鲜食葡萄果实质地遗传分析
2022-06-24马丽郭修武乔军孙凌俊
马丽 郭修武 乔军 孙凌俊





摘要:探索分析葡萄果实质地的遗传趋势,为葡萄育种过程中的亲本选择选配以及杂交后代果实质地预测提供理论依据。本试验以脆肉型品种红地球为母本,软肉品种玫瑰香、金星无核为父本配置杂交组合,测定和分析葡萄亲本及其杂交后代果实果皮硬度(PPH)、果皮破裂距离(PB)、果肉硬度(SF),并进行遗传分析。结果表明,2个杂交组合后代葡萄果实果皮硬度、果皮破裂距离、果肉硬度均表现出连续变异,属于多基因控制的数量性状,其中果皮硬度和果肉硬度子代均值小于亲中值,呈现向小回归的趋势,加性效应解体,但由于新的累加效应,后代也出现超高亲株系。葡萄果实质地是属于多基因控制的数量性状,广义遗传力较高,主要受遗传因素影响。
关键词:葡萄;果实质地;果皮硬度;果皮破裂距离;果肉硬度;遗传分析
中图分类号:S663.103 文献标志码:A
文章编号:1002-1302(2022)11-0136-06
收稿日期:2021-08-09
基金项目:国家现代农业产业技术体系建设专项(编号:CARS-29-6);农业攻关及产业化项目(编号:2020JH2/10200019);农业种质资源发掘、创制与利用专项(编号:2021GR2904)。
作者简介:马 丽(1980—),女,辽宁铁岭人,博士,副研究员,主要从事葡萄资源及育种工作。E-mail:43845590@qq.com。
通信作者:孙凌俊,硕士,副研究员,主要从事葡萄栽培及育种工作。E-mail:zglnykslj@163.com。
葡萄属于葡萄科(Vitaceae)葡萄属(Vitis)多年生植物,是世界上最古老的物种之一。葡萄在世界果树生产中占有非常重要的位置,据联合国粮食及农业组织(FAO)统计,2018年全世界葡萄栽培面积约715.8万hm,总产量约为7 912.6万t。我国葡萄栽培面积达到79.8万hm,约占全球总面积的11.1%,产量1 349.5万t,约占全球葡萄总产量的17.1%,是世界葡萄的第一生产大国。就全世界葡萄生产而言,近70%的葡萄用于加工,超30%用于鲜食,而我国则以鲜食为主,占据70%以上,鲜食葡萄生产是我国葡萄产业的主体。
葡萄属于典型的非呼吸跃变型果实,果实软化发生时间比呼吸跃变型果实要早,所以就其鲜食品质而言,果实质地已经成为判断鲜食葡萄果实新鲜程度的一个重要指标。然而葡萄果实质地性状是包括若干数量性状的综合性状,双亲基因型及其环境都可以对后代表现型产生不同程度的影响。了解葡萄果实质地的遗传规律,将有助于指导育种过程中亲本的选择选配,获得具有理想果实质地的葡萄新品种。目前有关葡萄果实质地遗传研究的报道还比较少,且以往研究中果实质地多以感官评价为主,没有明确的量化指标和界限标准,主观性较强,或杂交群体较小。本试验以前期构建的2个杂交群体红地球×玫瑰香、红地球×金星无核为研究对象,利用质构仪对群体果实质地进行整果穿刺法测定,并进行遗传分析,探讨不同群体果实质地的遗传趋势,为葡萄常规杂交育种及今后果实质地QTL定位奠定重要的理论基础和数据支撑。
1 材料与方法
1.1 材料
本试验全部试材均采自沈阳农业大学葡萄试验园(41°50′24″N,123°24′41″E),共包括3个亲本品种和2个杂交群体,亲本为典型的脆肉型品种红地球(Vitis vinifera L.)和软肉型品种玫瑰香(V. vinifera L.)、金星无核(V. vinifera×V. labrusca.),2个群体后代植株均采用贝达嫁接,每个后代定植3株嫁接苗。篱架、避雨栽培,常规管理,2018—2019年分别从杂交后代中随机选取部分结果后代及对应亲本进行果实质地各项指标测定。
样品采集分别于每年7—10月进行,鉴于群体后代果实成熟期差异较大,从果实转色开始,每5 d监测果实可溶性固形物含量,并结合果实发育状态、种子褐化程度等确定果实成熟及采样时期(种子呈深褐色,可溶性固形物含量一般达到18%),以确保测定的是成熟期果实质地。采样时间一般确定在上午07:00—09:00时,每个后代随机采集3穗果实,果实放在标记好的自封袋内后迅速带回实验室,将3穗果实剪成留有短果蒂的果粒,再从中随机选取30粒较为一致的果实进行果实质地检测,同一批样品尽量在1 d内检测完成。
1.2 方法
1.2.1 果实质地相关指标测定
葡萄果实质地测定采用的是整果穿刺测试法。在进行穿刺试验前需对果实进行筛选,剔除病果、烂果,畸形果,并尽量使果實成熟度、大小等基本保持一致。每个品种随机选取30个果粒进行测定,为了减少试验误差,所有的测量均由1人完成。穿刺试验使用的是英国Stable Micro Systems 公司生产的TA. XT plus质构仪,穿刺试验选择的是2 mm直径的P/2针状探头,测试在室温下进行,测试时将被测试的浆果放置在质构仪载物台的中心位置上,用手稍稍固定,力量以不压迫果实为宜,样品被穿刺部位为葡萄浆果中部偏上区域。质构仪主要参数设定:测前速度为1 mm/s,贯穿速度为1 mm/s,测后速度为10 mm/s,穿刺深度设为7 mm,负载触发力5 g。参数设定时需要注意,穿刺既要保证一定的穿刺深度,又要避免在穿刺过程中探头接触到葡萄种子,影响测试结果。质地相关指标包括:果皮硬度(PPH),果皮破裂距离(PB),果肉硬度(SF)。果皮硬度(PPH)是从探头接触果皮开始到探头穿刺果皮瞬间所用的力(g);果皮破裂距离(PB)是从接触到刺破果皮瞬间探头下行位移(mm);果肉硬度(SF)是探头穿刺果肉到测试结束所测感应力的平均值(g)。
1.2.2 数据处理
利用Excel对试验数据进行收集与整理,统计群体在不同年份平均值、亲中值、变异系数、超高亲率、超低亲率等。使用SPSS 19.0软件进行果实表型数据的正态性分布检验及各年份间Spearman相关性分析。
2 结果与分析
2.1 2个杂交群体果皮硬度遗传分析
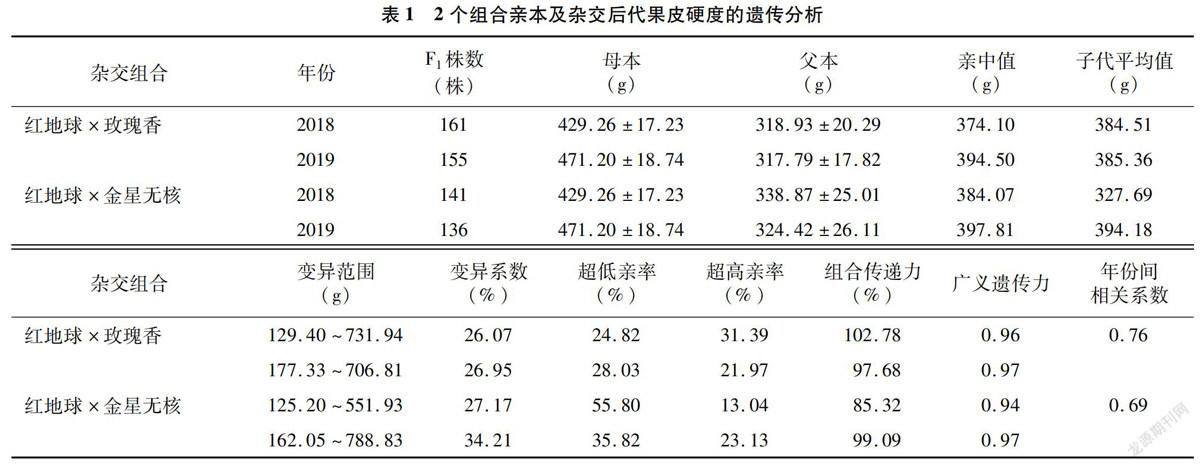
果皮硬度是果实质地的一个非常重要的指标。分别对2个杂交组合群体果皮硬度进行各年份间Spearman相关性检验(P<0.001),结果显示,红地球×玫瑰香组合2018年和2019年果皮硬度呈显著正相关,相关性系数为0.76,而红地球×金星无核2年的相关性系数为0.69(表1)。本试验对各组合表型数据进行了基于Shapiro-Wilk test的正态分布检验(P=0.05),红地球×玫瑰香2年的果皮硬度均符合正态分布。结合图1可以看出,果皮硬度在2个群体后代中均表现为连续性分布,属于典型的多基因控制的数量性状。通过对各杂交组合果皮硬度的遗传分析可以看出,2组合杂交后代的子代均值基本都低于亲中值(除红地球×玫瑰香2018年子代均值稍高于亲中值外),杂交果实果皮硬度的变异是向小的方向回归。2个杂交组合后代果皮硬度变异范围及变异幅度都较大,变异系数26.07%~34.21%,广义遗传力较高,在0.94~0.97,组合传递力都不低于85.32%。表明2个群体后代果皮硬度性状分离是比较广泛的,且果皮硬度差异主要受遗传因素影响。
2个组合后代中都出现了一定比率的超高亲和超低亲单株,但总体看来,超低亲率要高于超高亲率,红地球×玫瑰香2018年和2019年超高亲率分别为31.39%和21.97%,超低亲率分别为24.82%和28.03%;红地球×金星无核超低亲率分别为55.80%和35.82%,超高亲率分别为13.04%和23.13%(表1)。
2.2 2个杂交群体果皮破裂距离遗传分析
果皮破裂距离可以反映浆果形变度,也是很重要的质构参数。由图2可以看出,2个杂交组合后代的果皮破裂距离均呈现连续性分布,是多基因控制的数量性状,呈现正态分布。2个群体2018年和2019年果皮破裂距离子代平均值均高于亲中值,红地球×玫瑰香2年子代均值和亲中值分别为4.80、5.41 mm和4.12、4.76 mm,红地球×金星无核子代均值和亲中值分别为4.84、5.79 mm和4.08、4.12 mm;变异系数范围在19.11%~33.26%,变异范围较大,后代都呈现出广泛分离。2个群体均出现了较高的超高亲率,范围在31.06%~77.61%;超低亲率相对较低,都在15%以下,表明以红地球为母本,不同父本对果皮破裂距离变异程度有一定影响,但总体趋势一致。各杂交组合传递力都较高,均超过100%,广义遗传力范围在0.73~0.93,说明果皮破裂距离后代遗传传递能力较强,且主要受遗传因素影响。各年份间表型数据均呈显著性相关(P<0.001),红地球×玫瑰香2018和2019年相关性系数为0.69,红地球×金星无核2年间相关性系数为0.79(表2)。
2.3 2个杂交群体果肉硬度遗传分析
2个组合后代的果肉硬度均呈连续性分布,是典型的多基因控制的数量性状(图3)。红地球×玫瑰香杂交群体整体上呈现出向软肉回归的趋势,2年间果肉硬度呈显著的正相關(P<0.001),相关性系数为0.76。2018年和2019年亲中值分别为34.14、33.41 g,子代均值为31.27、32.62 g,组合传递力分别达到91.59%和95.94%,超高亲率10.37%和14.73%,超低亲率分别为17.78%和19.38%,变异系数为24.23%和26.00%,广义遗传均为0.91,说明该群体的果肉硬度差异主要受遗传因素影响较大。红地球×金星无核在果肉硬度遗传规律上与红地球×玫瑰香相似,2年间果肉硬度呈显著的正相关(P<0.001),相关性系数为0.63。
子代均值略小于亲中值,2年的变异系数均比前者大,说明该组合分离更为广泛,超低亲率较高,在30%以上,这个群体后代中果肉偏软的比率较高。组合遗传力分别为86.26%和89.53%,2年的广义遗传力均为0.93(表3)。
3 讨论与结论
葡萄在有性杂交过程中,受双亲基因间的加性和非加性效应的影响,后代性状往往出现广泛分离现象,形成不同的后代单株,也会产生一些超高亲植株,这也正是杂交育种的优势体现。果实质地是一个综合性状,在葡萄软化过程中,果皮硬度、果皮破裂距离、果肉硬度这3个穿刺参数可作为评价鲜食葡萄质地差异的关键质构参数。了解掌握葡萄果实质地遗传趋势与规律,合理选择选配亲本是缩短育种周期,提高育种效率的前提。有关果实质地遗传在葡萄中已有少量报道,但多数研究都针对的是单一质构参数或者是感官评价,标准多样、描述误差等造成了遗传分析的偏差。Carreo等分别以玫瑰香×绿宝石和红宝石无核×麝香葡萄的153株和78株杂交后代为试材,利用葡萄果实形变20%所用的力代表果实硬度,研究了果实硬度遗传特点,结果表明,2个群体杂交后代果实硬度均呈连续分布,属于典型的数量性状,并且认为2群体后代硬度均偏向于低值。Jiang等研究玫瑰香×克瑞森无核杂交群体果实硬度遗传规律,果实硬度以去皮后果实形变25%的最大力表示,得出群体后代的果实硬度呈近正态分布,连续变异,属于多基因控制的数量性状,并出现了一定数量的超亲后代。王勇等以红地球为母本的8个杂交组合的384个F植株为试材,利用感官描述评价,得出软肉型、中等、脆肉的比约为 5.3∶6.0∶1.0。认为红地球的脆肉性状传递给后代的比率较低,在遗传中呈减弱趋势,遗传效果也受到父本质地性状的影响。本试验通过对红地球×玫瑰香和红地球×金星无核2个杂交群体后代果皮硬度进行遗传分析得到了与前人类似的结果。2个群体的果皮硬度呈现向小回归的趋势,呈现正态分布的连续变异,认为果皮硬度是属于多基因控制的数量性状,并且控制果皮硬度的多基因产生加性效应,果皮硬度较大的红地球在杂交时非加性效应解体,果皮硬度出现了变小的倾向,且2个群体后代都出现了一定数量的超亲单株。
果皮破裂距离是质构仪进行整果穿刺试验过程中的一个重要的质构参数,迄今为止,还没有关于果皮破裂距离遗传规律的研究报道。本研究得出,2个杂交组合果皮破裂距离子代均值均高于亲中值,后代果皮破裂距离有趋大变异趋势,超高亲率较高,超低亲率较低。红地球×金星无核果皮破裂距离变异范围更广,出现了更多的超高亲植株,可能是种间杂交原加性效应解体,产生了新的加性效应和非加性效应。
有关果肉硬度遗传的研究在其他果树上报道较多,比如对杏果肉硬度遗传研究发现,杏果肉硬度属于数量性状遗传,且呈现偏向软肉亲本的连续变异。赵亚楠等对苹果梨与八月红210株F杂交后代研究得出,果肉硬度是由主效基因和微效多基因共同控制的,表现出超高亲遗传的倾向,说明在这个组合里高硬度性状遗传给后代的能力较强。研究者对秦冠、富士苹果杂交后代果实硬度遗传趋势的研究发现,秦冠的高硬度性状遗传给后代的能力较强,有偏硬遗传倾向,也有研究得出苹果后代果实硬度有低亲遗传的特点。由于葡萄的浆果特性,以往通过直接测定果肉硬度进行遗传研究的较少,本试验通过整果穿刺直接测定果肉平均硬度,分析发现杂种后代果肉硬度呈连续性分布,是典型的多基因控制的数量性状,广义遗传力较大,主要受遗传因素影响,这2个组合果肉硬度子代均值都小于中亲值,有向小回归的趋势,加性效应解体,与前人研究结果一致。但与一些研究结果相反,这可能与树种、品种差异有关。
参考文献:
[1]Dai Z W,Vivin P,Barrieu F,et al. Physiological and modelling approaches to understand water and carbon fluxes during grape berry growth and quality development:a review[J]. Australian JournalOf Grape and Wine Research,2009,16:70-85.
[2]刘 俊,晁无疾,亓桂梅,等. 蓬勃发展的中国葡萄产业[J]. 中外葡萄与葡萄酒,2020(1):1-8.
[3]Coombe B G. ResearchOn development and ripeningOf the grape berry[J]. American JournalOf Enology and Viticulture,1992,43(1):101-110.
[4]Carreo I,Cabezas J A,Martínez-Mora C,et al. Quantitative genetic analysisOf berry firmness in table grape(Vitis vinifera L.)[J]. Tree Genetics & Genomes,2014,11(1):1-10.
[5]王 勇,伍國红,李玉玲,等. ‘红地球’葡萄杂交F代重要果实性状遗传倾向分析[J]. 果树学报,2015,32(6):1099-1106.
[6]Jiang J F,Fan X C,Zhang Y,et al. ConstructionOf a high-density genetic map and mappingOf firmness in grapes(Vitis vinifera L.) basedOn whole-genome resequencing[J]. International JournalOf Molecular Sciences,2020,21(3):797.
[7]张翔宇,李喜宏,王妍丹,等. 穿刺测试和 fisher 判别法在葡萄贮藏期间质地评价的应用[J]. 食品工业科技,2017,38(3):342-347,352.
[8]Letaief H,Rolle L,Zeppa G,et al. AssessmentOf grape skin hardness by a puncture test[J]. JournalOf the ScienceOf Food and Agriculture,2008,88(9):1567-1575.
[9]朱 军. 运用混合线性模型定位复杂数量性状基因的方法[J]. 浙江大学学报(自然科学版),1999,33(3):327-335.
[10]徐 铭,刘威生,王爱德,等. 杏主要经济性状遗传分析[J]. 果树学报,2020,37(1):1-10.
[11]刘家成,章秋平,牛铁泉,等. ‘串枝红’与‘赛买提’杏正、反交后代果实性状遗传倾向分析[J]. 果树学报,2020,37(5):625-634.
[12]赵亚楠,欧春青,王 斐,等. 梨F代群体果实性状的遗传分析及相关性研究[J]. 西北农业学报,2018,27(12):1811-1818.
[13]王亚杰,孟 蕊,武月妮,等. 秦冠、富士苹果杂交后代果实性状遗传趋势分析[J]. 西北农业学报,2014,23(4):52-59.
[14]李林光,王宏伟,李慧峰,等. 皇家嘎啦苹果与藤牧一号杂交后代果实性状遗传研究[J]. 落叶果树,2009,41(1):4-6.
