重金属铅镉对洋葱根系生长的影响
2022-06-21张尔珂周贵寅张玉平邓新辉
张 涵 张尔珂 周贵寅 张玉平 邓新辉 谭 平
湖南工业大学 生命科学与化学学院 湖南 株洲 412007
0 引言
重金属对水体及土壤的污染也日益成为人们关注的问题[1]。调查显示,全国土壤重金属总超标率为16.1%,污染类型以无机型为主,土壤及水体重金属离子主要有铅、汞、镉等8种[2-5]。采矿污染土地面积已达200余万公顷,并且每年以(3.3~4.7)万公顷的速度递增[6]。西南地区土壤中,镉、铅、锌、铜、砷等背景值远远高于全国土壤背景值[7-10]。因此,研究土壤中重金属对植物生长的影响可为解决当前环境污染问题提供依据。
植物的根系直接与土壤相接触,植物对重金属有较强的富集能力[11-12]。当土壤中重金属浓度较高时,植物根系对重金属的富集作用会受到抑制,同时其生长发育也会受到影响[13-15]。双龙等[16]研究发现环境中重金属浓度过高会降低植物根系的含水量。P. Sharma等[17]的研究表明,当重金属的含量超过作物阈值,会对植物体内相关酶造成影响,进而影响植物生长发育,甚至导致植物死亡。在重金属胁迫下,植物生长状况恶化,根系生长受到抑制、根系体积减小,根鲜重降低[16]、根系活力减小,最终导致其产量、质量降低,生物多样性减少[17-20]。M. Jamla等[21]研究发现不同种类植物对重金属的耐受能力不同,木本植物对重金属有较强的耐性。国内外对重金属危害植物的研究层出不穷,但对重金属胁迫下,植物生长机理的研究相当有限。
鉴于洋葱根系对重金属铅、镉较为敏感[22],本研究采取水培试验的方法,以洋葱为供试植物,在模拟重金属铅、镉污染条件下,分析重金属类型和浓度对洋葱根系生长状况的影响,利用显微镜观察重金属Pb、Cd在不同浓度下对洋葱根尖细胞有丝分裂过程、细胞核形态的影响,通过测定吸光度值和显色反应的方法评估重金属铅、镉对过氧化物酶(peroxidase,POD)和过氧化氢酶(catalase,CAT)活力[23]等的影响。综合不同浓度重金属铅、镉对洋葱根系生长状况、有丝分裂过程、酶活力大小等各项生理指标的影响,初步探讨重金属对洋葱根系的影响机制,阐明洋葱对重金属铅镉胁迫下的生理响应机制,为重金属对植物生长机理的影响和重金属污染土壤的生态修复提供理论依据。
1 实验
1.1 供试植物
供试植物选取大小、生长状态基本一致,较新鲜,无病虫害及腐烂现象,符合实验要求的新鲜紫皮洋葱,购于湖南省株洲市蔬菜市场。试验前先用自来水将洋葱根部表面的土壤和污渍洗去,避免破坏洋葱的内部结构,再用蒸馏水润洗1~2次。
1.2 实验试剂、设备与仪器
1)实验试剂
硝酸铅(Pb(NO3)2)、硝酸镉(Cd(NO3)2)、酒精(体积分数为70%)、盐酸(体积分数为10%)、Carnoy固定液(甲醇与冰乙酸体积比为3:1)、磷酸缓冲液、愈创木酚、过氧化氢、硫酸,以上试剂均为分析纯,购于国药集团化学试剂有限公司。去离子水,实验室自制。
2)实验设备与仪器
超纯水处理系统,YK-RO-B型,上海和泰仪器有限公司;加热磁力搅拌器,HJ-85-2型,郑州长城科工贸有限公司;高速离心机,TG16G型,湖南凯达科学仪器有限公司;电热恒温水浴锅,HH-2型,北京市永光明医疗仪器有限公司;精密电子天平,BSM120.4型,上海舜宇恒平科学仪器有限公司;光学显微镜,xsp-01型,上海浦丹光学有限公司;紫外-可见分光光度计,通用T6新世纪,北京普析通用仪器有限公司。
1.3 实验方法
将两种重金属试剂Cd(NO3)2和Pb(NO3)2配制成不同浓度的水溶液,共设置9个实验组,将去离子水作为空白对照组;每个浓度设置3个平行实验,采用水培法培养洋葱,共30个样品。每天上午八点至九点统计每个洋葱根的数量,并对洋葱每条根的长度进行测量并记录。7 d后收集不同重金属浓度下培养的洋葱根尖,观察洋葱根尖的有丝分裂情况,并对过氧化物酶和过氧化氢酶活力进行测定。
1.4 观察与测定方法
1)洋葱根尖细胞有丝分裂的观察。剪下洋葱根尖约0.5 cm,置于固定液中固定20 min后转入酒精中,放置片刻将根尖取出,水洗后放入盛有少量10%盐酸的小烧杯中,并置于60 ℃恒温水浴锅中解离8 min,直至根尖发白变软。用镊子将根尖伸长区部分取出,在盛有清水的玻璃皿中漂洗后,置于载玻片上。夹住根尖,用解剖针或刀片在根尖乳白色部分0.3 cm处剔除1/3至1/2,然后放在载玻片中央,将所取的材料充分研磨、并用醋酸洋红染液染色,用解剖针将组织分散,染色5~20 min。吸去多余染液,盖上盖玻片,在盖玻片上放2层吸水纸,对准标本扣压,使细胞和染色体铺展开。先用低倍镜观察洋葱根尖压片,选择分裂相较多的部位,再转至视野中央,利用高倍镜进行观察,拍照记录观察结果。
2)过氧化物酶活力的测定。称取洋葱根系1 g,量取5 mL 20 mmol/L KH2PO4溶液置于研钵中研磨、匀浆,以4000 r/min转速离心15 min,收集上清液保存在冷处,残渣用5 mL KH2PO4溶液提取一次,合并两次提取液,取上清液。在比色皿中加入3 mL愈创木酚反应混合液,1 mL KH2PO4溶液作对照;另一比色皿中加入3 mL反应混合液与1 mL酶液,立即用秒表计时,在紫外-可见分光光度计470 nm处测定吸光度值,每隔1 min测定一次,并记录数据,以每分钟内吸光度变化为一个酶活力单位(U)。
3)过氧化氢酶活力的测定。取足量洋葱根系,加入少量pH=7.8的磷酸缓冲溶液,研磨成匀浆,转移至10 mL离心管中,用该缓冲液冲洗研钵,并将冲洗液转至管中,10 000 r/min离心12 min,上清液即为过氧化氢酶的粗提液。取若干小试管,加入2.5 mL酶液,再加入2.5 mL 0.1 mol/L H2O2,于30 ℃恒温水浴中反应10 min,立即加入2.5 mL 10% 硫酸溶液停止反应,用0.1 mol/L高锰酸钾溶液滴定,至出现粉红色(30 s内不消失)为终点,停止滴定并记录消耗的KMnO4溶液体积,酶活力用每克鲜重样品在10 min内分解H2O2的毫升数表示,酶活力计算参照实验教材的通用公式。
1.5 数据分析
所有实验数据利用Excel 2003进行平均值计算,利用数据统计软件Origin对洋葱根系长度和生长时间进行Logistics生长模型拟合。
2 结果与分析
2.1 重金属对洋葱根系生长状况的影响
2.1.1 重金属对洋葱根系形态的影响
为了研究重金属类型和浓度对洋葱根系生长状况的影响,在不同浓度Pb、Cd下观察洋葱根系的生长情况,结果如图1所示。

图1 不同溶液中洋葱根系的生长状态Fig. 1 Growth situation of onion roots in different solutions
由图1可知,不同浓度的Pb(NO3)2及Cd(NO3)2溶液中洋葱根系主要出现了以下4种具有代表性的现象:图1a在60 mg/L的Cd(NO3)2、2000 mg/L的Pb(NO3)2溶液中,部分洋葱根系表面发黑,根系短而粗;图1b在20 mg/L Cd(NO3)2溶液中洋葱的根尖变黄变弯曲;图1c在40 mg/L的Cd(NO3)2、1000 mg/L 的Pb(NO3)2溶液中,部分洋葱根系出现变黄、变软、变透明弯曲等现象;图1d在3000 mg/L的Pb(NO3)2溶液中,部分洋葱出现了根部腐烂的现象。这是由于铅对根尖细胞产生严重毒害,高浓度Pb(NO3)2使洋葱根系腐烂,高浓度的Cd(NO3)2加速根尖细胞木质化[24-25]。洋葱对重金属铅、镉胁迫均较为敏感,可作为重金属铅、镉的监测植物。
2.1.2 重金属对洋葱根系长度的影响
不同浓度Cd(NO3)2溶液中,一周内洋葱根系长度变化如表1所示。
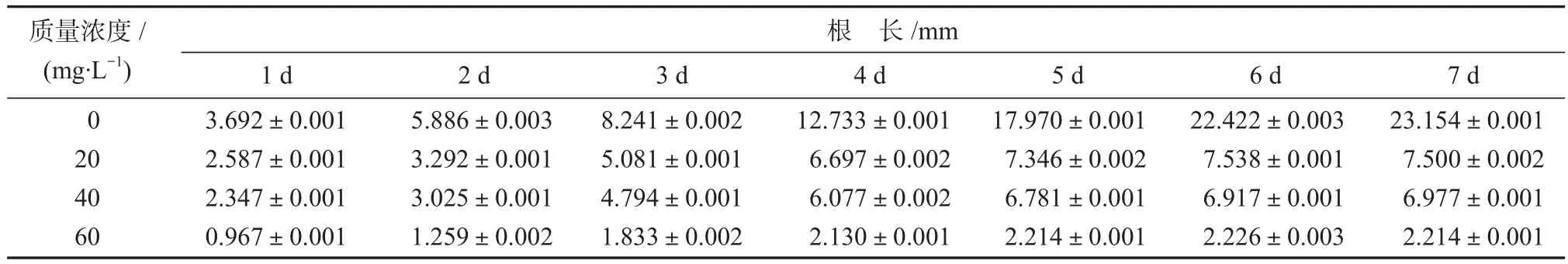
表1 不同浓度Cd(NO3)2中洋葱根系长度Table 1 Root length of onion in Cd(NO3)2 with different concentrations
由表1可知,随着Cd(NO3)2浓度的增大,洋葱根系平均长度均减小,且与对照组相差越大。当Cd(NO3)2质量浓度为60 mg/L时,培养1 d后根系长度与对照组的相差2.725 mm,第2~7 d分别相差4.627,6.408, 10.603, 15.756, 20.196, 20.940 mm,时间越长根长差别越大。从组内来看,对照组7 d内根系平均生长速率为3.308 mm/d,当Cd(NO3)2质量浓度为20,40, 60 mg/L时,洋葱根系平均生长速率分别为1.071,0.997, 0.316 mm/d。高浓度的Cd(NO3)2加速根尖细胞木质化,Cd(NO3)2浓度越大,越不利于洋葱根系生长。
表2为不同浓度Pb(NO3)2溶液中一周内洋葱根系的长度变化。由表2可知,整个培养期间,随着Pb(NO3)2浓度的增大,洋葱根系生长越缓慢,平均长度越小,与对照组相差不断增大。当Pb(NO3)2质量浓度达到3000 mg/L时,培养1 d后与对照组相差3.692 mm,第2~7 d分别相差5.886, 8.191, 11.858, 16.970,21.422, 22.154 mm,时间越长根长差别越大。从组内来看,当Pb(NO3)2质量浓度为1000, 2000, 3000 mg/L时,洋葱根系平均生长速率分别为0.302, 0.167, 0.143 mm/d,其中3000 mg/L的Pb(NO3)2溶液中洋葱根尖从第5 d起停止生长,这与高浓度Pb(NO3)2使洋葱根系腐烂有关。

表2 不同浓度Pb(NO3)2中洋葱根系长度Table 2 Root length of onion in different concentrations of Pb(NO3)2
2.1.3 洋葱根系Logistics生长模型
利用Origin对不同溶液中洋葱根系长度和生长时间进行Logistics生长模型拟合,结果如图2所示。
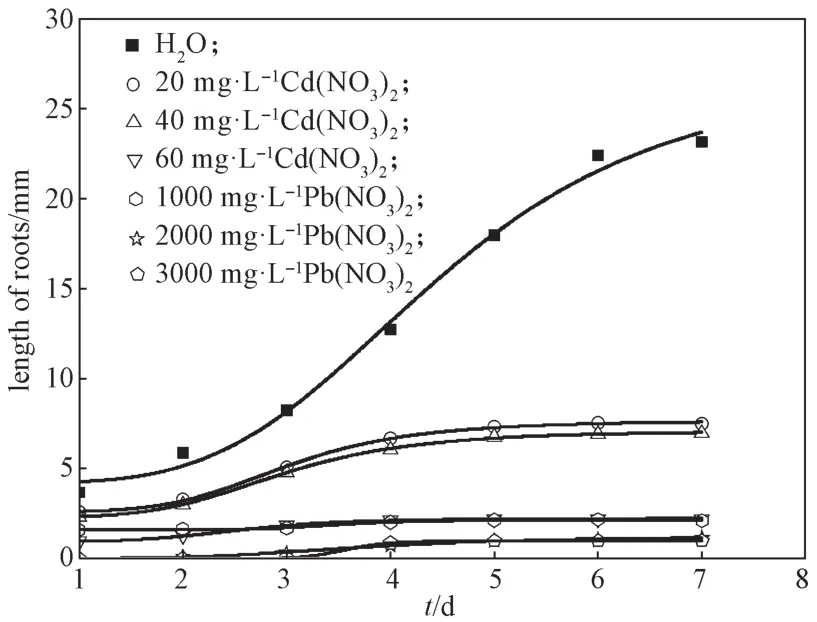
图2 洋葱根系生长状况的Logistics方程拟合Fig. 2 Logistics equation fitting effect of onion root growth
由图2可知,在Cd(NO3)2、Pb(NO3)2溶液中,洋葱根系生长状态均符合Logistics生长模型。Cd(NO3)2和Pb(NO3)2浓度越大,洋葱根系平均长度越短。在60 mg/L Cd(NO3)2溶液中培养7 d后,洋葱根系最短,平均长度为2.214 mm;在相同的培养时间内,Pb(NO3)2质量浓度为3000 mg/L时,洋葱根系最短,平均长度为1.000 mm。洋葱水培7 d后,根系长度基本稳定,可作为实验结果反映洋葱根系生长情况。
表3为不同浓度Cd(NO3)2、Pb(NO3)2溶液中根系长度与生长时间Logistics方程拟合相关系数。

表3 不同溶液中Logistics方程拟合相关系数Table 3 Fitting correlation coefficient of Logistics equation in different concentrations of solution
由表3可知,在任何一种溶液中相关系数均大于0.99,符合相关系数R2≥0.99,洋葱根系生长均符合Logistics生长模型。
2.2 重金属对洋葱根尖有丝分裂的影响
2.2.1 镉对洋葱根尖有丝分裂的影响
为了深入研究不同重金属的对洋葱根系的影响,利用显微镜观察在不同浓度Cd条件下洋葱根系细胞的形态变化,结果如图3所示。
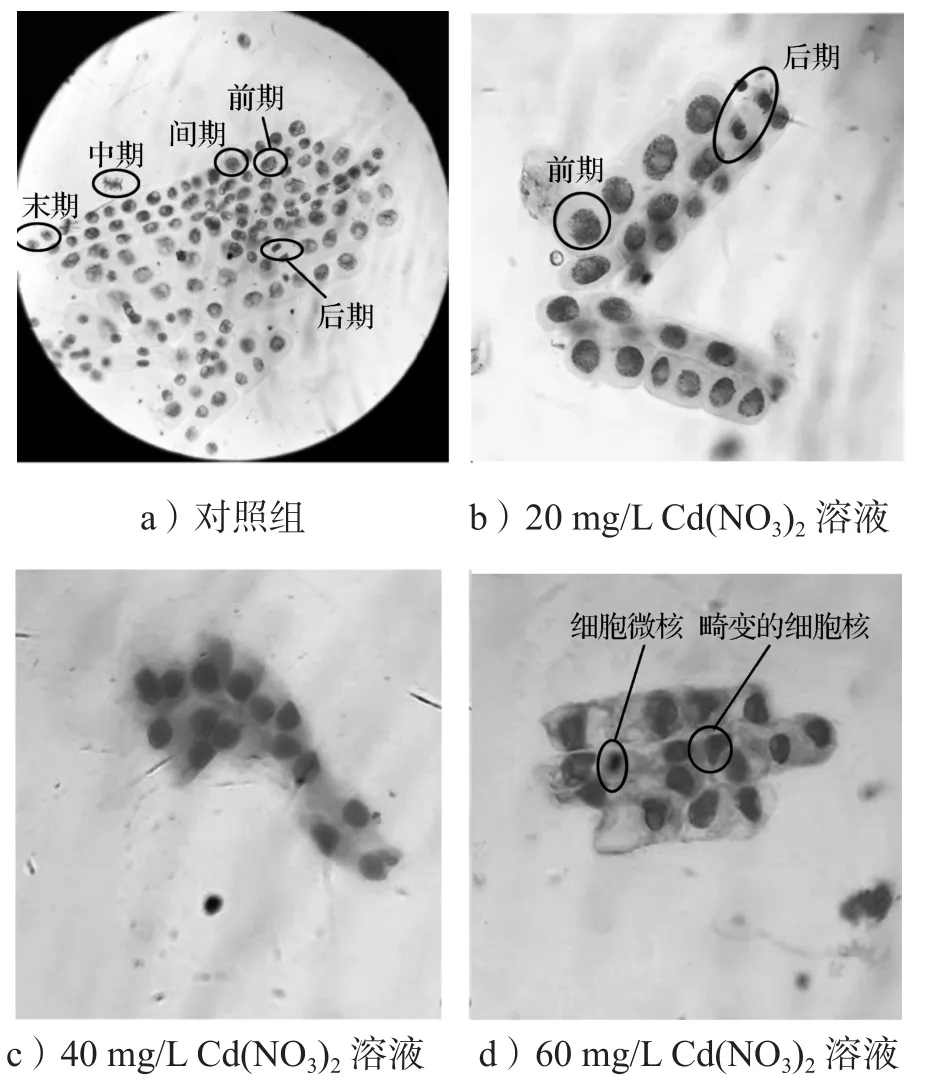
图3 不同浓度Cd(NO3)2溶液中洋葱根尖细胞有丝分裂Fig. 3 Mitosis of onion root tip cells in different concentrations of Cd(NO3)2 solution
由图3可知,对照组的洋葱根尖细胞有丝分裂各个时期均正常;在20 mg/L Cd(NO3)2溶液中,细胞形态完好,染色均匀,此浓度下根尖细胞主要处于有丝分裂前期,细胞分裂不够旺盛;在40 mg/L Cd(NO3)2溶液中,细胞核完好,根尖细胞主要处于分裂间期;在60 mg/L Cd(NO3)2溶液中,根尖细胞处于分裂间期,细胞核畸变,呈不规则形状,同时出现细胞微核。
2.2.2 铅对洋葱根尖有丝分裂的影响
利用显微镜观察在不同浓度Pb条件下洋葱根系细胞的形态变化,结果如图4所示。
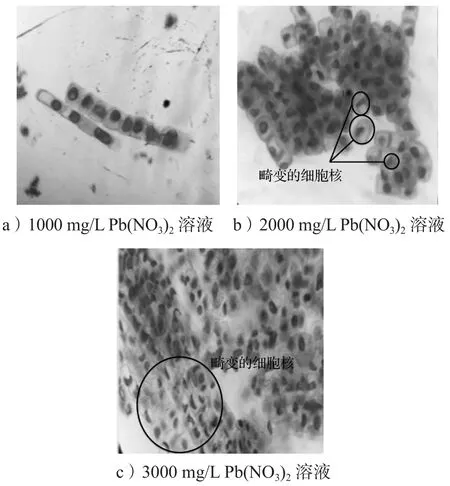
图4 不同浓度Pb(NO3)2溶液中洋葱根尖细胞有丝分裂Fig. 4 Mitosis of onion root tip cells in different concentrations of Pb(NO3)2 solution
由图4可知,在1000~2000 mg/L Pb(NO3)2溶液中培养7 d,部分洋葱根尖细胞的细胞核畸变,呈现不规则的形状。在3000 mg/L Pb(NO3)2溶液中,洋葱根尖细胞核由圆形或椭圆形变为半月牙状,根尖细胞的细胞核、染色体等结构受到破坏,培养后期根尖细胞停止生长甚至出现根系腐烂现象。这主要是因为Cd(NO3)2、Pb(NO3)2可抑制洋葱根尖细胞纺锤体的形成,从而抑制根尖细胞的有丝分裂,导致根系生长速度减慢,甚至使根系细胞停止生长。
2.3 重金属对洋葱根尖过氧化物酶的影响
通过计算得出不同浓度Cd(NO3)2和Pb(NO3)2溶液中培养的洋葱根系的POD酶活力,再以Cd(NO3)2、Pb(NO3)2质量浓度为横坐标,POD酶活力为纵坐标作图,结果如图5所示。
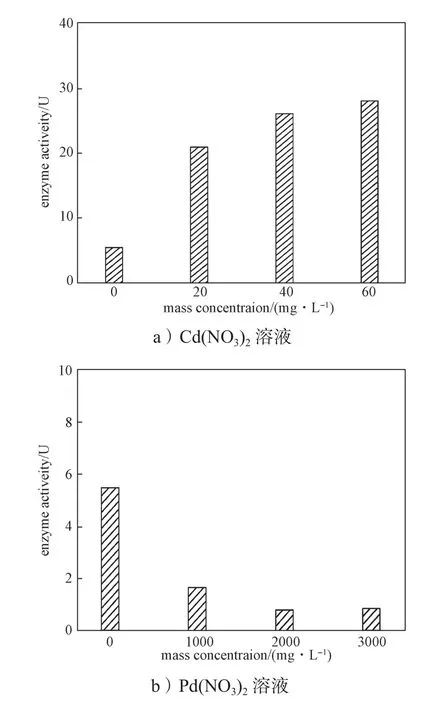
图5 不同浓度溶液中洋葱根系POD酶活力Fig. 5 POD activity of onion roots in different concentrations of solution
由图5a可知,当Cd(NO3)2的质量浓度在0~60 mg/L范围内时,POD酶活力与Cd(NO3)2浓度呈正相关,说明在此浓度范围内Cd(NO3)2对POD酶具有激活作用,也就是说此浓度范围为Cd(NO3)2对洋葱根尖细胞毒害所能承受的浓度范围。当Cd(NO3)2质量浓度为60 mg/L,洋葱根系的过氧化物酶活力最大,为28.0068 U。由图5b可知,POD酶活力随Pb(NO3)2浓度增大,呈现降低趋势。当Pb(NO3)2质量浓度为3000 mg/L时,其POD酶活力最小,为0.8 U。高浓度的Pb(NO3)2对洋葱根尖细胞POD酶活力有抑制作用,低浓度Cd(NO3)2溶液可激活POD活力。
2.4 重金属对洋葱根尖过氧化氢酶的影响
通过计算得出CAT酶活力,以Cd(NO3)2、Pb(NO3)2质量浓度为横坐标,POD酶活力为纵坐标作图,结果如图6所示。
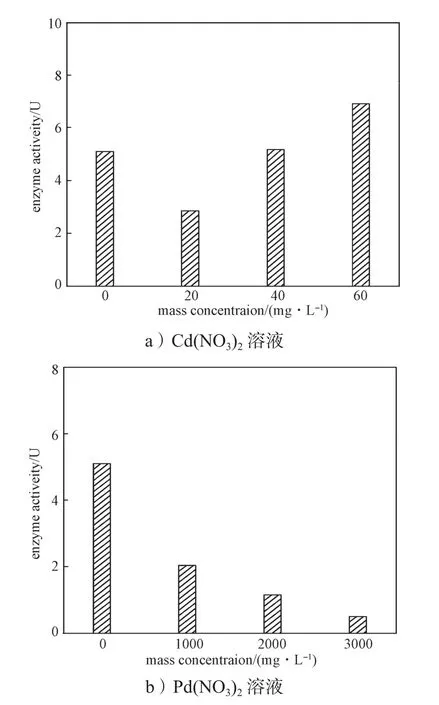
图6 不同浓度溶液中洋葱根系CAT酶活力Fig. 6 CAT activity of onion root in different concentrations of solution
由图6a可知,当Cd(NO3)2质量浓度为20 mg/L时,洋葱根尖细胞CAT酶活力较对照组的低;当Cd(NO3)2质量浓度为40 mg/L时,洋葱根尖细胞CAT酶活力与对照组的相当;当Cd(NO3)2质量浓度为60 mg/L时,洋葱根尖细胞CAT酶活力高于对照组的,即高浓度的Cd(NO3)2对CAT酶具有激活作用。这是因为植物体具有对外界环境的应激性,实验中重金属镉可能抑制了洋葱H2O2的分解或促进了洋葱产生H2O2,使得洋葱根系细胞中积累了部分H2O2[26],为分解过多的H2O2,从而使得CAT酶活力上升。由图6b可知,在Pb(NO3)2胁迫下,各浓度CAT酶活力均低于对照组的,在Pb(NO3)2质量浓度在1000~3000 mg/L范围内,CAT酶活力呈现逐渐减少的趋势。这是因为外界重金属浓度升高会使洋葱对其产出应激反应,促使CAT酶活力升高,但由于重金属的毒性对酶造成的损害更大,酶活力虽有增加趋势仍不能抵消重金属的毒害作用,无法达到对照组的CAT酶活力。对比图6a~b可知,Pb(NO3)2对CAT酶活力影响更明显[27-28]。
3 结论
本研究通过模拟重金属铅、镉污染环境,研究在Pb(NO3)2、Cd(NO3)2胁迫下洋葱根尖形态、生长及根尖细胞有丝分裂、细胞POD和CAT酶活力等方面的变化,从形态和生理方面揭示了洋葱根尖对重金属Pb(NO3)2、Cd(NO3)2胁迫的响应特征。研究发现,重金属Pb(NO3)2、Cd(NO3)2对洋葱根系的生长均具有抑制作用,可抑制根尖细胞的有丝分裂,使细胞停留在有丝分裂的前期或间期;抑制作用均随着浓度的增大而增强。在一定浓度范围内Cd(NO3)2能激活洋葱根尖细胞的过氧化物酶体系,促使洋葱对重金属Cd(NO3)2的胁迫呈现出积极的生理响应特征。较高浓度的Pb(NO3)2、Cd(NO3)2溶液,使细胞无法进行正常生命活动、停止生长甚至腐烂。本研究通过分析重金属铅、镉对洋葱根系长度、有丝分裂过程、酶活力等各项生理指标的影响及其影响机制,为重金属对植物生长机理的影响和重金属污染土壤的生态修复提供理论依据。
