毛竹磷转运蛋白Ⅰ家族基因鉴定及表达模式
2022-06-28王绍良张雯宇高志民周明兵杨克彬宋新章
王绍良,张雯宇,高志民,周明兵,杨克彬,宋新章
(1. 浙江农林大学 省部共建亚热带森林培育国家重点实验室,浙江 杭州 311300;2. 国际竹藤中心 竹藤资源基因科学与基因产业化研究所 国家林业和草原局竹藤科学与技术重点开放实验室,北京 100102)
磷(phosphorus,P)是植物中脂质、核酸、二磷酸腺苷(ADP)、三磷酸腺苷(ATP)和糖类的重要组成元素,是含量仅次于氮的第二大限制性养分[1],在植物生长发育过程中发挥着不可替代的作用。植物吸收磷素主要以可溶性的磷酸根离子形式为主[2],虽然土壤中磷素含量很高,但是可供利用的无机磷素含量较少,是导致植物缺磷的主要原因之一[3]。植物磷转运蛋白(phosphate transporter,PHT)是植物从土壤中吸收无机磷酸盐并在体内进行再分配利用的载体。磷吸收动力学研究表明:植物在进化过程中形成了2类不同的磷吸收转运系统[4],分别为高亲和力磷吸收转运系统(highaffinity P uptake system)和低亲和力磷吸收转运系统 (low-affinity P uptake system)[5]。研究者最早在拟南芥Arabidopsis thaliana中克隆出了磷转运蛋白基因AtPHT1.1,随后通过基因组测序、同源序列分析等方法陆续在大豆Glycine max[6]、玉米Zea mays[7]、大麦Triticum aestivum[8]和番茄Lycopersicon esculentum[9]等植物中克隆出了磷转运蛋白基因。植物磷酸盐转运蛋白属于MFS超级家族(major facilitator superfamily)[10],有12个疏水的跨膜区域,蛋白质结构高度相似,在氨基酸序列中存在保守特征序列GGDYPLSATIMSE,以及保守的磷酸化位点和糖基化位点。磷转运蛋白分为4个亚家族,分别为PHTⅠ、PHTⅡ、PHTⅢ、PHTⅣ[11]。拟南芥中,AtPHT1.1、AtPHT1.2和AtPHT1.3对拟南芥吸收磷的贡献非常大,而且AtPHT1.1在长距离运转磷的过程中具有重要作用[12]。研究表明:高磷环境中OsPHT1是水稻Oryza sativa吸收和转运磷素的关键PHTⅠ成员[13]。缺磷显著诱导OsPHT2、OsPHT4、OsPHT8、OsPHT9 和OsPHT10等的表达。OsPHT8几乎在水稻的各个器官中都强烈表达,是组成型磷转运子;OsPHT8下调会使新叶磷含量下降,老叶磷含量上升,处于灌浆期的胚乳和胚中的磷含量显著下降,说明水稻OsPHT8对磷素从源到库的再分配起到至关重要的作用[14]。大豆中,GmPHT7在根际成熟丛枝菌Arbuscular mycorrhiza(AM)根的根冠小柱细胞、皮层细胞和无菌根的侧根原基细胞中表达,在衰老叶片的维管束末端少数管胞中也有表达,主要负责向种子转运再活化的磷素[15]。此外,GmPHT10和GmPHT11也会受AM诱导表达[16],即磷酸盐转运蛋白基因广泛存在于各种植物中。
毛竹Phyllostachys edulis属于禾本科Gramineae刚竹属Phyllostachys,具有经济价值高、用途广泛、栽培面积大等特点[17]。影响毛竹生长发育的因素有很多,如林地养分、水分不足[18],粗放的抚育管理和病虫害防治不及时等[19],其中,磷元素是限制毛竹林生长的重要营养元素之一。因此,研究毛竹磷素转运和吸收的相关基因的表达具有重要意义[20]。由于土壤对磷素的化学固定作用,磷素利用率普遍较低[21]。磷酸盐转运蛋白是植物吸收和转运磷素的重要参与者。本研究利用生物信息学方法鉴定毛竹PHTⅠ家族成员,分析其基因启动子、蛋白质理化性质、基因结构、氨基酸保守基序、基因在染色体的位置以及基因的组织表达特异性等,以期为深入研究毛竹PHT基因功能,探索毛竹磷素利用机制提供参考。
1 材料与方法
1.1 毛竹 PHTⅠ家族序列的获取和确定
拟南芥和水稻PHT基因的CDS序列、基因序列以及氨基酸序列分别下载于Tair (https://www.arabidopsis.org)和 Rice Genome Annotation Project (http://rice.plantbiology.mus.edu)。在毛竹基因组数据库Bamboo GDB (http://bamboo.bamboogdb.org/)进行 BlastP、BlastN (e-value=1e-10) 比对,获取毛竹 PHTⅠ同源基因的候选序列。通过SMART数据库(http://smart.embl-heidelberg.de)和PFAM数据库(http://pfam.xfam.org)确定获取的候选序列的保守结构域的准确性和完整性,保留具有编码完整保守结构域的候选序列,并进行基因命名(PePHTs)。
1.2 毛竹 PHTⅠ的生物信息学分析
利用ProtParam (http://web.expasy.org/prot-param/)获取毛竹PHTⅠ基因编码蛋白质的基本理化特性,使用 Gene Structure Display Server 2.0 (http://gsds.cbi.pku.edu.cn/)在线工具分析 PePHTs 的基因结构,通过MEME Version 5.3.2 (http://meme-suite.org/tools/meme)获取毛竹PHTⅠ的保守基序,利用 PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线分析平台对PePHTs所含作用元件进行分析,在Excel软件中整理Tab结果文件, 用TBtools软件的Basic BioSequence View工具展示顺式作用元件的分布[22],使用Plant-mPLoc算法(http://www.csbio.sjtu.edu.cn)预测毛竹PHTⅠ的亚细胞位置。
1.3 毛竹 PHT Ⅰ基因系统进化分析
使用Clustal X 1.8软件对毛竹、拟南芥、水稻等PHT基因的CDS序列进行多重比对,比对结果在MEGA 7.0软件中使用邻接法(neighbor-joining)构建系统进化树,bootstrap试验重复1 000次[23],其他参数设置为默认值。利用在线工具Evolview (https://www.evolgenius.info/evolview/)进行树图编辑[24]。
1.4 毛竹PHTⅠ基因的染色体分布及适应性进化分析
利用在线软件 MEME Version 5.3.2 (http://meme-suite.org/tools/meme)对毛竹 PHTⅠ的 motif进行预测和分析,motif数量设置为 10。使用 TBtools软件的 Gene Location Visualize from GTF/GFF 工具展示基因在染色体上的位置。使用KaKs_Calculator软件计算毛竹PHTⅠ基因的CDS序列的选择压力值(ω)[25]。
1.5 毛竹 PHTⅠ基因组织表达特异性分析
根据毛竹和竹笋不同 组织的转录组数据[26],用 PePHTs 的 RPKM (reads per kilo-bases per million reads)值表示基因的表达丰度,利用TBtools制作热图展示基因的表达丰度。
2 结果与分析
2.1 毛竹PHTⅠ家族基因鉴定及基本理化特征
通过比对分析毛竹基因组,鉴定并确定编码完整MFs_1的候选基因共20个。植物PHT一般都含有MFs_1保守结构域,候选基因编码的蛋白质都含有保守的跨膜结构域,与MFS超家族的PHT家族特征相同。根据PHT候选基因在毛竹Scaffold中的位置以及同源基因的名称依次命名为:PePHT1~PePHT20。
对20个PHTⅠ家族基因进行生物信息学分析,其理化性质结果(表1)显示:PePHTs编码的氨基酸序列长度,最长为696 个氨基酸 (PePHT1),最短为432 个氨基酸(PePHT17),理论等电点为6.84~9.30,除PePHT15外,其他均为碱性蛋白质,分子量为48.61~76.37 kDa。疏水性测试显示:所有蛋白质疏水性值(grand average hydropathicity,GRAVY)>0,说明该家族蛋白质均为疏水性蛋白质。脂肪族氨基酸指数显示:PHTⅠ家族的蛋白质热稳定性为 83.79~105.07,热稳定性差异较大。亚细胞定位结果显示:PePHTs均定位于细胞膜中。
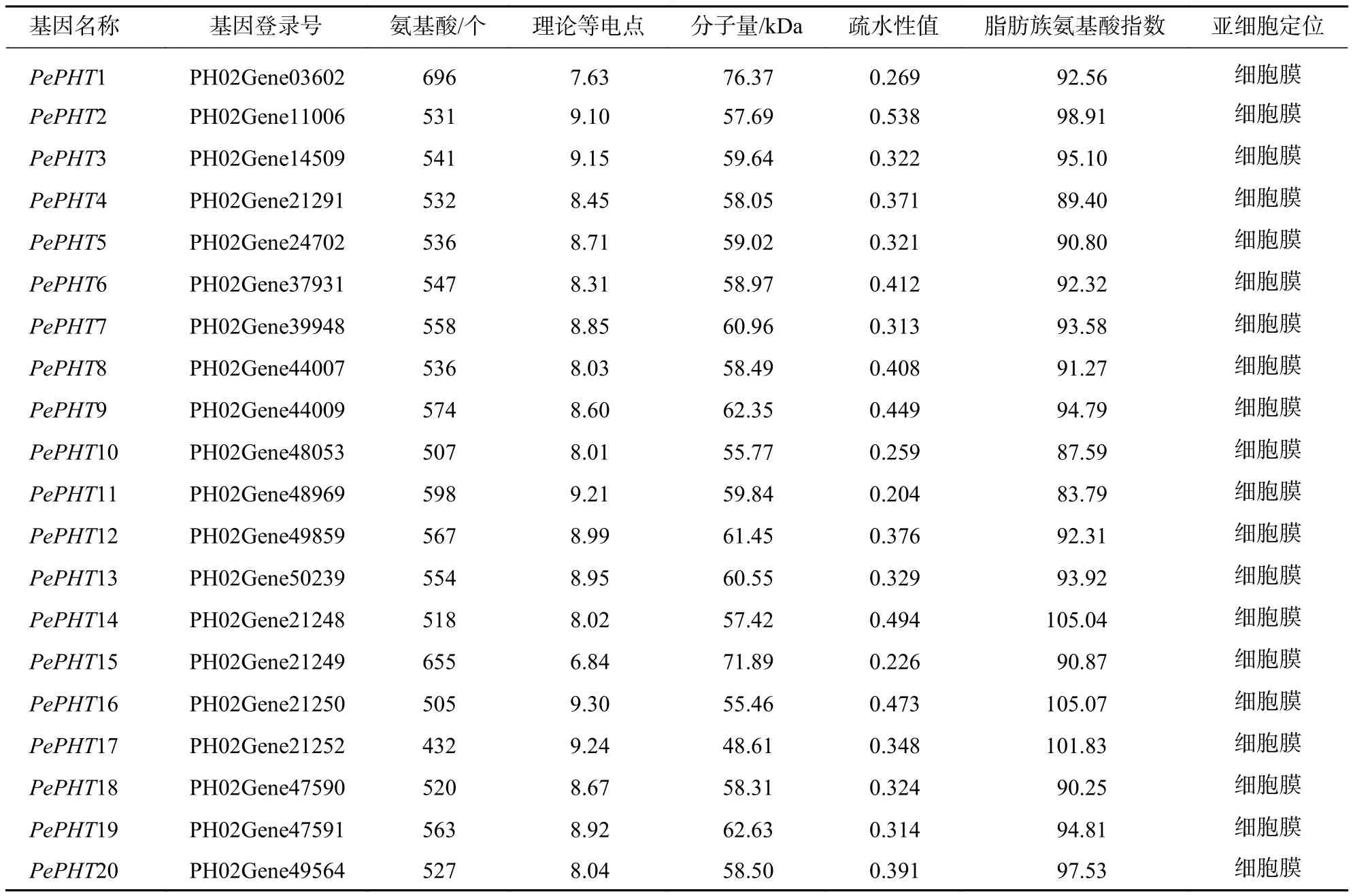
表1 毛竹 PHTⅠ家族基因编码蛋白序列的理化性质Table 1 Physicochemical properties of proteins encoded by PHTⅠ gene family in Ph. edulis
2.2 毛竹PHTⅠ家族基因结构及保守基序特征分析
由图1可知:毛竹PHTⅠ家族基因多数含1~2个内含子(intron),在所有PePHTs中,PePHT14内含子区域最长,PePHT8和PePHT11内含子区域最短。保守基序分析显示:PePHTs含有7~10个保守基序,分别命名为motif1~motif10。其中有6个基序高度保守,分别是motif2、motif3、motif4、motif5、motif6、motif8,其他基序在部分序列中缺失。9个PePHTs中含有10个motif,其他11个PePHTs缺失1~3个motif,PePHT1、PePHT10都缺少motif7,PePHT11缺少motif1,PePHT12缺少motif9、motif10,PePHT14、PePHT15、PePHT16、PePHT18、PePHT19、PePHT20都缺少motif10,PePHT17则缺少motif7、motif9、motif10。PePHTs高度保守基序的氨基酸数目也不尽相同,最长的motif由50个氨基酸组成,最短的motif由29个氨基酸组成。

图1 毛竹 PHTⅠ家族基因的结构Figure 1 Structures analysis of PHTⅠ gene family in Ph. edulis
为探究PePHTs受内在调控因子调节的情况和对外界环境的响应,选取PePHTs距离起始密码子上游2 000 bp的序列,对其所含顺式作用元件和应答元件进行分析。如图2所示:所有启动子的顺式作用元件种类比较相似,包括MYB转录因子结合的顺式作用元件,生长素、赤霉素、水杨酸、脱落酸等激素响应元件以及低温、干旱、缺氧、光等非生物胁迫响应元件。由此表明:PePHTs的转录表达可能会受到非生物胁迫和激素的影响。
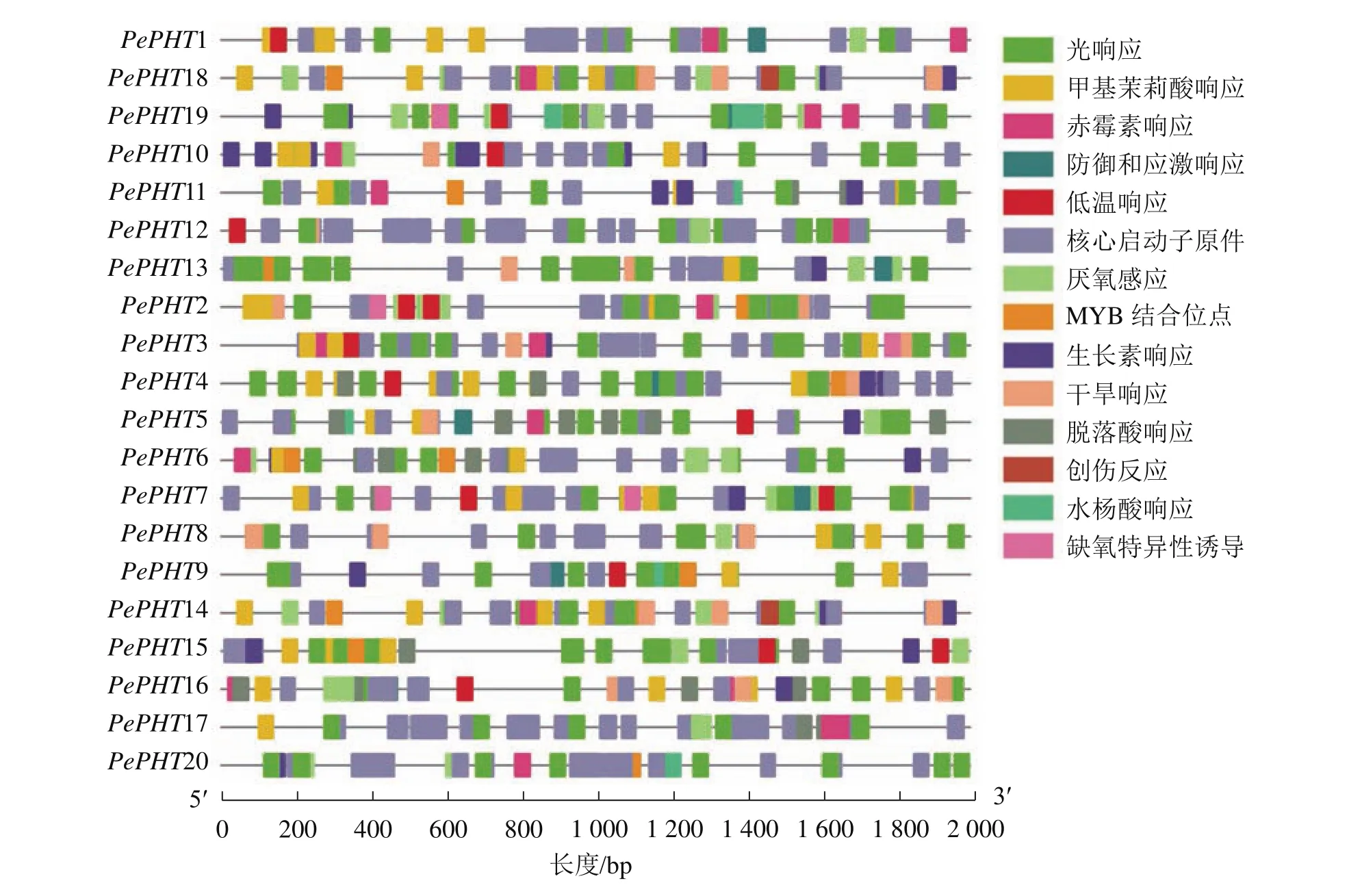
图2 毛竹PHTⅠ家族基因启动子区顺式作用元件位置信息Figure 2 Location information of cis-acting regulatory elements identified in the promoter region of PHTⅠ gene family in Ph. edulis
2.3 毛竹 PHTⅠ家族基因系统进化分析
为了解PePHTs的进化关系,预测基因潜在功能,本研究提取20个毛竹、18个拟南芥和25个水稻的PHT基因的CDS序列进行多重序列比对,并利用MEGA 7.0软件根据邻接法构建系统进化树。由图3显示:毛竹、拟南芥和水稻的PHT基因被聚类到4个亚家族中,来自毛竹的20个PePHTs均分布在第Ⅰ亚家族的5个分支上,1个分支中只有PePHTs基因,另4个分支和水稻聚类在一起。此外,第Ⅰ亚家族中还包含9个拟南芥和12个水稻的PHT基因。PHT亚家族中的基因在功能上存在一定差异,如大多数第Ⅰ亚家族成员主要在直接接触根际环境的根毛和表皮细胞中表达,参与根系对环境中磷元素的吸收过程[27-28],定位在细胞膜上。相比拟南芥,毛竹PHT基因均优先与水稻PHT基因聚类,推测PePHTs在功能上可能与水稻同源基因更为相似。
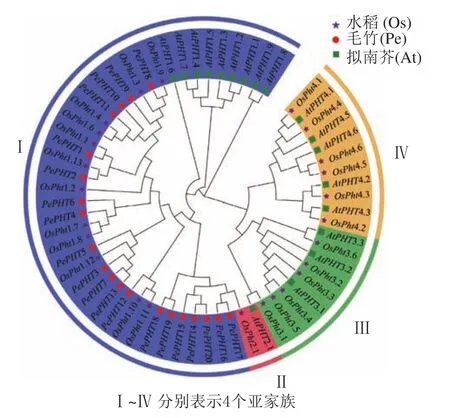
图3 毛竹、拟南芥和水稻 PHT 家族基因系统进化树Figure 3 Phylogentic tree of PHT gene family from Ph. edulis, A.thaliana and O. sativa
2.4 毛竹PHTⅠ家族基因在染色体上的位置及适应性进化分析
染色体定位显示:20个毛竹PHT基因位于10条染色体上,其中23号染色体上最多,有6个,分别是PePHT14、PePHT15、PePHT16、PePHT17、PePHT18、PePHT19;其次是24号染色体,有3个,分别是PePHT10、PePHT11、PePHT20,推测这2个基因簇中的基因可能分别编码催化2种新陈代谢途径中不同步骤的磷转运酶[29]。5号、14号以及15号染色体上各有2个基因,其余染色体各有1个(图4)。除数量分布不均匀外,各基因在染色体上的分布位置也不均匀,大多数基因位于染色体的中部,少量则位于顶部和底部。适应性分析结果表明:PePHT7的ω>0,其他基因的ω<0,表明PePHT7受到正选择压力,其他基因受到负选择压力[30]。
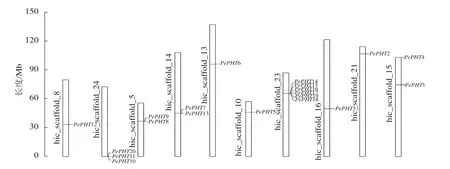
图4 毛竹 PHTⅠ基因在染色体上的位置Figure 4 Chromosomal location of PHTⅠ genes from Ph. edulis
2.5 毛竹 PHTⅠ家族基因组织表达模式分析
根据毛竹6个不同部位的转录组表达谱数据[26],对毛竹PHTⅠ家族基因进行组织特异性表达分析。由图5可知:毛竹不同组织中的PePHTs表达丰度差异较大,其中PePHT5、PePHT7、PePHT8、PePHT12、PePHT14、PePHT19在箨鞘中大量表达;PePHT2、PePHT3、PePHT6、PePHT13、PePHT15在根中表达丰度较高;PePHT18在叶中大量表达,PePHT10、PePHT20在叶鞘中大量表达,PePHT16、PePHT17在叶和叶鞘中也有少量表达。
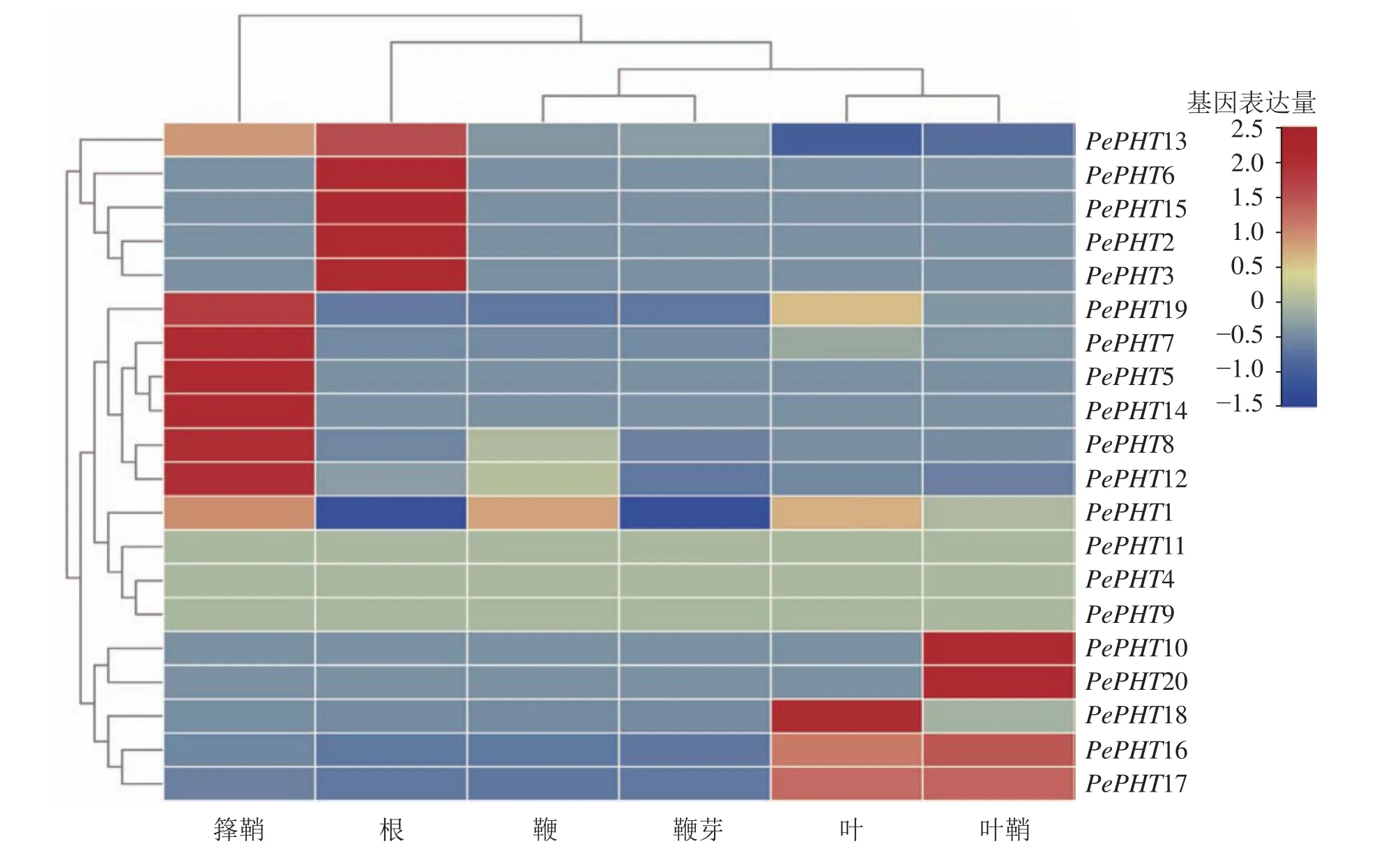
图5 PePHTs在毛竹 6 个部位的表达Figure 5 Expression analysis of PePHTs in 6 parts of Ph. edulis
3 讨论
植物从环境中吸收磷元素的过程中,磷酸盐转运蛋白起着至关重要的载体作用。植物PHTⅠ磷酸盐转运蛋白为膜蛋白,大多数属于高亲和力转运系统,并且具有相似的蛋白质序列和化学结构。在双子叶植物拟南芥和单子叶植物水稻中分别发现了9个[31-32]和12个PHTⅠ家族成员[6]。VERSAW等[33]证实拟南芥AtPHT2.1除了参与植物对磷元素的吸收与转运,还可能参与茎部对磷的转运,其蛋白质定位于叶绿体内膜上。PHTⅡ磷酸盐转运蛋白也在拟南芥[34]、马铃薯Solanum tuberosum[35]、茄Solanum melongena[36]、菠菜 Spinacia oleracea[37]、烟草 Nicotiana tabacum[38]和小麦 Triticum aestivum[39]等多种植物中被发现。PHTⅢ磷酸盐转运蛋白最初在拟南芥中被克隆得到的。研究表明:PHTⅢ 磷酸盐转运家族与 PHTⅠ家族一样,通过 P/H+同向转运和P/OH-反向转运方式参与细胞质间磷的交换[39-40]。随后水稻、玉米和大豆等其他植物也克隆得到了该蛋白[41]。但是PHTⅣ 磷酸盐转运蛋白只在少数几种植物中发现[42],研究报道很少。
植物磷转运蛋白是众多转运蛋白中的一类重要蛋白家族,它们在植物的根、茎、叶、花等器官都有分布,是磷元素吸收和运转的主要载体[1]。物种中均存在PHT基因,说明该基因具有重复性和多样性,这也是基因组重新排列和扩展的结果。本研究从毛竹基因组中鉴定出20个PHTⅠ家族基因,每个成员都含有Sugar_tr和MFs_1保守结构域,这是它们具有相似功能的基础。理化性质分析显示:PHT长度、理论等电点以及分子量区间跨度较大,这有可能是因为基因进行多次复制转录后进化的结果。本研究预测了毛竹PHTⅠ基因在细胞中的位置,发现毛竹PHTⅠ基因都分布在细胞膜上,与已有研究一致[3],说明鉴定出的毛竹PHTⅠ亚家族中的基因符合磷酸盐转运蛋白基因的特性。在植物进化过程中,选择压力能很好地体现发挥重要功能的蛋白的变化。基因适应性进化分析显示:大多数PePHTs基因受到较强烈的负选择压力,说明毛竹磷转运蛋白相对趋于稳定,是其保持原有重要功能的原因;同时PePHT7基因受到正选择压力,提示该基因编码的蛋白可能会延伸出一些新的功能[30]。植物基因组织特异性表达与基因的功能关系密切,毛竹磷转运蛋白在箨鞘、根、叶和叶鞘都有1个及以上的基因大量表达,PePHT4、PePHT9、PePHT11等3个基因在任何组织中都没有检测到,还需深入研究;PePHT1在根和鞭芽中明显下调,说明基因在不同组织中表达丰度不一样且发挥着不同的作用。
系统进化分析发现:毛竹PHTⅠ基因聚类在第Ⅰ亚家族的5个分支上,同一支中的基因可能具有相似的功能。拟南芥的PHTⅠ亚家族中有9个成员,其中AtPHT1.6在花粉中表达,其余8个在根中表达[43];当拟南芥缺磷时,AtPHT1.1和AtPHT1.4会大量表达,这2个磷酸盐转运蛋白提供了70%的磷酸盐转运活性[44]。水稻PHTⅠ亚家族中有12个成员[6],其中OsPHT1主要功能是吸收磷素和体内磷酸盐的再分配[13],OsPHT2主要在地上部分表达,也是水稻PHTⅠ亚家族中唯一一个低亲和力磷转运蛋白[45]。PHTⅠ家族在水稻和拟南芥吸收和转运磷的过程中具有重要作用[32,45],推测在毛竹对磷的吸收转运过程中也可能具有重要作用,但需要进一步的转录组数据进行验证。
