茶树富集氟的特点及其机制的研究进展
2022-06-21邢安琪武子辰徐晓寒孙怡王艮梅王玉花
邢安琪,武子辰,徐晓寒,孙怡,王艮梅,王玉花*
茶树富集氟的特点及其机制的研究进展
邢安琪1,武子辰1,徐晓寒1,孙怡1,王艮梅2,王玉花1*
1. 南京农业大学园艺学院,江苏 南京 210095;2. 南京林业大学南方现代林业协同创新中心,江苏 南京 210037
茶树是一种超累积氟的植物,其体内的氟含量远远高于其他植物,却不表现氟中毒症状。氟不是茶树生长的必需元素,但在高氟胁迫下,氟可通过破坏茶树的细胞结构、抑制酶活性等影响其正常生长。通过阐述茶树吸收、富集氟及其累积/解毒机制等方面的研究进展,以期为未来研究茶树氟累积和降氟技术提供理论依据。
茶树;氟;富集机制
氟是植物生长过程中的非必需元素,根据植物对氟污染反应的不同可以分为敏感、中等敏感和抗性3种类型[1]。其中,茶树属于氟抗性植物,且叶片是茶树氟富集的主要器官[2]。闽南茶树的一芽三叶中氟含量大致范围在69~299 mg·kg-1[3],四川茶园茶树叶片的氟含量可高达2 965 mg·kg-1[4],安徽茶园茶树的成熟叶片氟含量也高达1 155.2 mg·kg-1[5]。针对闽南采集的55份茶树样本,通过组间平均联结法对茶叶氟含量的差异进行聚类分析得出3个类群,分别为低累积、中度累积和高累积氟元素的茶树类型[3]。可见这些茶树叶片氟含量的差异与品种紧密相关,并且这些茶树在存在一定浓度氟的生长环境中并未呈现受伤害的状况。但当植物氟含量超过耐受范围时,植物损伤最先出现在叶片上,即叶缘和叶尖坏死,随后出现叶片萎黄甚至坏死等一系列症状[6]。
茶树作为叶用植物,其叶片的氟含量过高,会对茶叶的儿茶素、氨基酸、香气等品质成分产生不利影响[7-8]。此外,茶叶在冲泡过程中约有42%~86%的氟溶于茶汤,进而被人体吸收,长期积累,会增加人体患“饮茶型氟中毒”的风险[9]。因此,了解茶树吸收、累积氟的特点,可以针对性地提出有效降低茶树累积氟的措施,减少茶叶的氟含量,对提高茶叶品质和质量安全具有重要意义。
1 茶树吸收、富集氟的特性
1.1 茶树对氟的吸收
在氟污染的大气环境中,茶树主要通过叶片气孔或表皮角质层吸收大气中的氟,并且可沿导管向叶片尖端和边缘转移(图1)。
茶树主要通过根系吸收土壤和水环境中的氟,并通过蒸腾作用、质外体和共质体途径等进行氟转运[10]。王玉梅等[11]通过研究茶树根系对氟的吸收发现,在0~5.0 mmol·L-1的氟处理条件下茶树对氟的吸收符合Michalis-Menten动力学模型。张磊等[12]也发现,茶苗在0~2.0 mg·L-1的氟处理下其吸收动力学曲线符合Michalis-Menten动力学模型;在氟浓度为0~0.8 mg·L-1时,茶苗吸收速率随外界氟浓度增加而迅速增加;而在氟质量浓度为0.8~2.0 mg·L-1时,其吸收速率趋于稳定。张磊[13]认为,茶树对氟的吸收,在氟浓度较低时为主动吸收过程,而在氟浓度较高时以被动吸收为主导。彭传燚等[14]对茶树吸收氟的浓度动力学特性研究表明,在0.1~10 mg·L-1的氟质量浓度范围内,茶树根系对氟离子的吸收是主动吸收,并符合Michaelis-Menten动力学模型,而在高氟条件(50~100 mg·L-1)下则表现出被动吸收氟的过程。此外,茶树主要吸收土壤中的水溶性氟。诸多研究表明黔西北高氟病区[15]、安徽[5]、粤东凤凰山茶区[16]、四川、重庆、云南、湖北、湖南茶园[17]等不同产茶区土壤的水溶性氟含量均低于10 mg·kg-1。因此,我国大多数茶园茶树的氟吸收主要依赖于根系的主动吸收。
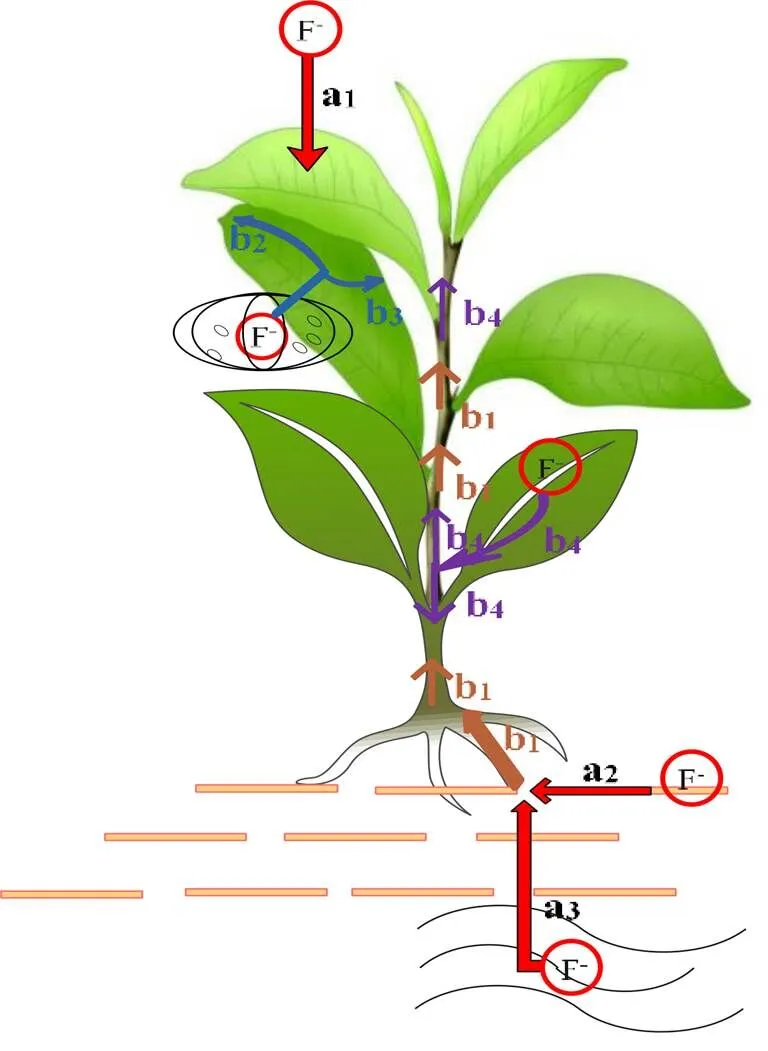
注:茶树吸收来自空气、土壤、大气(a1、a2、a3)中的氟(F);茶树根部吸收的F可向地上部转移并贮存(b1);沿导管向叶片尖端和边缘转移并积累(b2和b3);老叶中的F可向新梢和地下部转移(b4)
主动吸收需要的能量主要通过H+-ATPase产生,王玉梅[18]研究发现,外源施加NaF可以激发H+-ATPase的活性,刺激质子驱动力的形成,促进茶树根系对F的吸收。但H+-ATPase介导的茶树根系跨膜主动吸收氟的分子机制仍有待研究。Ca2+-ATPase介导的茶树根系跨膜主动吸收氟主要是通过激活类受体蛋白激酶(RLK)识别氟,激活Ca2+-ATPase活性促进茶树吸收氟[19](图2)。而目前关于茶树吸收氟分子机制的研究较多针对离子通道蛋白,尤其是近来发现的特定吸收转运氟的离子通道[20],CLC超家族阴离子转运蛋白的一个亚类——CLCF型F-/H+反向转运蛋白,以及在原核生物中称作“crcB(Fluc)”,但在真核生物中称为FEX(Fluorie exporter)的小膜蛋白家族。茶树的CLC基因对F-的响应更敏感[21];茶树氟化物输出基因FEX具有组织特异性,并在茶树的氟吸收转运中起重要作用[22]。王玉梅[18]研究表明,ABC(ATP-binding cassette)转运蛋白主要是ABC中A—G亚家族在茶树F/C1跨膜转运中发挥载体作用。广敏[23]研究发现,CsCL667蛋白含有典型的ABC转运蛋白结构域,并且和茶树CsFEX属于同一基因家族的不同分支;氟不仅诱导该基因的表达量,同时也显著增加了ABC转运蛋白的含量,而且ABC转运蛋白对氟的转运能力增强,大量的氟吸收进入细胞内累积。研究茶树氟富集首先需探明茶树如何吸收氟,虽然目前已有研究对相关基因进行了克隆及异源表达验证,但有关茶树吸收氟的分子机制仍需探索。
1.2 影响茶树吸收及转运氟的因素
茶树根系对氟的吸收转运受氟的存在形态、pH、其他元素(如Al3+、Ca2+、Cl-等)离子通道、离子通道抑制剂及温度等因素的影响。
氟在土壤中主要以难溶态、可交换态和水溶态等形式存在[17],其中难溶态和可交换态氟难以被茶树吸收,水溶态则是茶树吸收氟的主要形式,而水溶性氟浓度受pH的影响显著。茶树适宜生长在pH为4.0~6.5的酸性环境中,当培养液pH在4.0~6.0时,茶树根系的氟吸收量随pH的增加呈现先增加后减少的变化,并且在pH为5.5时达到最大吸收量[24]。在酸性条件下F-优先与Al3+形成络合物,并以该络合物为主要形式被茶树吸收[25]。有学者针对茶树在木质部的转运提出两种不同的氟转运形式,一种认为茶树分别转运铝和氟[26]至叶片后再进行积累;另一种则认为茶树直接以氟铝络合物形式转运[27]。铝同样在缓解茶树氟胁迫中起重要作用。当Al/F摩尔浓度比小于1时,氟将导致茶树叶片坏死,而Al浓度的适当增加,茶树会通过形成Al-F络合物减少游离氟存在的形式缓解茶叶的氟损害[25],以此增强茶树对氟的抗性,这可能是茶树氟富集的生理机制之一。此外,Ca2+与F-会形成CaF2沉淀,进而降低土壤水溶性氟的含量,影响茶树对氟的吸收[28]。也有报道发现,外源施加0.01~0.5 mmol·L-1Ca2+时,能够有效降低茶叶中的氟含量[29],这可能与Ca2+改变了茶树根系细胞壁结构和膜通透性有关[24]。另一方面,一些相同价态的阴离子也会对茶树体内的F-产生影响,例如Cl-和F-可能通过竞争同一载体的吸附位点,从而两者间存在竞争性抑制作用,因此Cl-会降低茶树对F-的吸收,同时F-也会干扰Cl-的吸收转运[11]。Zhang等[30]发现,施用阴离子通道抑制剂NPPB[5-硝基-2-(3-苯丙胺)-苯甲酸]会造成茶树的氟累积量减少,并且通过比较外施阴离子通道抑制剂、阳离子通道抑制剂、水通道抑制剂的研究发现[31],外施阴离子通道抑制剂可以更显著地减少茶树根系对氟的吸收。因此,部分氟可通过水通道蛋白被吸收,而阴离子通道则是茶树根系吸收氟的主要通道(图2)。
茶树根系对氟的主动吸收依赖于质膜H+-ATPase水解ATP形成的电势差和能量[32]。在低温(4℃)条件下,茶树根系质膜H+-ATPase的活性被抑制,茶树根系对氟的吸收也受到显著的抑制[11];高温(35℃)处理时,则明显提高根系对氟的吸收[33]。氟的吸收还与Ca2+-ATPase相关,受体蛋白激酶(RLK)通过活化Ca2+-ATPase并将其作为茶树吸收氟的载体,进而促进氟的吸收[19]。与此同时,Al3+处理可以诱导茶树根系产生Ca2+信号,进而激活钙调蛋白(CaM),促进氟的吸收[32]。另一方面,质膜H+-ATPase产生的H+梯度,也可以促进植物体内Ca2+信号传导,以此调控离子的跨膜转运[34],影响氟吸收。
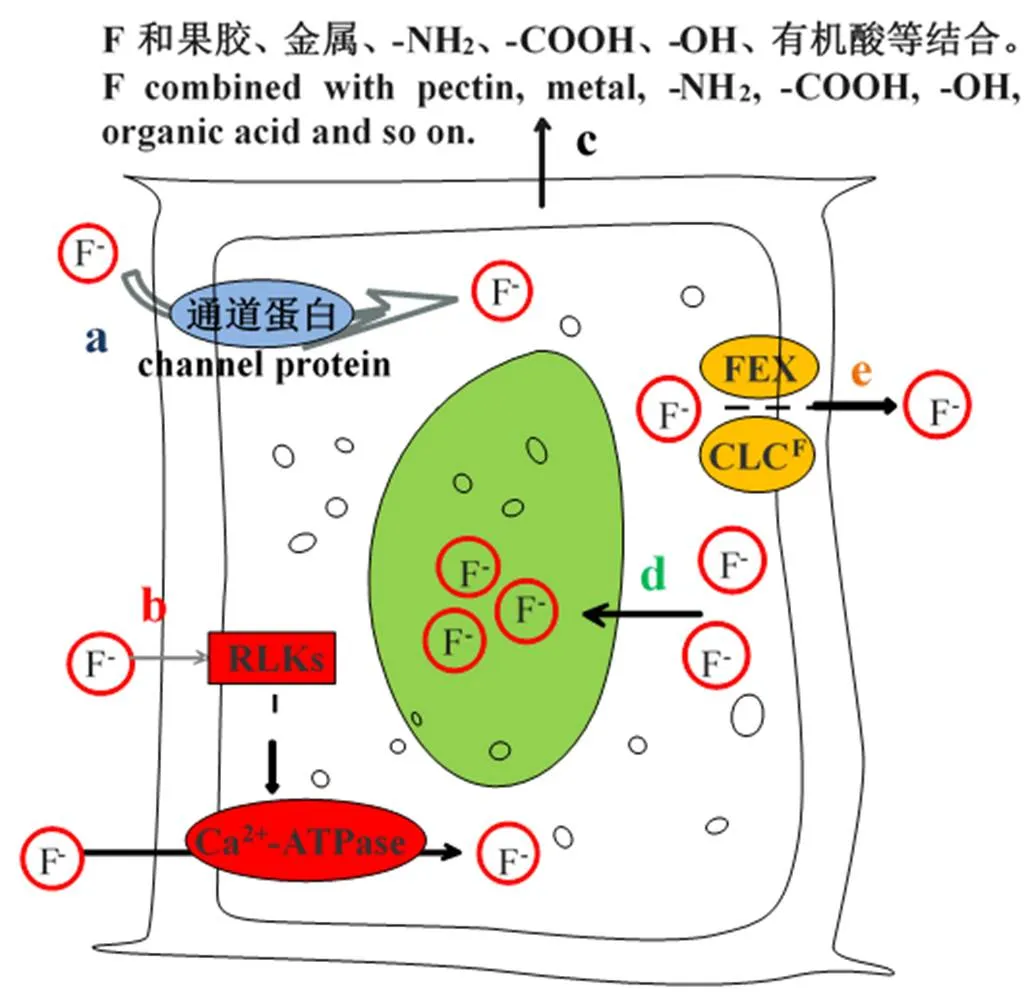
注:茶树通过水通道蛋白吸收F(a);F刺激RLKs表达,进一步激活Ca2+-ATPase,同时Ca2+-ATPase本身作为载体吸收F (b);细胞壁中的果胶及其周围的金属离子等与F螯合(c);液泡贮存F(d);通过FEX、CLCF等离子通道外排F(e)
1.3 茶树富集氟的特性
茶树各器官对氟的积累强度有很大不同,其中叶片的氟含量远大于吸收器官根和运输器官茎。沙济琴等[2]发现茶树各器官的氟积累强度顺序为叶>花蕾>茶籽>茶果皮>细枝>骨干枝>细根>主轴茎>主干茎>主根>侧根,唐茜等[35]的研究也证实了茶树各组织的聚氟能力依次为叶片>新梢>枝条>茎>根。此外,不同成熟度的茶树叶片氟含量差异显著,一般表现为成熟叶>嫩叶[5]。李丽霞等[36]发现,处于无氟水培条件下的茶树,其吸收根和新梢的氟含量增加,并认为这是由于茶树体内的氟进行了转移与再分配的结果。茶树体内的氟具有可移动性且这种转移特性决定了氟在新梢中的含量变化,同时发现氟可进行茶树体内的再分配,具有向生长部位转移的特性,即将枯萎的老叶可以将氟向新梢和根系转移分配。不止是茶树,萝卜为保护其可食用部分不受损害也具有将过量的氟从根部转移到叶片的功能[37]。菠菜在0~800 mg·kg-1土培氟处理中,发现同样也具有在组织水平上再分配氟化物的独特机制[38],造成菠菜根部的氟含量高于地上部氟含量,这可能与根部内胚层的渗透性较低有关[39]。总而言之,茶树叶片是氟富集的主要组织,而且由于氟可能以相当部分的游离态形式存在,这种形态的氟在植物体内具有转移与再分配的特性,使得茶树叶龄也对氟的累积产生了一定的影响。
2 氟对茶树的影响及其抗性机制
氟对植物的损害主要表现在对细胞结构的破坏,如细胞膜系统、叶绿体和线粒体等,进而对光合作用、呼吸作用等生理活动及一些关键酶活性产生抑制作用等。此外,氟对植物的损害程度与其浓度及接触时间密切相关。当环境中氟浓度较低时,植株仅会出现叶片变黄、萎蔫的症状;随着氟浓度逐渐升高,植物叶片将会枯死,甚至整个植株也会死亡,并且氟处理浓度越高,植物出现伤害症状的时间就越早[40]。李丽霞等[36]发现,福鼎大白茶在低氟浓度下仍然表现出良好的长势;当氟质量浓度达到200 mg·L-1时,14 d后的茶苗开始出现叶色黯淡,少量叶片脱落的现象;而氟质量浓度升至250 mg·L-1以上时,茶苗在第12天开始出现胁迫症状并表现为叶片轻易脱落,根系逐渐转黑。植物可通过外排机制和内部耐受的方式增加自身对氟胁迫的抗性,同样的,茶树可通过外部排斥的方式防止茶树吸收过量的氟,即利用茶树细胞壁对氟进行抵御,同时,茶树体内也产生一系列反应机制增强自身的氟抗性,应对氟胁迫。
2.1 细胞壁和液泡对氟的螯合与区隔化
细胞壁作为植物抵御外界不良环境的第一层屏障,其多糖、果胶、蛋白质等组分含有的醛基、羧基、氨基和磷酸盐等基团结构及其吸附在细胞壁周围的一些金属离子等都具有螯合氟的作用[28,41-42]。氟主要分布于茶树叶片和根的细胞壁中,且90%以上的氟聚集在细胞壁的果胶和半纤维素,并且与-NH2、-COOH、-OH和金属离子分别以氢键、离子键结合的形式螯合[42]。外源氟进入细胞壁会对细胞壁离子稳态产生干扰,进而通过改变细胞壁的电荷、酶活性等对细胞壁组成结构产生影响,进而突破细胞壁的阻滞进入原生质体。Gao等[43]通过离子色谱和原子吸收光谱法检测茶树中氟的亚细胞分布,发现茶叶原生质体中98.1%的氟储存于液泡中,表明胞质中过多的氟被液泡吸收区隔化,液泡是氟累积的主要场所。因此,不仅是细胞壁对氟的螯合,液泡的区隔化作用同样可以阻止过多的氟对细胞器造成伤害,这些是茶树具有氟抗性的主要因素。
此外,添加氟可以增加果胶含量和脱甲基酯化作用,从而增加金属阳离子的吸收以及氟在细胞壁的螯合[44]。在外源施加氟后,与果胶生物合成途径相关的关键酶的基因表达、酶活性以及果胶多糖的含量增加,这表明外源施加氟促进了茶树体内果胶的生物合成,进而有助于果胶结合茶树吸收的外源氟[45]。茶多糖本身也具有吸附结合氟的能力,这与其组成、结构有关。茶多糖包含蛋白质、糖醛酸、中性糖、金属离子等多种物质。与其他植物多糖相比,茶多糖中氟含量最高、络合氟能力最强,茶叶中80%的氟与茶多糖结合[46],这种特殊络合形式也是增强植物氟抗性的因素之一。
2.2 其他离子结合
高慧敏等[41]研究表明,Mn2+、Fe3+、Al3+等离子在多糖结合氟中发挥作用。游离氟可以与Al3+、Fe3+和Ca2+等阳离子螯合形成AlFn3-n、FeFn3-n和CaF2等络合物,从而降低氟对茶树的毒害[43]。铝的施加显著增加叶片吸收和转运氟,同时缓解氟胁迫造成的茶树叶片光合效率下降,尤其是添加低浓度的铝,可以显著地提高叶片光合效率[47]。Luo等[29]发现,外源施加5 mmol·L-1Ca2+能够有效降低茶树的氟含量及丙二醛(MDA)含量,增加咖啡因、多酚和儿茶素的含量,同时还可以调节B3+、Al3+、Cu2+和Zn2+的离子稳态等,进而增强茶树的氟抗性。由此可见,茶树在氟处理后可通过其体内不同离子的变化,调节离子平衡,从而维持渗透稳态增强茶树的氟耐受性。
2.3 初级代谢产物和次生代谢产物
氟处理诱导茶叶的次要多肽,碳水化合物和氨基酸等代谢物质发生变化,为维持氟环境下茶树的稳态发挥重要的自防御机制[48]。Li等[49]研究表明,茶叶的多酚、总儿茶素和蛋白质等成分在0~0.84 mmol·L-1的氟处理下随氟浓度的增加而显著下降。而卢莉等[50]发现,武夷水仙嫩叶及老叶中茶多酚、氨基酸、咖啡碱、水浸出物等含量在氟处理下,其变化呈现低浓度促进、高浓度抑制的规律,说明氟对茶树的生理代谢具有双重作用。4 mg·L-1以下的氟质量浓度会促进一些有利于茶叶品质的氨基酸合成,改善茶叶品质,而6~16 mg·L-1的氟质量浓度则会减少氨基酸的合成并降低茶叶的品质[7],因此推测外源施加6 mg·L-1的氟质量浓度可能是茶苗对氟耐受性的阈值。此外,香气作为茶叶重要的品质特征之一,已有研究指出茶叶香气成分的含量随氟浓度的增加而降低,其中挥发性烷烃、烯烃、酯、芳香族化合物、醛、酮和酚类等化合物都呈现出先增加后减少的趋势,只有醇类呈现相反的趋势,可见高浓度氟对茶叶香气和风味品质具有不利影响[8]。
植物在逆境条件下,其细胞会迅速累积脯氨酸、有机酸、糖醇类等物质。氟胁迫下茶树游离脯氨酸的含量增加,而脯氨酸具有提高原生质渗透压,防止水分散失的作用,说明氟胁迫下积累脯氨酸是茶树应对氟胁迫的一种保护反应[51]。氟同样对茶树体内的有机酸含量产生显著影响,叶片中草酸含量随氟浓度升高呈现先升高后降低的趋势,而柠檬酸的含量不断升高[52]。还有研究发现氟处理下茶树叶片中苹果酸脱氢酶和柠檬酸合成酶的活性均显著高于根部,有利于叶片中苹果酸和柠檬酸合成并向根系运输,进而参与根系有机酸的分泌,调节茶苗对氟的适应[53]。
茶多酚(包括儿茶素)作为茶叶的次级代谢产物,具有清除自由基、提高抗氧化能力、减少脂质过氧化产物等作用,在抗逆性中发挥积极作用。儿茶素通过自身的分解代谢为木质素的合成提供基本的碳骨架,增强细胞壁,参与茶树形态建成,同时促进木质部的运输,参与茶树的抗逆性[54]。此外,还发现PAL(Phenylalanine ammonialyase),DFR(Dihydroflavanol 4-reductase)和LCR(Leucoanthocyani-din reductase)的活性与不同浓度氟处理下的多酚含量具有相似趋势,这些与生物合成相关的酶调节茶树在氟处理下茶多酚的生物合成[7],进而增加茶树的抗逆性。
此外,一些植物生长调节剂(PGRs)如赤霉素(Gibberellins,GA)、ABA、生长素、乙烯、细胞分裂素、水杨酸(Salicylic acid,SA)、油菜素类固醇(Brassinosteroids)、茉莉酸酯(Jasmonates)和多胺(Polyamines)等在植物逆境条件下和信号传导过程中发挥着重要作用。勾晓华等[55]发现在外界不良环境刺激下,乙烯含量增加将参与气孔的调节,使通过气孔进入植物的大气污染物减少,从而发挥保护植物的作用。而氟化物可通过抑制SOD的活性,导致产生过量乙烯诱导纤维素酶的合成,最终引起叶片过早脱落[56]。但针对激素类对茶树富集氟方面的研究甚少,可作为今后研究的方向之一。
2.4 抗氧化系统
高氟胁迫下茶树受到明显的氧化损伤,有研究报道称,茶树对氟耐受极限为11.84 mg·L-1,暴露于超过此浓度的生长条件下,茶树的亚细胞结构会有一定程度的破坏[57]。因此,当氟浓度升高至超过其防御能力时,茶树体内将累积过多的活性氧(ROS),导致叶绿体类囊体膜遭受伤害,进而造成细胞膜裂解、叶绿体膜结构破坏、叶绿素含量降低[57]。茶树叶片中的ROS包括超氧阴离子(O2-)和过氧化氢(H2O2)不断积累,丙二醛(MDA)的含量也在显著升高(图3),电导率不断增大[58]。与此同时,一些活性氧清除剂如过氧化物酶(POD)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)的活性在低浓度氟处理下明显增加(图3),以消除ROS,维持细胞代谢的稳定性[58]。然而,高浓度氟处理导致ROS积累量超出茶树自身所能调节的范围后,抗氧化酶的清除能力明显降低[58]。另有研究发现4 mg·L-1和12 mg·L-1氟处理时,叶片质膜透性和根系活力均低于对照组,而茶叶的CAT、POD和SOD活性增强[52],表明茶树可以通过增加酶的活性抵御氟带来的氧化损伤。
在一定浓度氟阈值下,茶树可以通过调控自身代谢清除过多的ROS,使自身免受伤害,其中抗坏血酸-谷胱甘肽(ASA-GSH)循环在茶树响应氟胁迫时也起到了重要作用[59]。在低浓度的氟胁迫下,茶苗可通过ASA-GSH循环及体内的抗氧化系统,在一定程度上提高抗氧化酶活性以清除氟胁迫产生的过量活性氧自由基,以减轻ROS对膜的伤害[60]。在ASA-GSH循环中一些主要的酶,如谷胱甘肽还原酶(GR)、抗坏血酸过氧化物酶(APX)和脱氢抗坏血酸还原酶(DHAR)的活性,随氟处理浓度的升高表现出先升高后降低的趋势(图3)[58]。因此,这也说明了在低浓度的氟胁迫下,茶树可以通过提高AsA-GSH循环中的酶性活增强对不良环境的抵抗能力。
2.5 有机物代谢
低浓度氟处理下,茶树叶片的叶绿素含量和光合速率略微升高[61],而过量的F-容易与叶绿素中的Mg2+结合或固定[57],导致叶绿素的合成受抑制,进而抑制植物的生长。此外,高浓度氟破坏叶片的细胞超微结构[57],削弱茶树叶片的光合电子传递能力,进而影响茶树的光合效率(图4)。此外,氟化物还可以通过抑制叶绿体类囊体膜上的ATP合酶活性从而阻碍光合磷酸化[62],并通过抑制核酮糖-1,5-二磷酸羧化酶(Rubisco)的活性从而阻碍碳同化过程[63]。碳水化合物的代谢包括光合作用合成有机物及有机物分解释放能量[63],其代谢所需能量主要依赖于糖类,而高氟茶树品种可以补偿在高氟环境下光能消耗而导致的离子不平衡,增加糖基转移酶和水解酶的表达水平,使更多的淀粉和纤维素被重新分配给葡萄糖,为茶树的耐受过程提供更多能量[64]。
氟化物还通过抑制谷酰胺合成酶、谷氨酸脱氢酶和丙氨酸转氨酶的活性,导致植物的固氨能力下降,减少蛋白质合成所需的氨基酸,抑制蛋白质的合成,加速蛋白质的分解,破坏植物的氮代谢[65]。Yu等[66]研究发现,高浓度氟处理能明显抑制与呼吸作用相关的酶活性,并导致茶树叶片线粒体空泡化,甚至降解,严重时会对线粒体造成不可逆的损害,进而导致酶分子附着的表面减少,减弱细胞呼吸,茶树有氧糖代谢受阻,从而降低呼吸作用。也有研究发现,在氟处理初期,植物的呼吸方式由糖酵解途径变为磷酸戊糖途径,致使呼吸作用增强[67]。Miller等[67]观察到,从氟处理初期植物组织中分离出的线粒体,其呼吸速率和ATPase活性明显增强,而在受氟抑制的组织发现呼吸速率和ATPase活性减弱。但目前,有关通过呼吸作用增强植物氟抗性的机制研究甚少,仍有待深入探讨。
2.6 氟在茶树体内的转移
茶树体内存在着独特的转移模式,可以使茶树积累大量的氟后依然保持正常的生理状况。这种转移模式主要表现为茶树根系不断将吸收的氟运输至茶树体内(图1)。当叶片的氟累积到一定程度时,氟会向其他器官转移,但不会影响茶树对氟的吸收[68]。此外,李丽霞等[36]发现老叶中的氟可以向根系和新梢转移,但老叶中的氟含量仍高于其他部位。Niu等[69]通过正电子发射断层成像(Positron-emitting tracer,PET)发现F-被根吸收后约1.5 h到达茶树地上部,根系对F-有滞留作用,且根部吸收的F-主要通过木质部向上运输,但部分通过茎中的韧皮部向上运输。针对茶树叶片吸收的F-,发现F-首先沿茎部向下输送,然后再向上输送,并在顶部和同侧叶片积累,但未在根部观察到[69]。可见,叶片和根系吸收转运氟的方式各有不同。
2.7 基因表达
信号转导通路在转录因子的调控中发挥重要作用,响应特定的应激信号。Li等[19]研究发现,氟处理诱导类受体蛋白激酶(Receptor-like kinases,RLK)基因的表达,进而激活Ca2+ATPase,活化的Ca2+ATPase促进氟化物的吸收和运输(图2)。氟信号同样激活[70]、[71]等转录因子的基因表达,它们在茶树的氟累积及氟耐受性中发挥作用。氟也可以通过激活ABA、GA和多胺等相关的合成基因[72],进而增加植物的氟抗性。
外源施加阴离子通道抑制剂显著抑制了茶树对氟的吸收,可见通过阴离子通道是茶树吸收、迁移F-的主要方式[31]。近年来的研究发现,许多单细胞生物以及绿色植物可以通过细胞膜的F-输出蛋白保持细胞质较低的F-浓度,从而使植物的氟胁迫降到最低。现在已知两个系统发育独立的F-输出家族[20]:CLCF型F-/H+反向转运蛋白,以及在原核生物中称作“crcB(Fluc)”、在真核生物中称为FEX(Fluoride exporter)的小膜蛋白家族。其中,电压门控氯离子通道(Voltage-gated chloride channel,CLC)不仅以2Cl-/1H+运输Cl-,还可以通过NO3-、F-、Br-、I-等其他卤族元素阴离子[73]。而F-/H+反向转运蛋白CLCF在细菌中具有相比Cl-优先转运F-的功能[74],且茶树的CLC基因对F-的响应比Cl-更敏感[21]。另外一类是分布更为广泛的氟输出蛋白家族crcB(Fluc),目前的研究表明大肠杆菌及酿酒酵母中存在氟输出蛋白crcB(Fluc),并且具有外排细胞F-的功能[20]。近来也发现crcB(Fluc)的同源基因茶树FEX可以通过降低组织内的氟含量增强对氟化物的耐受性[22],且FEX膜转运蛋白发挥的氟转运作用是植物氟防御的重要机制[74]。具有典型的ABC转运蛋白结构域的CsCL667蛋白和氟离子转运蛋白CsFEX属于同一基因家族,在氟处理下表达量显著上调,ABC转运蛋白含量也受氟诱导显著增加,使ATP水解释放能量,促使细胞累积大量的F-[23]。进一步通过大肠杆菌异源表达验证CsCL667具有一定耐氟性以及外排氟离子的功能[23]。由此可见,探索相关的氟输出蛋白有利于茶树氟富集特性的研究,这将是未来分子研究的新方向。

图3 氟对茶树抗氧化系统的影响
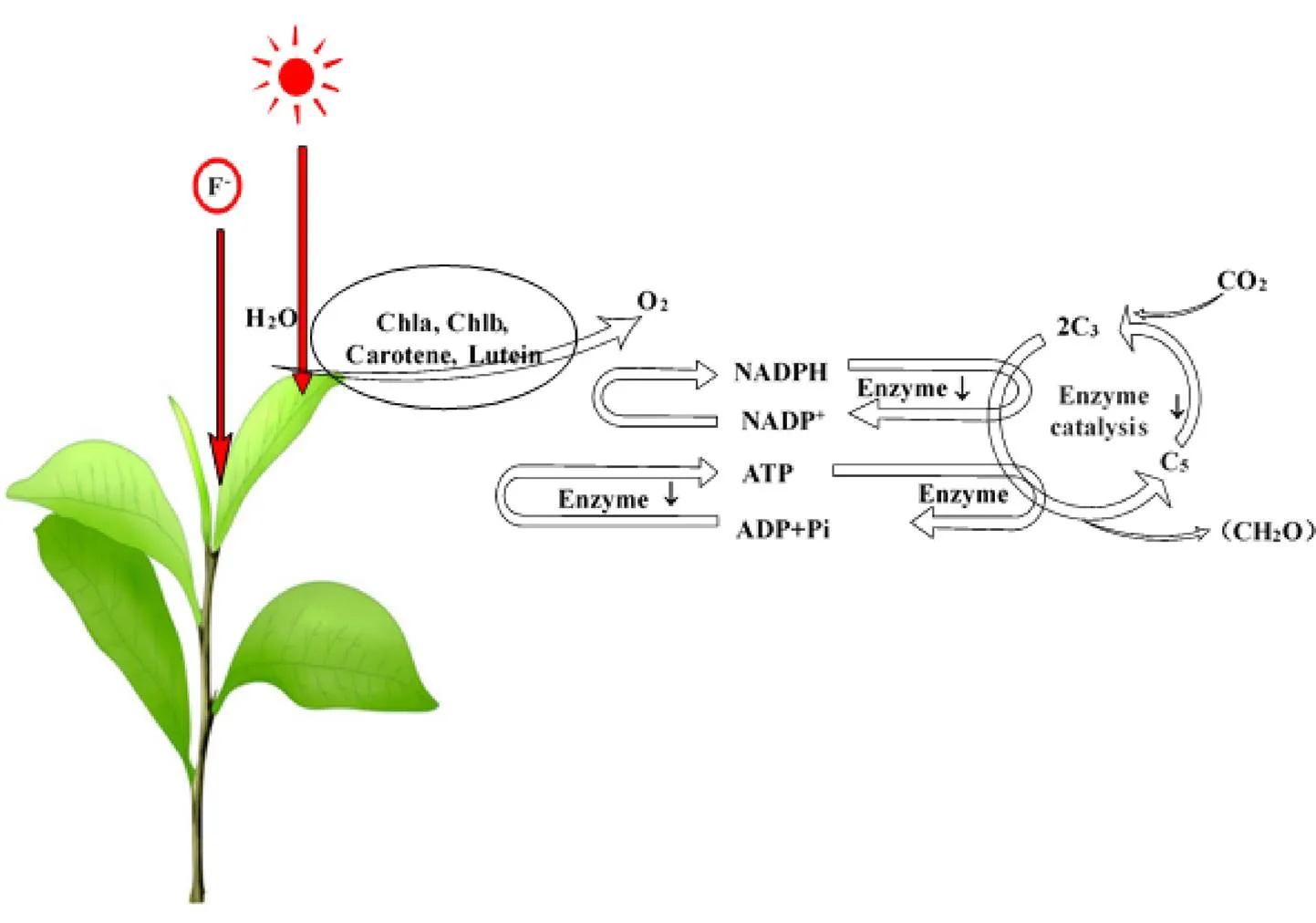
图4 氟对茶树光合作用的影响
3 降氟措施
茶树虽然具有聚氟耐氟的特性,但过多的氟累积仍会危害茶树,影响茶叶产量和品质。研究者针对茶树品种的差异、茶园土壤氟含量的降低措施、加工技术的改良等方面,探索提出了降低茶叶氟含量的方法。
3.1 选育低氟茶树品种
不同茶树品种对氟的累积量具有很大的差异性,选择合适的低氟品种是首要的降氟措施。前期对乌牛早、浙农138等31个茶树品种进行氟含量的研究,发现不同茶树品种成熟叶片的氟含量具有差异性[75]。石元值等[76]的研究也表明,不同品种茶树对氟的吸收存在差异,并发现中茶108与中茶102、龙井43、乌牛早相比,具有稳定的低氟吸收富集特点,认为这与茶树的基因型相关。不同茶树品种对氟的富集能力不同由遗传差异所决定,且有研究者根据氟富集程度的不同将55个茶树品种分为了高、中、低3种[77]。一般来讲,品种间氟含量的差异与叶片的结构相关。云南大叶种的叶片大而薄,海绵组织发达,细胞排列疏松,细胞间隙大,有利于叶片进行气体交换,相比之下,凤凰水仙这种小乔木型大叶种的叶片小且厚,栅栏组织发达,细胞排列紧密,不利于茶树吸收空气中的氟[77]。可见,培育、选用低氟品种的茶树,是减少茶叶氟含量的根本举措。
3.2 改善茶园土壤管理措施
施肥也是改变茶树氟含量的一种措施,一般通过根部施肥或者叶面喷肥的方式进行茶园管理。其中,对叶面喷施适当浓度的氮肥可以降低茶树新梢中的氟含量,例如在夏季进行叶面喷施含量2%以下的尿素,可以显著降低茶树新梢中的氟含量,但当喷施的尿素含量高于2%时,便会提高茶树新梢中的氟含量[78]。通过根部施氮肥同样可以降低茶叶新梢的氟含量,但仅在茶叶新梢氟含量较高的情况下施用有效[78]。
氟对茶叶的品质有很大的影响,含氟量高的茶叶,其没食子儿茶素含量比较低,而适量的铝元素可以提高儿茶素的含量[79]。施加适量的铝可以提高茶叶中氨基酸、咖啡碱、可溶性糖以及茶多酚的含量,能够促进茶叶品质的提升,但铝施用量超过2 g·m-2时,茶叶咖啡碱与茶多酚的含量增多,造成茶汤发苦发涩;所以,合理控制茶叶中的氟、铝含量,能够促进茶叶品质的提升。
钙通过与氟形成CaF2,通过改变土壤有效性氟的含量以降低茶树对氟的吸收。张永利等[80]研究发现,施用不同浓度的Ca(NO3)2、CaCl2、Ca(OH)2会影响酸性土壤pH的变化,以pH5.0为拐点,pH的降低或升高均会使土壤水溶态氟含量增加。可见,钙不止是通过形成CaF2对土壤有效氟含量产生影响,还可以改变土壤颗粒表面电荷、离子交换能力、络合物/配合物的稳定性进而调整土壤pH[81],降低土壤交换态氟含量。因而,在酸性土壤中施用碱性的Ca(OH)2等,降低土壤中氟的有效性,可以减少茶树对氟的吸收富集。
研究发现添加木炭和竹炭能显著提高土壤中Fe/Mn氧化物结合态氟的浓度,而水溶性氟和交换性氟含量显著降低,这有效减少了土壤氟的可利用性,影响土壤氟的有效性,进而降低茶树对氟的吸收与积累[82]。腐植酸铝(HAA)吸附剂能通过与可溶性F-螯合的方式,使得土壤溶液中的F-含量显著降低,最终降低茶叶对氟化物的吸收[83]。目前,已经研发出茶园土壤降氟剂B,具有降低土壤氟10%~40%的效果,但鲜叶的氟含量降低不明显,有待进一步提升研发产品[78]。
3.3 改进茶叶加工工艺
茶叶的加工工艺会对其含氟量造成一定的影响。通过水潦青、炒青、蒸青3种不同的杀青方式比较青砖茶氟含量的差异,发现水潦青、炒青后的茶叶氟含量显著低于蒸青茶叶的氟含量[84]。揉捻也是影响氟含量的工艺,砖茶原料在揉捻后细胞破碎,用室温水洗1~2 min不仅可以有效降低茶叶氟含量,而且能最大限度地保留其有效成分[85]。并且发现在砖茶渥堆发酵工序前,翻拌的同时喷洒微生物制剂[86],可以有效地降低砖茶氟含量,提高砖茶品质、香气和滋味。李兰英等[87]针对传统砖茶的加工工艺,探索出一种低氟优质的砖茶生产工艺,即鲜叶→摊放→杀青→揉捻→复炒→复揉→晒→渥堆→蒸→渥堆→炒干→蒸/压砖→砖茶。此外,采用拼配技术也是一种有效降低茯砖茶氟含量的加工方法,可以在选择原料时进行氟含量的测定,筛分不同氟含量的茶鲜叶,对高氟含量的原料、中氟含量的原料和低氟含量的原料进行拼配[88],最后达到控制茯砖茶氟含量的目的。
4 展望
近来,关于茶树氟富集特性的研究颇丰,同时对茶树的氟抗性机制也有了一定的研究进展,为茶叶降氟措施的研究提供了一定的理论基础,但相关的研究仍然存在一定的问题,有待进一步探索。
(1)近年来,关于茶树根系吸收、转运氟方面的研究较多,也在茶树中发现了氟离子转运通道蛋白——CLC和FEX,并对其转运氟的功能进行了初步验证,但其分子结构、互作机制、调控机制等分子生物学机理仍不清楚。未来,在鉴定更多的氟离子转运通道蛋白、验证其功能、解析其分子机制等方面的研究有待进一步的深入和挖掘,以期解析茶树富集氟的机制。
(2)茶树不仅具有聚氟特性,还兼具富铝、喜酸、偏铵吸收等特征。已有研究联合氟与不同pH环境或外源施加铝后茶树的氟累积特性,综合分析茶树的氟富集特性,但该方面的研究仅进行了初步的探索,深入研究可能发掘茶树自身耐氟的独特机制。同时,茶树具有富含茶多酚、生物碱和茶氨酸等次生代谢物质的特性,已有研究表明茶树富集过量的氟能影响这些次生代谢物质的含量,但茶中大量的次生代谢物质在茶树富集氟、耐受氟中的作用及作用机制还鲜有报道,值得深入研究以发掘茶树富集氟的特有生物学机制。
(3)在茶叶降氟方面,通过对比不同茶树品种氟含量,筛选出了氟累积相对较低的品种,但传统茶树低氟品种的选育周期长,难度大,现代分子育种技术在低氟品种选育方面至今仍然是空白。通过茶园土壤管理,对茶园土壤中交换态氟含量降低具有一定效果,但对茶树叶片降氟效果相对较弱。总之,茶叶降氟措施研究中,在品种选育、土壤管理、茶叶加工工艺改良等方面都取得了一定的进展,但目前为止,仍然没有系统、高效、完善的降氟管理措施。
综述所述,在未来的研究探索中,需加快解析茶树富集氟的分子生物学机制,加强建设低氟品种、低氟茶园管理模式、低氟茶叶加工工艺等相结合的完善降氟技术体系。
[1] 庞廷祥. 大气氟污染对作物的危害及防治措施[J]. 热带农业工程, 2000(1): 3-6, 22.
Pang T X. Harm of atmospheric fluorine pollution to crops and prevention measures [J]. Tropical Agricultural Engineering, 2000(1): 3-6, 22.
[2] 沙济琴, 郑达贤. 茶树黄棪对氟的生物积累特征[J]. 福建茶叶, 1993(3): 25-28.
Sha J Q, Zheng D X. The bioaccumulation characteristics of fluoride in‘Huangyan’ [J]. Tea in Fujian, 1993(3): 25-28.
[3] 王琼琼, 薛志慧, 陈志丹, 等. 不同茶树种质间氟铝元素积累特性的研究[J].热带作物学报, 2016, 37(5): 862-869.
Wang Q Q, Xue Z H, Chen Z D, et al. Accumulation and distribution of fluoride/aluminum elements in different tea cultivars [J]. Chinese Journal of Tropical Crops, 2016, 37(5): 862-869.
[4] Shu W S, Zhang Z Q, Lan C Y, et al. Fluoride and aluminium concentrations of tea plants and tea products from Sichuan Province, PR China [J]. Chemosphere, 2003, 52(9): 1475-1482.
[5] 郜红建, 刘腾腾, 张显晨, 等. 安徽茶园土壤氟在茶树体内的富集与转运特征[J]. 环境化学, 2011, 30(8): 1462-1467.
Gao H J, Liu T T, Zhang X C, et al. Bioaccumulation and translocation of fluoride from soils to different parts of tea plants in Anhui province [J]. Environmental Chemistry, 2011, 30(8): 1462-1467.
[6] Shahab S, Mustafa G, Khan I, et al. Effects of fluoride ion toxicity on animals, plants, and soil health: a review [J]. Fluoride, 2017, 50(4): 393-408.
[7] Ni D J, Li C L. Effect of fluoride on the amino acid composition of tea leaves [J]. Research Report Fluoride, 2016, 49: 274-278.
[8] Yang X, Yu Z, Zhang B, et al. Effect of fluoride on the biosynthesis of catechins in tea [(L.) O. Kuntze] leaves [J]. Scientia Horticulturae, 2015, 184: 78-84.
[9] 高绪评, 王萍. 饮茶摄氟量的探讨[J].植物资源与环境, 1998 (3): 54-58.
Gao X P, Wang P. Researches of the uptaken fluorine amount from drinking tea [J]. Journal of Plant Resources and Environment, 1998 (3): 54-58.
[10] Baunthiyal M, Ranghar S. Physiological and biochemical responses of plants under fluoride stress: an overview [J]. Fluoride 2014, 47(4): 287-293.
[11] 王玉梅, 柴如山, 郜红建. 茶树根系跨膜主动吸收氟的表观特征[J]. 农业环境科学学报, 2016, 35(8): 1473-1479.
Wang Y M, Chai R S, Gao H J. Apparent characteristics of active transmembrane uptake of fluoride by tea plant roots [J]. Journal of Agro-Environment Science, 2016, 35(8): 1473-1479.
[12] 张磊, 阮建云. 茶树氟吸收动力学参数测定方法的研究[J]. 茶叶科学, 2008(3): 195-200.
Zhang L, Ruan J Y. Comparison on research methods for fluoride uptake kinetics of tea plant [J]. Journal of Tea Science, 2008(3): 195-200.
[13] 张磊. 茶树氟吸收动力学特性的研究[D]. 杭州: 中国农业科学院, 2008.
Zhang L. Research for fluoride uptake kinetics characteristic of tea plant [D]. Hangzhou: Chinese Academy of Agricultural Sciences, 2008.
[14] 彭传燚, 陈静, 蔡荟梅, 等. 茶树对氟的吸收动力学特性研究[J]. 热带作物学报, 2013, 34(3): 495-500.
Peng C Y, Chen J, Cai H M, et al. The kinetic characteristics of solution fluoride uptake by tea plant [J]. Chinese Journal of Tropical Crops, 2013, 34(3): 495-500.
[15] 秦樊鑫, 吴迪, 黄先飞, 等. 高氟病区茶园土壤氟形态及其分布特征[J]. 中国环境科学, 2014, 34(11): 2859-2865.
Qin F X, Wu D, Huang X F, et al. Distribution characteristics and speciation of fluorine in tea Garden soils in the high fluoride area [J]. China Environmental Science, 2014, 34(11): 2859-2865.
[16] 李张伟. 粤东凤凰山茶区土壤氟化学形态特征及其影响因素[J]. 环境化学, 2011, 30(8): 1468-1473.
Li Z W. Chemical forms of fluoride in soils forms 12 tea gardens of Fenghuang mountaing, East of Guangdong Province [J]. Environmental Chemistry, 2011, 30(8): 1462-1467.
[17] 谢忠雷, 陈卓, 孙文田, 等. 不同茶园茶叶氟含量及土壤氟的形态分布[J]. 吉林大学学报(地球科学版), 2008, 38(2): 293-298.
Xie Z L, Chen Z, Sun W T, et al. Content of fluoride in tea leaves and distribution of fluoride in soils from different tea gardens [J]. Journal of Jilin University (Earth Science Edition), 2008, 38(2): 293-298.
[18] 王玉梅. 茶树根系跨膜吸收氟的微观机制和转录组学特征[D]. 合肥: 安徽农业大学, 2017.
Wang Y M. Microscopic mechanisms transcriptome characteristics of transmembrane absorption of fluoride by tea plant roots [D]. Hefei: Anhui Agricultural University, 2017.
[19] Li Q S, Lin X M, Qiao R Y, et al. Effect of fluoride treatment on gene expression in tea plant () [J]. Scientific Reports, 2017, 7: 9847.
[20] Mcilwain B C, Ruprecht M T, Stockbridge R B. Membrane exporters of fluoride ion [J]. Annual Review of Biochemistry, 2021, 90 (1): 559-579.
[21] Xing A Q, Ma Y C, Wu Z C, et al. Genome-wide identification and expression analysis of the CLC superfamily genes in tea plants () [J]. Functional Integrative Genomics, 2020, 20(4): 497-508.
[22] Zhu J J, Xing A Q, Wu Z C, et al. CsFEX, a fluoride export protein gene from, alleviates fluoride toxicity in transgenicand[J]. Journal of Agricultural Food Chemistry, 2019, 67: 5997-6006.
[23] 广敏. ABC转运蛋白介导茶树根系跨膜吸收转运氟的分子机制研究[D]. 合肥: 安徽农业大学, 2020.
Guang M. Molecular mechanisims of ABC transporter mediated transmembrane absorption and transport of of fluoride by tea plant roots [D]. Hefei: Anhui Agricultural University, 2020.
[24] Ruan J Y, Ma L F, Shi Y Z, et al. The impact of pH and calcium on the uptake of fluoride by tea plants (L.) [J]. Annals of Botany, 2004, 93(1): 97-105.
[25] Yang Y, Liu Y, Huang C F, et al. Aluminium alleviates fluoride toxicity in tea () [J]. Plant Soil, 2016, 402(1): 179-190.
[26] Morita A, Horie H, Fujii Y, et al. Chemical forms of aluminum in xylem sap of tea plants (L.) [J]. Phytochemistry, 2004, 65(20): 2775-2780.
[27] Nagata T, Hayatsu M. Aluminium kinetics in the tea plant using27Al and19F NMR [J]. Phytochemistry, 1993, 32(4): 771-775.
[28] Cai H M, Peng C Y, Chen J, et al. X-ray photoelectron spectroscopy surface analysis of fluoride stress in tea ((L.) O. Kuntze) leaves [J]. Journal of Fluorine Chemistry, 2014, 158: 11-15.
[29] Luo J L, Ni D J, He C, et al. Influence of exogenous calcium on the physiological, biochemical, phytochemical and ionic homeostasis of tea plants ((L.) O. Kuntze) subjected to fluorine stress [J]. Plant Growth Regulation, 2019, 87: 455-465.
[30] Zhang X C, Gao H J, Yang T Y, et al. Anion channel inhibitor NNPB-inhibited fluoride accumulation in tea plant () is related to the regulation of Ca2+, CaM and depolarization of plasma membrane potential [J]. International Journal of Molecular Sciences, 2016, 1(17): 57. doi: org/10.3390/ijms17010057.
[31] Zhang X C, Gao J H, Zhang Z Z, et al. Influences of different ion channel inhibitors on the absorption of fluoride in tea plants (L.) [J]. Plant Growth Regulation, 2013, 69(1): 99-106.
[32] Zhang X C, Gao H J, Yang T Y, et al. Al3+-promoted fluoride accumulation in tea plants () was inhibited by an anion channel inhibitor DIDS [J]. Journal of the Science of Food Agriculture, 2016, 96(12): 4224-4230.
[33] Zhang L, Li Q, Ma L F, et al. Characterization of fluoride uptake by roots of tea plants ((L.) O. Kuntze) [J]. Plant Soil, 2013, 366(1): 659-669.
[34] Pottosin I, Velarde-Buendía A M, Bose J, et al. Polyamines cause plasma membrane depolarization, activate Ca2+-, and modulate H+-ATPase pump activity in pea roots [J]. Journal of Experimental Botany, 2014, 65(9): 2463-2472.
[35] 唐茜, 赵先明, 杜晓, 等. 氟对茶树生长,叶片生理生化指标与茶叶品质的影响[J]. 植物营养与肥料学报, 2011, 17(1): 186-194.
Tang Q, Zhao X M, Du X, et al. Effects of fluorine stress on growth, physiological-biochemical characteristics and quality of tea leaves [J]. Plant Nutrition and Fertilizer Science, 2011, 17(1): 186-194.
[36] 李丽霞, 杜晓, 何春雷. 水培茶苗对氟的吸收累积特性[J]. 四川农业大学学报, 2008, 26(1): 62-66, 78.
Li L X, Du X, He C L. Absorption and accumulation characteristics of fluorine in nutrient liquid cultured tea plant [J]. Journal of Sichuan Agricultural University, 2008, 26(1): 62-66, 78.
[37] Chen Y Z, Wang S L, Nan Z R, et al. Effect of fluoride and cadmium stress on the uptake and translocation of fluoride and cadmium and other mineral nutrition elements in radish in single element or co-taminated sierozem [J]. J Environmental Experimental Botany, 2017, 134: 54-61.
[38] Jha S K, Nayak A K, Sharma Y K. Response of spinach () to the added fluoride in an alkaline soil [J]. Food and Chemical Toxicology, 2008, 46(9): 2968-2971.
[39] Takmaz-Nisaneiouglu S, Davison A W. Effects of aluminium on fluoride uptake by plants [J]. New Phytologist, 1988, 109: 149-155.
[40] 周丽丽, 高必达, 宋奎. 不同植物对KF的剂量反应及其伤害阈值和致死浓度[J]. 中国农学通报, 2015, 31(14): 164-170.
Zhou L L, Gao B D, Song K. Dose-response of different plants on potassium fluoride and acute injury thresholds and lethal concentrations [J]. Chinese Agricultural Science Bulletin, 2015, 31(14): 164-170.
[41] 高慧敏. 茶多糖对氟离子的吸附特性研究[D]. 武汉: 华中农业大学, 2019.
Gao H M. Study on the absorption characteristics of tea polysaccharides on fluoride [D]. Wuhan: Huazhong Agricultural University, 2019.
[42] 刘思怡, 朱晓静, 房峰祥, 等. 茶树叶片氟亚细胞分布及其与细胞壁结合特性的研究[J]. 茶叶科学, 2018, 38(3): 305-312.
Liu S Y, Zhu X J, Fang F X, et al. Fluorine subcellular distribution and its combining characteristics with cell wall in tea leaves () [J]. Journal of Tea Science, 2018, 38(3): 305-312.
[43] Gao H J, Zhao Q, Zhang X C, et al. Localization of fluoride and aluminum in subcellular fractions of tea leaves and roots [J]. Journal of Agricultural and Food Chemistry, 2014, 62(10): 2313-2319.
[44] Luo J L, Ni D J, Li C L, et al. The relationship between fluoride accumulation in tea plant and changes in leaf cell wall structure and composition under different fluoride conditions [J]. Environmental Pollution, 2021, 270: 116283.
[45] Luo J L, Hu K, Qu F F, et al. Metabolomics analysis reveals major differential metabolites and metabolic alterations in tea plant leaves (L.) under different fluorine conditions [J]. Journal of Plant Growth Regulation, 2020(3): 1-13.
[46] 春晓亚. 氟在茶树新梢的分布特性及与多糖的结合方式初探[D]. 武汉: 华中农业大学, 2011.
Chun X Y. Study on distribution of fluorine in tea shoots and the way of combination with polysaccharides [D]. Wuhan: Huazhong Agricultural University, 2011.
[47] 钟秋生, 林郑和, 郝志龙, 等. 氟铝互作对茶树叶片叶绿素荧光特性的影响[J]. 茶叶科学, 2019, 39(5): 537-546.
Zhong Q S, Lin Z H, Hao Z L, et al. Effect of fluoride and aluminum interaction on the chlorophyll fluorescence characteristics of tea leaves [J]. Journal of Tea Science, 2019, 39(5): 537-546.
[48] Peng C Y, Xu X F, Zhu H Y, et al. Metabolics and ionomics responses of tea leaves ((L.) O. Kuntze) to fluoride stress [J]. Plant Physiology and Biochemistry, 2021 158: 65-75.
[49] Li C L, Ni D J. Effect of fluoride on chemical constituents of tea leaves [J]. Fluoride, 2009, 42(3): 195-202.
[50] 卢莉, 刘金仙, 程曦, 等. 氟铝交互处理对茶叶主要化学成分的影响[J]. 热带作物学报, 2017, 38(10): 1956-1962.
Lu L, Liu J X, Cheng X, et al. Effect of Al and F interaction on the main chemical components in tea leaves [J]. Chinese Journal of Tropical Crops, 2017, 38(10): 1956-1962.
[51] 王小平, 刘鹏, 罗虹, 等. 铝氟交互处理对茶树生理特性的影响[J]. 园艺学报, 2009, 36(9): 1359-1364.
Wang X P, Liu P, Luo H, et al. Effect of Al and F interaction on physiological characteristics of tea plant [J]. Acta Horticulture Sinica, 2009, 36(9): 1359-1364.
[52] 马士成. 铝对茶树氟吸收、累积、分布特性的影响及其机理研究[D].杭州: 浙江大学, 2012.
Ma S C. Effect of aluminum on uptake, distribution and accumulation of fluorine in tea plants and its mechanism [D]. Hangzhou: Zhejiang University, 2012.
[53] 王丽霞. 茶树对氟的富集及其生理响应机制研究[D]. 杨凌: 西北农林科技大学, 2014.
Wang L X. Fluoride accumulation in tea plant and its physiological response mechanism [D]. Yangling: Northwest A&F University, 2014.
[54] 杨贤强, 王岳飞, 陈留记, 等. 茶多酚化学[M]. 上海: 上海科学技术出版社, 2003.
Yang X Q, Wang Y F, Chen L J, et al. Tea polyphenol chemistry [M]. Shanghai: Shanghai Science and Technology Press, 2003.
[55] 勾晓华, 王勋陵, 陈发虎. 氟化氢熏气和喷施防护剂对小麦应激乙烯产生的影响[J]. 应用与环境生物学报, 2000, 6(2): 117-120.
Gou X H, Wang X L, Chen F H. Effect of HF fumigation and spraying protective agents on stress ethylene production in wheat [J]. Chinses Journal Applied and Environmental Biology, 2000, 6(2): 117-120.
[56] 孟范平, 李桂芳, 吴方正. 氟害大豆超氧化物歧化酶活性与叶绿素含量及叶片脱落的关系[J]. 生态与农村环境学报, 2002, 18(2): 34-38.
Meng F P, Li G F, Wu F Z. Relationship between SOD activity and chlorophyll content and abnormal defoliation of soybean () leaves exposed to airborne fluoride [J]. Rural Eco-environment, 2002, 18(2): 34-38.
[57] Li C, Zheng Y, Zhou J, et al. Changes of leaf antioxidant system, photosynthesis and ultrastructure in tea plant under the stress of fluorine [J]. Biologia Plantarum, 2011, 55(3): 563-566.
[58] 董阳阳. 氟处理对茶树叶片光合系统、活性氧代谢及超微结构的影响[D]. 合肥: 安徽农业大学, 2015.
Dong Y Y. Effect of F supply on photosynthesis, reactive oxygen metabolism and ultra-structure of tea plant () leaves [D]. Hefei: Anhui Agricultural University, 2015.
[59] 李春雷. 氟对茶树抗坏血酸-谷胱甘肽循环系统的影响[J]. 江苏农业学报, 2016, 32(5): 1018-1022.
Li C L. ASA-GSH cycle in tea plant exposed to fluoride application [J]. Jiangsu Journal of Agricultural Science, 2016, 32(5): 1018-1022.
[60] 黄鑫, 宋晓维, 陈玉琼. 茶树吸收富集氟的机制研究进展[J]. 茶叶科学, 2016, 36(6): 551-556.
Huang X, Song X W, Chen Y Q. Advances in fluorine absorption and accumulation mechanisms in tea plant [J]. Journal of Tea Science, 2016, 36(6): 551-556.
[61] 李春雷. 氟对茶树幼苗生理生化的影响及其作用机制研究[D]. 武汉: 华中农业大学, 2011.
Li C L. Study on the Effect and mechanism of fluoride in the physiology and biochemistry of tea seedlings [D]. Wuhan: Huazhong Agricultural University, 2011.
[62] 孟范平, 吴方正. HF对梅树超氧化物歧化酶和纤维素酶活性的影响[J]. 生态学杂志, 1997, 16(5): 28-31.
Meng F C, Wu F Z. Effect of HF on the activities of superoxide dismutase and cellolase in plum () leaves [J]. Chinese Journal of Ecology, 1997, 16(5): 28-31.
[63] 徐丽珊. 大气氟化物对植物影响的研究进展[J]. 浙江师范大学学报, 2004, 27(1): 66-71.
Xu L S. Effects of atmospheric fluoride pollution on plant [J]. Journal of Zhejiang Normal University (Natural Science Edition), 2004, 27(1): 66-71.
[64] Yang P D, Liu Z, Zhao Y, et al. Comparative study of vegetative and reproductive growth of different tea varieties response to different fluoride concentrations stress [J]. Plant Physiology and Biochemistry, 2020, 154: 419-428.
[65] 申秀英, 吴方正. 氟化物对桑叶氮代谢某些影响的研究[J]. 农业环境科学学报, 1991, 10(5): 194-197, 211.
Shen X Y, Wu F Z. Effect of fluoride on the metabolism of nitrogen in the leaves of mulberry [J]. Agro-environmental Protection, 1991, 10(5): 194-197, 211.
[66] Yu M H, Miller G W. Effect of fluoride on the respiration of leaves from higher plants [J]. Plant Cell Physiology 1967(3): 483-493.
[67] Miller J E, Miller G W. Effects of fluoride on mitochondrial activity in higher plants [J]. Physiologia Plantarum, 2010, 32(2): 115-121.
[68] 杨晓, 张月华, 余志, 等. 氟对茶树生理的影响及茶树耐氟机制研究进展[J]. 华中农业大学学报, 2015, 34(3): 142-146.
Yang X, Zhang Y H, Yu Z, et al. Physiological effects of fluoride on tea plant and fluoride-resistant mechanism of tea [J]. Journal of Huazhong Agricultural University, 2015, 34(3): 142-146.
[69] Niu H L, Peng C Y, Zhu X D, et al. Positron-emitting tracer imaging of fluoride transport and distribution in tea plant [J]. Journal of the Science of Food and Agriculture, 2020, 100(8): 3554-3559.
[70] Wang Y H, Chang P P, Pan J T, et al. Effect of aluminium and fluoride on R2R3-MYB transcription factor characterization and expression in[J]. Biologia Plantarum, 2019, 63(1): 298-307.
[71] Pan J T, Chang P P, Ye X L, et al. Transcriptome-wide analysis of MADS-box family genes involved in aluminum and fluoride assimilation in[J]. Plant Biotechnology, 2018, 35(4): 313-324.
[72] Banerjee A, Roychoudhury A. Melatonin application reduces fluoride uptake and toxicity in rice seedlings by altering abscisic acid, gibberellin, auxin and antioxidant homeostasis [J]. Plant Physiology and Biochemistry, 2019, 145: 164-173.
[73] Stockbridge R B, Lim H H, Otten R, et al. Fluoride resistance and transport by riboswitch-controlled CLC antiporters [J]. Proceedings of the National Academy of Sciences of the United States of America Elife, 2012, 109(38): 15289-15294.
[74] Tausta S L, Berbasova T, Peverelli M, et al. The fluoride transporter Fluoride Exporter (FEX) is the major mechanism of tolerance to fluoride toxicity in plants [J]. Plant Physiology, 2021, 186(2): 1143-1158.
[75] 陈瑞鸿, 梁月荣, 陆建良, 等. 茶树对氟富集作用的研究[J]. 茶叶, 2002, 28(4): 187-190.
Chen R H, Liang Y R, Lu J L. Studies on fluorine enrichment in tea plant () [J]. Journal of Tea, 2002, 28(4): 187-190.
[76] 石元值, 王新超, 方丽, 等. 四个茶树品种的氟吸收累积特性比较研究[J]. 植物营养与肥料学报, 2013, 19(2): 396-403.
Shi Y Z, Wang X C, Fang L, et al. Characteristics of fluorine absorption and accumulation of four different tea tree varieties [J]. Plant Nutrition and Fertilizer Science, 2013, 19(2): 396-403.
[77] 黎南华. 不同生态环境的茶叶氟含量浅析[J]. 福建茶叶, 1994(2): 21-23.
Li N H. Analysis on the fluoride content of tea in different ecological environments [J]. Tea in Fujian, 1994(2): 21-23.
[78] 包小村. 茶叶最新降氟科技成果[J]. 湖南农业, 2020, 510(6): 17.
Bao X C. The latest scientific and technological achievements of tea defluoridation [J]. Hunan Agriculture, 2020, 510(6): 17.
[79] 于静怡, 王军, 于晓峰. 基于计算机技术下氟铝交互化学处理对茶叶化学成分的影响分析[J]. 福建茶叶, 2018, 40(4): 10.
Yu J Y, Wang J, Yu X F. Analysis of the influence of fluorine-aluminum interactive chemical treatment on the chemical composition of tea based on computer technology [J]. Tea in Fujian, 2018, 40(4): 10.
[80] 张永利, 廖万有, 王烨军, 等. 添加含钙化合物对茶园土壤pH及有效氟的影响[J]. 中国农学通报, 2013, 29(1): 132-137.
Zhang Y L, Liao W Y, Wang Y J, et al. Influence of addition of calcium compounds on pH and available fluoride content in tea garden soil [J]. Chinese Agricultural Science Bulletin, 2013, 29(1):132-137.
[81] 王凌霞, 胡红青, 闵艳林, 等. 茶园土壤水溶性氟含量的模拟调控[J]. 环境科学学报, 2011, 31(7): 1517-1525.
Wang L X, Hu H Q, Min Y L, et al. Simulated control of water-soluble fluoride content in tea garden soils [J]. Acta Scientiae Circumstantiae, 2011, 31(7): 1517-1525.
[82] Gao H J, Zhang Z Z, Wan X C. Influences of charcoal and bamboo charcoal amendment on soil-fluoride fractions and bioaccumulation of fluoride in tea plants [J]. Environmental Geochemistry and Health, 2012, 34(5): 551-562.
[83] Huang C Y, Zhang H, Zeng W H, et al. Enhanced fluoride adsorption of aluminum humate and its resistance on fluoride accumulation in tea leaves [J]. Environmental Technology, 2020,41(3): 329-338.
[84] 陈玉琼, 倪德江, 春晓娅, 等. 不同杀青方式对青砖茶原料氟含量的影响[J]. 湖北农业科学, 2011, 50(6): 1193-1195.
Chen Y Q, Ni D J, Chun X Y, et al. Effects of different fixation ways on the fluoride contents of Qingzhuan tea material [J]. Hubei Agricultural Sciences, 2011, 50(6): 1193-1195.
[85] 春晓亚, 陈玉琼, 倪德江, 等. 水洗对砖茶揉捻叶氟含量及主要品质成分的影响[J]. 湖北农业科学, 2011, 50(12): 2453-2455.
Chun X Y, Chen Y Q, Ni D J, et al. Effects of rolling tea leaves with washing water on fluorine and quality components [J]. Hubei Agricultural Sciences, 2011, 50(12): 2453-2455.
[86] 李国林, 梅树华, 齐桂年. 采用微生物制剂处理的低氟砖茶生产工艺: CN101427716[P]. 2009-05-13.
Li G L, Mei S H, Qi G N. Production process of low-fluoride brick tea treated with microbial preparation: CN101427716 [P]. 2009-05-13.
[87] 李兰英, 王云, 尧渝, 等. 低氟砖茶加工工艺及品质研究[J]. 中国农学通报, 2017, 33(15):133-138.
Li L Y, Wang Y, Rao Y, et al. Processing technology and quality of low-fluoride brick-tea [J]. Chinese Agricultural Science Bulletin, 2017, 33(15): 133-138.
[88] 纪晓明, 李三原, 周兴长. 以拼配技术降低茯砖茶氟含量的生产方法: CN101731379A[P]. 2010-06-16.
Ji X M, Li S Y, Zhou X Z. Production method for reducing fluoride content of Fuzhuan tea by blending technology: CN101731379A [P]. 2010-06-16.
Research Advances of Fluoride Accumulation Mechanisms in Tea Plants ()
XING Anqi1, WU Zichen1, XU Xiaohan1, SUN Yi1, WANG Genmei2, WANG Yuhua1*
1. College of Horticulture, Nanjing Agricultural University, Nanjing 210095, China;2. Co-innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China
(L.) O. Kuntze is a hyper fluoride (F) accumulation plant, whose F content in tea leaves is much higher than other plants, without any toxic symptoms. However, F is not an essential element for tea plant growth, and under high F stress, F affects the normal growth of plants by destroying the cell structure and inhibiting enzyme activities. In order to provide a theoretical basis for the future study of F accumulation in tea plants, the research progresses in the absorption, enrichment and accumulation/detoxification mechanisms of F in tea plants were reviewed.
, fluoride, accumulation mechanism
S571.1
A
1000-369X(2022)03-301-15
2021-09-01
2021-11-03
国家自然科学基金(31972458)、江苏现代农业(茶叶)产业技术体系(JATS[2021]457)
邢安琪,博士研究生,主要从事茶树育种与栽培研究。*通信作者:wangyuhua@njau.edu.cn
(责任编辑:赵锋)
