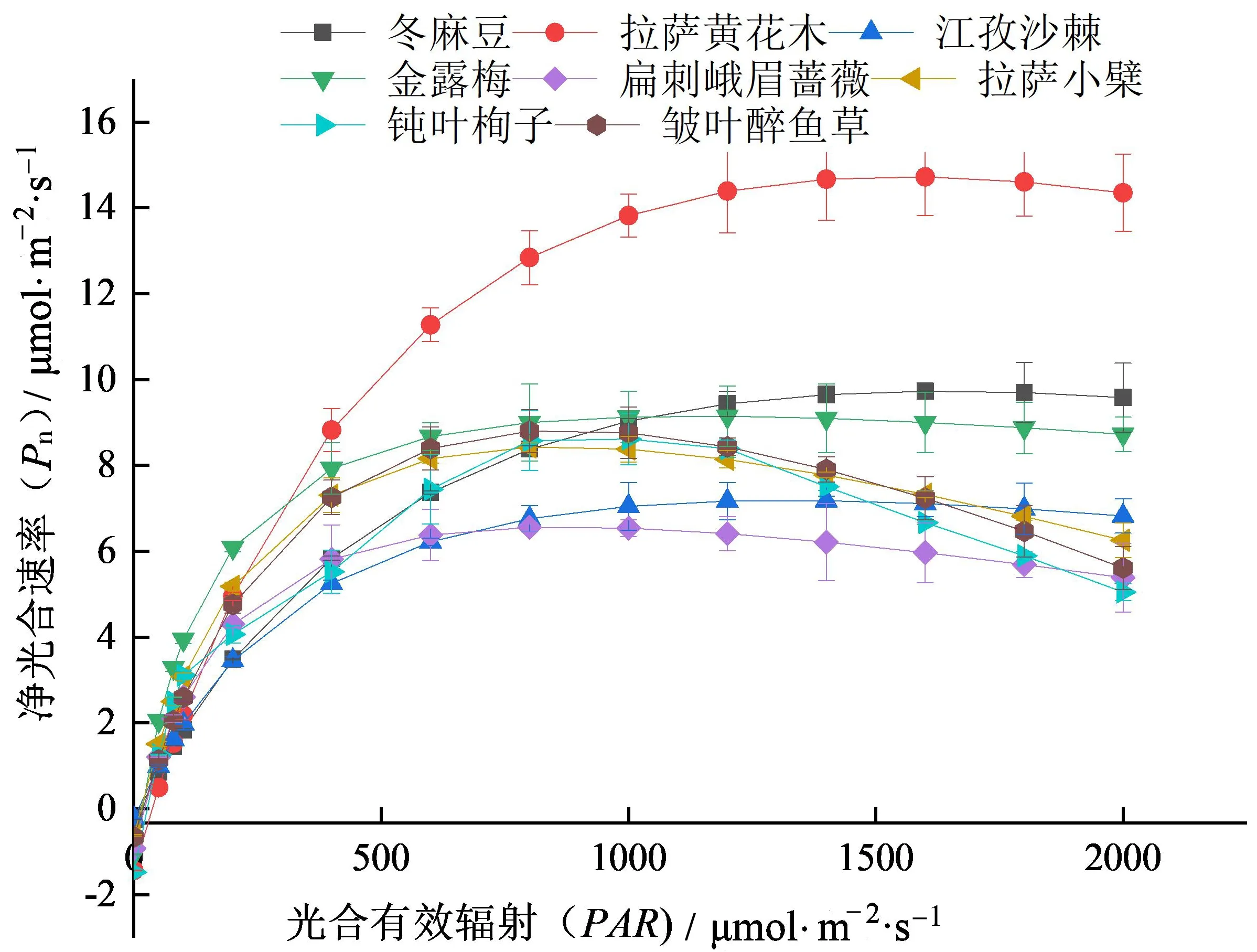
图1 8种灌木净光合速率-光合有效辐射曲线
利用叶子飘中直角双曲线修正模型拟合8种灌木Pn-PAR曲线,各参数见表3。由表中可知,8种灌木表观量子效率(α)、最大净光合速率 (Pnmax)、饱和光强(Isat)、光补偿点(Ic)、 暗呼吸速率(Rd)之间存在明显差异。8种灌木的α的变化范围为0.024~0.083,其中金露梅α最高(为0.083),冬麻豆(Salweeniawardi)最低(为0.024),说明金露梅在弱光下具有较强的光合能力,江孜沙棘、冬麻豆和钝叶栒子α之间比较接近,说明这三种植物利用弱光的能力相似;拉萨黄花木Pnmax最大,分别高于冬麻豆、江孜沙棘、金露梅、扁刺峨眉蔷薇(Rosaomeiensisf.pteracantha)、拉萨小檗、钝叶栒子、皱叶醉鱼草的51.31%、104.91%、60.92%、124.16%、74.45%、76.84%、66.80%;8种灌木Isat在800~1 700 μmol/(m2·s)之间,冬麻豆最大〔1 635.191μmol/(m2·s)〕,扁刺峨眉蔷薇最小〔872.164 μmol/(m2·s)〕,说明冬麻豆具有较强的向光性;Ic、Rd分别在8~18 μmol/(m2·s)和0.223~1.438 μmol/(m2·s)之间,其中拉萨黄花木的Ic、Rd最大,江孜沙棘最小,说明拉萨黄花木对光合产物的消耗较大,江孜沙棘对光合产物的消耗较小,有利于光合产物的积累。
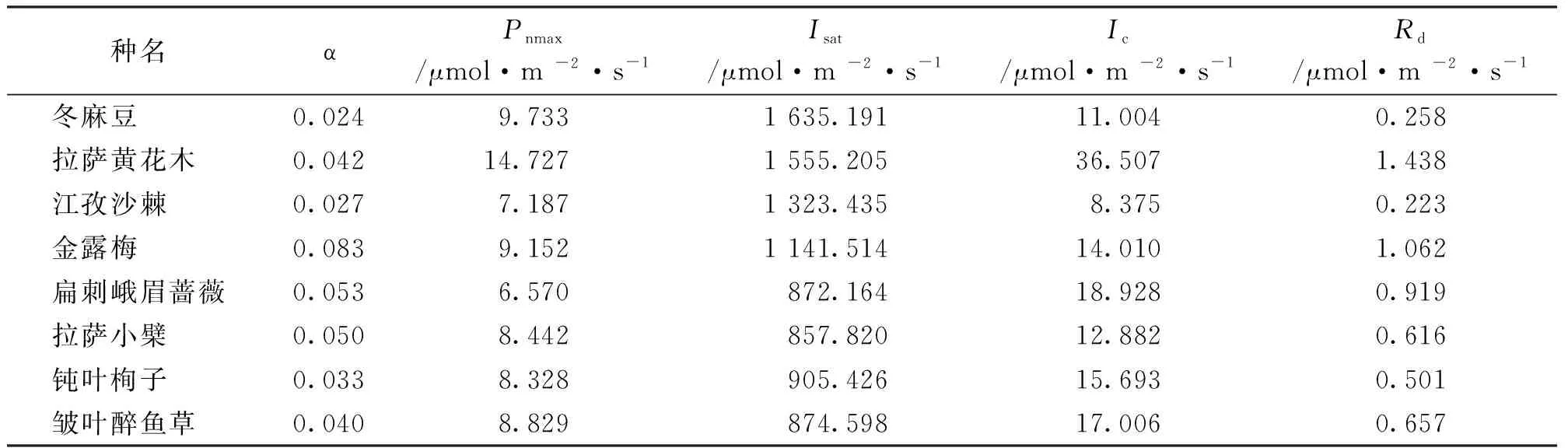
表3 8种灌木光合参数比较
2.3 电子传递速率-光合有效辐射曲线及参数比较
由8种灌木ETR-PAR曲线(图2)可知,8种灌木ETR-PAR曲线存在差异,变化趋势一致总体分为3个阶段:PAR在0~400 μmol/(m2·s)时,ETR随PAR的增加呈线性上升趋势;PAR在400 μmol/(m2·s)光饱和点时,ETR随PAR的增加上升幅度减缓;PAR到达光饱和点时,ETR最大,之后随PAR的增加ETR呈下降趋势,出现明显的光抑制现象。
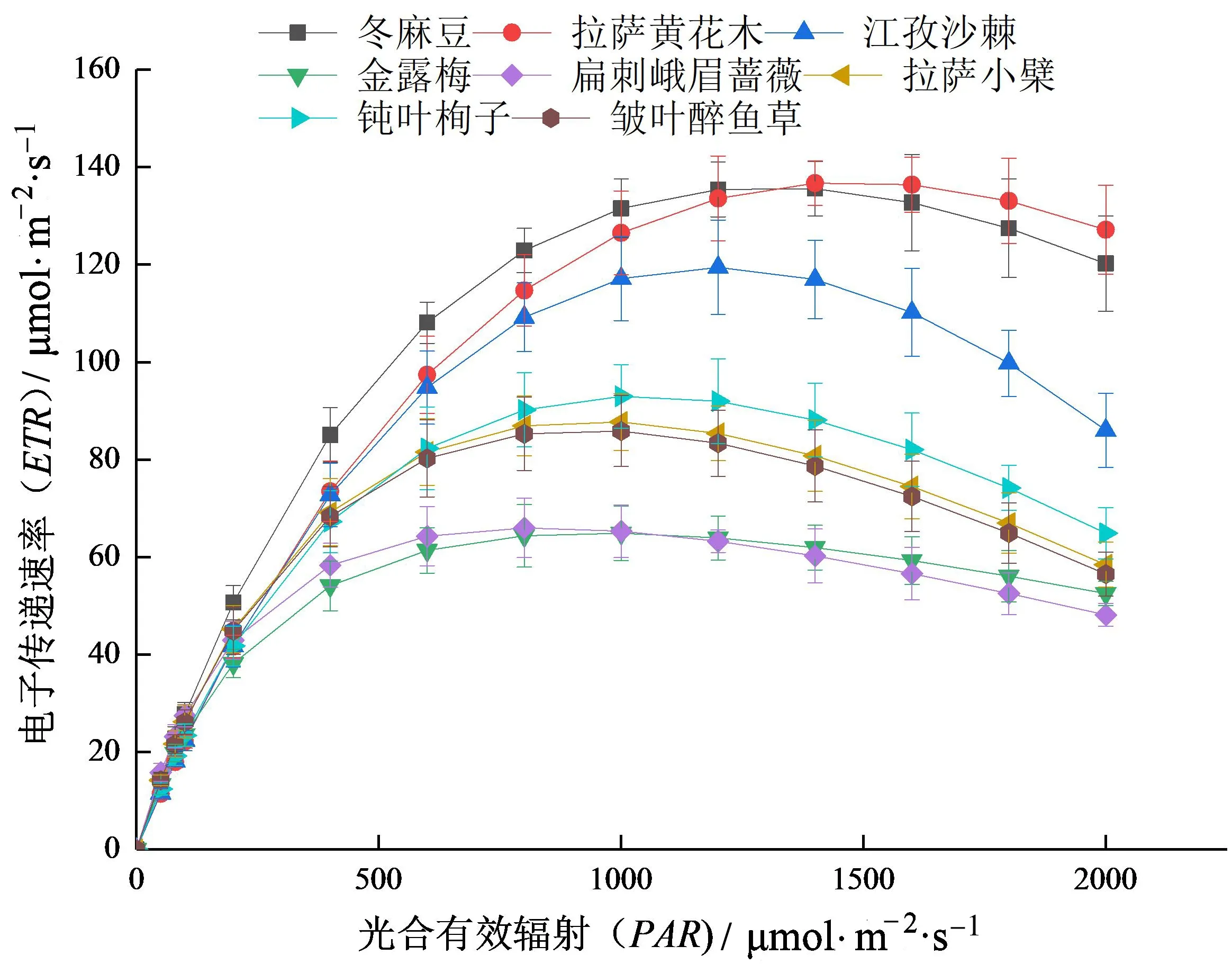
图2 8种灌木电子传递速率-光合有效辐射曲线
8种灌木叶绿素荧光参数见表4。由表4可知,8种灌木ETR-PAR曲线初始斜率(A)在0.230~0.380之间,其中扁刺峨眉蔷薇最大(为0.372),江孜沙棘最小(为0.236),表明扁刺峨眉蔷薇在西藏环境中捕获电子的能力较强;8种灌木最大电子传递速率(Jmax)之间差异较大,其中拉萨黄花木Jmax最大〔136.955 μmol/(m2·s)〕,分别是金露梅、扁刺峨眉蔷薇、拉萨小檗、钝叶栒子和皱叶醉鱼草的2.11倍、2.07倍、1.56倍、1.47倍、1.59倍;PARsat:拉萨黄花木>冬麻豆>江孜沙棘>钝叶栒子>金露梅>拉萨小檗>钝叶栒子>皱叶醉鱼草。
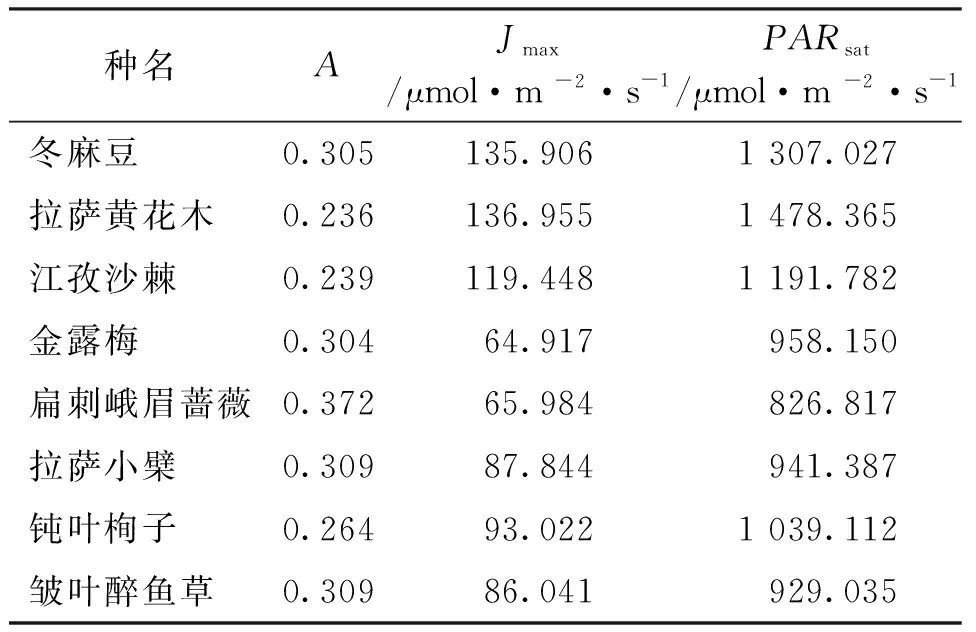
表4 8种灌木的叶绿素荧光参数比较
2.4 8种灌木光合指标比较
由表5可知,8种灌木各光合指标之间存在差异。拉萨黄花木与其余7种灌木的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)在5%水平上差异显著,江孜沙棘、扁刺峨眉蔷薇与冬麻豆、金露梅、拉萨小檗、钝叶栒子、邹叶醉鱼草的Pn之间存在显著性差异(P<0.05),扁刺峨眉蔷薇和江孜沙棘与冬麻豆、金露梅、拉萨小檗、钝叶栒子、邹叶醉鱼草的Tr、Gs在5% 水平上存在差异显著,金露梅与其余7种灌木的胞间CO2浓度(Ci)在5% 水平上差异显著。
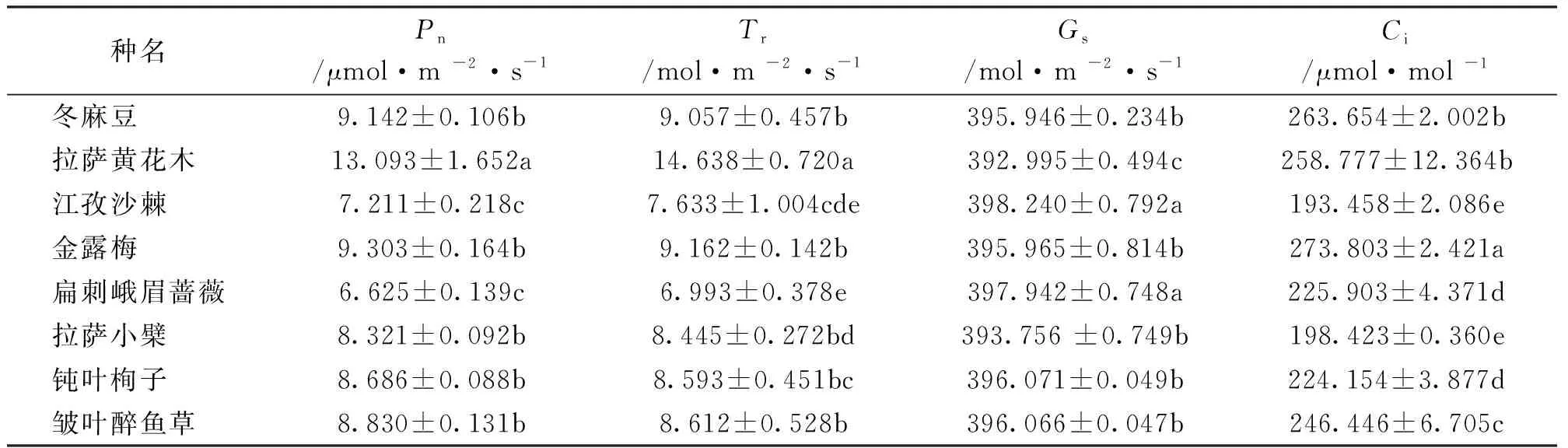
表5 8种灌木光合指标
8种灌木中,拉萨黄花木Pn、Tr值最高,分别为13.093 μmol/(m2·s)、14.638 mol/(m2·s),拉萨小檗Pn〔8.321μmol/(m2·s)〕是拉萨黄花木的63.55%,扁刺峨眉蔷薇Tr最小〔6.993 mol/(m2·s)〕;Gs比较结果依次为:江孜沙棘>扁刺峨眉蔷薇>钝叶栒子>邹叶醉鱼草>金露梅>冬麻豆>拉萨小檗>拉萨黄花木;金露梅Ci最大,江孜沙棘最小,前者是江孜沙棘的1.4倍。
2.5 8种灌木叶绿素荧光指标比较
由表6可以看出,冬麻豆、拉萨黄花木与其余6种植物的ETR之间存在显著差异(P<0.05),冬麻豆最高,扁刺峨眉蔷薇最小。8种灌木中Fo、Fm、Fv/Fm分别在79~251、326~1 091、0.70~0.78之间,其中冬麻豆的Fo、Fm显著高于其他灌木,说明冬麻豆荧光潜力较大,在光合反应中可吸收较多的光能,钝叶栒子最小;皱叶醉鱼草Fv/Fm最大,说明皱叶醉鱼草在本地对强光的适应性强。拉萨小檗、皱叶醉鱼草与其余6种植物的YpsⅡ之间存在显著差异(P<0.05),YpsⅡ变化范围在0.190~0.387之间,拉萨黄花木最高,扁刺峨眉蔷薇最低,说明8种灌木中,拉萨黄花木光合系统PSⅡ反应中心的实际光能转化效率最高,扁刺峨眉蔷薇最低。
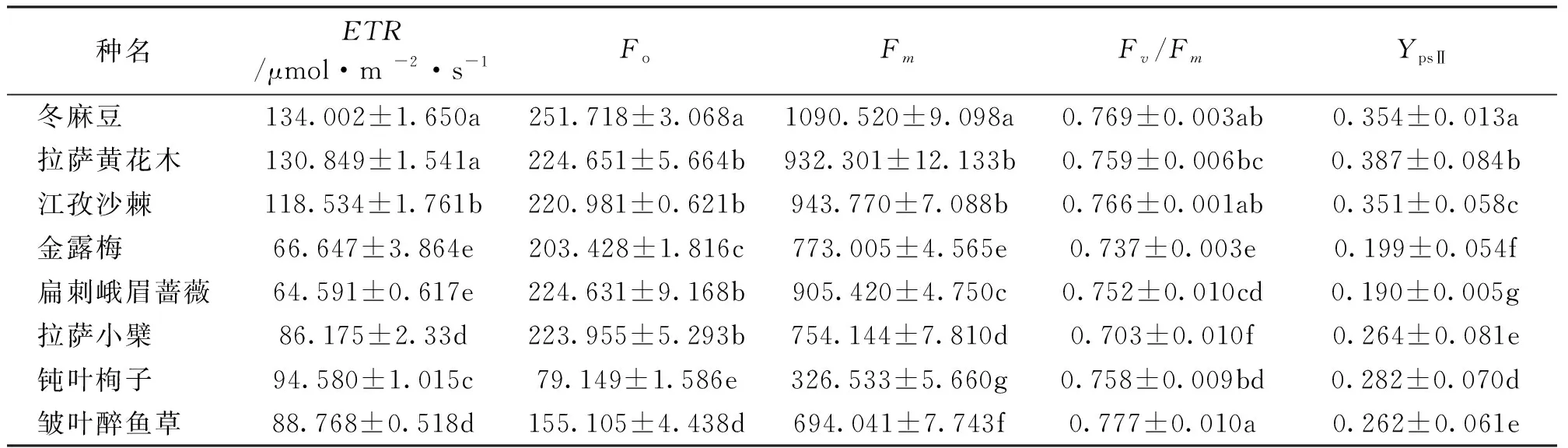
表6 8种灌木的叶绿素荧光指标比较
2.6 相关性分析
由表7可知,SPAD值与ETR之间呈极显著正相关;Pn与Tr、Ci之间存在极显著正相关,与Gs之间存在极显著负相关,与ETR、YpsⅡ之间存在显著正相关;Tr与Gs之间存在极显著负相关,与Ci、ETR之间存在显著正相关,说明Tr与Pn、ETR、Ci均有着密切关系,Tr是直接影响植物光合作用的内部因素;Gs与Ci之间存在显著负相关;ETR与Fm、Fv/Fm之间存在显著正相关,与YpsⅡ之间存在极显著正相关;Fo之间Fm存在极显著正相关。
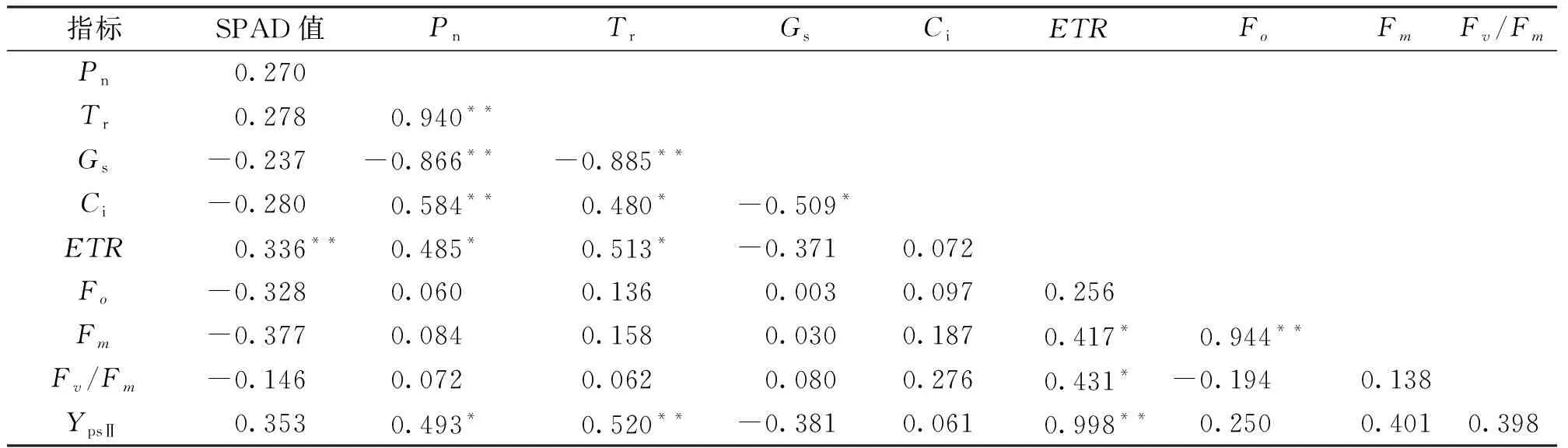
表7 相关性分析
3 讨论与结论
SPAD值可反映植物叶片的叶绿素含量,而叶绿素是植物叶片进行光合作用的物质基础[17-18]。8种灌木的苗高和SPAD值P值均小于0.05,说明8种灌木SPAD值之间差异显著,其中钝叶栒子SPAD值最大,拉萨黄花木次之,皱叶醉鱼草最小,钝叶栒子和拉萨黄花木叶片中叶绿素含量比较高,在光合作用中吸收的光能或光能转化比较好。
光响应和叶绿素荧光响应曲线是研究植物光合特性的方法之一,对于研究植物光反应过程及对环境的适应性具有重要意义[19],光响应宏观上体现了植物光合能力强弱,而叶绿素荧光响应从微观方面反映植物叶片叶绿素吸收光能和光系统Ⅱ (PSⅡ)的电子传递速率[20]。8种灌木Pn-PAR曲线和ETR-PAR曲线变化趋势基本一致,总体随PAR的增加呈现上升后下降趋势,当环境中光和辐射超过光饱和点时,植物会出现明显的光抑制现象。
植物生长环境中,Pnmax和Jmax决定了植物的最大光合能力,α反映了植物叶片对弱光的利用效率,Isat、Ic可直接反应植物在光环境的适应性,Rd、Lc反应植物对光合产物消耗及积累情况[21-22]。本研究中,8种灌木表观量子效率(α)、最大净光合速率 (Pnmax)、饱和光强(Isat)、光补偿点(Ic)、 暗呼吸速率(Rd)之间存在明显差异。金露梅α最大,江孜沙棘最小,说明金露梅在弱光环境下对光的利用效率较高;拉萨黄花木Pnmax最大,说明拉萨黄花木在强光下具有较大的光合能力,相比较其他7种灌木,强光环境中其产生光抑制现象滞后;冬麻豆Isat最大,拉萨黄花木次之,扁刺峨眉蔷薇最小,说明冬麻豆、拉萨黄花木对光的适应范围较广,具有较强的向光性;拉萨黄花木的Ic、Rd最大,江孜沙棘最小,说明拉萨黄花木对光合产物的消耗较大,江孜沙棘对光合产物的消耗较小,有利于光合产物的积累,抗性较强。拉萨黄花木Pnmax、Ic、Rd最大,说明拉萨黄花木属于高生产、高消耗的植物。
Pn是植物光合作用中光能利用的基础,其大小是植物光合能力的直接体现[23];Tr是反映植物水分调节能力、耗水潜力和适应性能力的重要生理指标之一[24-25];气孔是植物光合的门户,Gs是反映植物叶片气孔的重要参数,可调节植物叶片的蒸腾作用[26-27]。本研究中,拉萨黄花木Pn、Tr值最高,因为植物进行光合作用时需要水分,并通过水分的运输供应植物所需的营养成分,说明拉萨黄花木在强光环境条件下的自身调节能力较强;江孜沙棘Gs最高,拉萨黄花木Gs的最小;金露梅Ci最大,江孜沙棘最小。
植物的光合作用过程中,在外界环境条件相同的情况下,ETR和YpsⅡ值越高,说明植物光合作用碳同化过程中形成的电子传递载体就越多[28-29]。本研究中,冬麻豆、拉萨黄花木与其余6种植物的ETR之间存在显著差异(P<0.05),拉萨小檗、皱叶醉鱼草与其余6种植物的YpsⅡ之间存在显著差异(P<0.05);冬麻豆ETR最高,拉萨黄花木次之,扁刺峨眉蔷薇最小;拉萨黄花木YpsⅡ最高,冬麻豆次之,扁刺峨眉蔷薇最低;说明,拉萨黄花木和冬麻豆电PSⅡ电子传递活性高于其余6种植物,光合作用碳同化的能力和光能实际转化效率较强,而扁刺峨眉蔷薇较低。扁刺峨眉蔷薇A最大,说明扁刺峨眉蔷薇光合作用PSⅡ启动效率较快,捕获电子的能力较强;拉萨黄花木Jmax和PARsat最大,皱叶醉鱼草最小,说明在西藏立地环境中,拉萨黄花木叶片的PSⅡ电子传递活性和光能利用率较强,皱叶醉鱼草叶片光合作用受到西藏环境的影响,导致光合效率较低。植物在没有受到胁迫情况下,Fv/Fm在0.80~0.85之间[30],本研究中,8种灌木Fv/Fm的变化范围为0.70~0.78之间,说明8种灌木均受到环境的胁迫;冬麻豆的Fo、Fm显著高于其他灌木,说明冬麻豆荧光潜力较大,在PSⅡ光合反应中心中可吸收较多的光能,但是易受到环境的胁迫[31]。
综上所述,拉萨黄花木和冬麻豆在西藏强光环境中自身调节能力较强,光合能力较大,对光的适应范围较广,是明显的阳性树种, 适合种植在全光环境中。在西藏造林树种选择时,如果只考虑光照因素的情况下,拉萨黄花木应该为优先树种。此次研究仅考虑环境中的光照因素,是一个单因素情况下林木生长阶段的光合特性比较,在综合环境因素及整个生长过程中,光合特性存在的差异,还需深入研究。
