乡村小流域不同土壤景观表土有机质团聚体分布与分子组成变化
2022-06-17刘晓雨龙国刚陈硕桐林清美陈敬龙包旭华卞荣军郑聚锋李恋卿潘根兴
刘 纯,赵 正,刘晓雨,龙国刚,陈硕桐,林清美,陈敬龙,包旭华,卞荣军,郑聚锋,李恋卿,潘根兴*
(1 南京农业大学农业资源与生态环境研究所,江苏南京 210095;2 南京农业大学资源环境科学学院土壤学系,江苏南京210095;3 江苏省南京市溧水区晶桥镇农业技术推广站,江苏南京 211224)
土壤有机质(SOM)是地球最大的陆地碳库,是全球碳循环的重要组成部分[1]。全球1 m深土壤中有机碳储量约为1550 Pg,分别是大气和陆地植被碳库的2和3倍[2]。土壤有机质通过保持和缓冲土壤养分和水分,保障微生物生长及其多样性[3-4],在维持土壤肥力和作物生产力等生态系统功能中发挥着关键作用。土壤有机质积累及其组分结构随土地利用和人为管理的变化可能影响土壤质量和生态系统功能的小尺度空间变异。
关于土壤有机质的本质认识是近20多年来土壤学发展的重要领域[5]。一方面,土壤有机质被认为是一系列来源、结构和组成不同的生物源有机化合物的集合体,其在土壤中表现出不同的化学稳定性[6];另一方面,土壤有机质是与土壤矿物质特别是土壤团聚体呈不同的结合和保护状态的连续分布体系,其在土壤中的积累和稳定受土壤过程和生态系统过程所控制[7-8]。实际上,土壤有机质是来源于植物组织的不同程度降解产物和微生物的残体或代谢产物在土壤中的持留[5,7],前者多保存于微团聚体和宏团聚体中,而后者多紧密结合于微团聚体和粉黏粒中。近年来发展的连续提取—GC/MS鉴定法可以用来探析陆地生态系统中土壤有机质的生命来源与降解程度[9-12]。例如,脂类和木质素衍生的酚类是土壤有机质常见组分,其相对丰度可以指示有机质的来源和降解程度[13-15]。而非破坏性土壤结构和水稳性团聚体的分离及配合探针的有机质原位鉴定可以用来揭示有机组分的土壤结合和保护特征[8]。因此,土壤有机质的形态(水溶性有机质、颗粒态有机质等)、团聚体分布(轻组有机质和重组有机质等)及脂类和木质素等生物标志物丰度的分析,可以探讨土壤有机质的动态变化[16]。例如,东北黑土区从草地、农田和裸地土壤有机质含量的差异反映在颗粒态有机质和宏团聚体分数的差异。农田恢复为林地和草地后,有机质含量的快速恢复与残体生物量输入密切相关[17]。由于土地利用和人为活动的影响,土壤有机质在量和形态上变化的同时,基于生物标志物的土壤有机碳分子组成和基于团聚体分组的有机质保护状态可能改变。分析一定区域内土壤有机质含量、团聚体分布和化学组成变化,可以反映土地利用和人为管理对土壤有机质的改变。
土壤景观是长期尺度的地貌形成与相对短期尺度的人为土地利用构成的地表系统,是地理景观与生态系统相互作用的地表形态[18-19],其形成、分布、性质及分类属于土壤地理学(或者地理土壤学,pedology)的主要研究范畴[20]。了解和模拟生态系统过程及功能的土壤景观小尺度空间变化是当前面向全球变化和可持续发展管理的核心科学问题。作为地理学、生态学和地貌学与土壤学的交叉研究,研究土壤景观生态行为,特别是联系植被覆盖和土壤侵蚀的土壤景观模拟研究在最近20年来得到长足发展。这些研究中,除了表土厚度、土壤质地和水分外,土壤有机碳积累和储存的分异一直是小流域内土壤景观分异及模拟的核心内容[18-19,21-22]。有关土壤景观有机质变化的研究尚未深入关注与土壤有机质储存有关的化学组成及其团聚体分布的空间变异。从几万m2到数千万m2的丘陵小流域土壤和生态系统性质的空间差异往往被认为是土壤空间异质性(变异性),而实际是植被覆盖的多样性本质。了解土壤有机质的土壤景观分异对于丘陵地区乡村多样性农业产业布局和乡村生态环境可持续发展具有重要意义。
本研究选取江苏省南京市溧水区某行政村小流域内4种土壤景观,采集非破坏表土样品分析土壤有机碳含量及其团聚体分布,进而采用连续提取法提取土壤有机分子,分析生物质标志物分子组成和丰度差异,探讨地理景观基础上人为土地利用影响下土壤有机碳的量与团聚体分配及化学结构变化的关系,为阐明乡村地区耕作利用方式下农田土壤有机质变化动态提供新视角。
1 材料与方法
1.1 研究区概况
研究区位于江苏省南京市溧水区晶桥镇芝山行政村村域范围,属于宁镇扬(南京、镇江、扬州的简称)山地丘陵区,海拔220 m以上的芳山为境内最高峰(图1),母岩主要为砂岩、泥岩,局部有石灰岩。依山势西南山麓依次表现丘陵缓坡和坡地间谷地。该地区秦汉以来就有人类栖居,目前芳山山地为森林保护区,仍保留有亚热带常绿混交林,山麓坡地除保留集体林地外,多开垦为果树蔬菜园地或玉米、杂粮旱地,土壤类型为黄棕壤,而谷地长期耕垦为稻田,多为潴育型水稻土,土壤深厚。形成了山地森林土壤景观、坡地园地-耕地土壤景观和谷地稻田土壤景观,在村域小流域内镶嵌分布。该区域属亚热带季风气候,年平均气温16.4℃,年平均降雨量1147.0 mm,每年6月下旬到7月上旬为梅雨季节,年平均日照1969.0 h,无霜期224天。
1.2 土样采集与处理
于2020年11月进行采样。分别在芳山西坡山地、芳山西南山麓缓坡地和西麓谷地选择林地景观、园地景观、旱地景观和稻田景观样地,每个土壤景观类型设3个重复样地(林地的样地间距在100 m以上,农地采用邻近的不同田块)。采样前,通过访问农民确定土壤没有经受过冲刷、填埋或平整。采样时,去除地面凋落物,采用土芯采样器采集0—20 cm表土,每个样地梅花形随机五点采样形成一个混合样品,混合均匀后密封于容积为1 L的不锈钢样品罐中。样品当天带回实验室,四分法分为3份,一份过2 mm筛后-20℃保存用于土壤微生物分析,一份-4℃冷藏用于团聚体分离,一份经自然风干后磨碎过筛用于理化性质分析和土壤有机质分析。采样点的基本信息见表1。
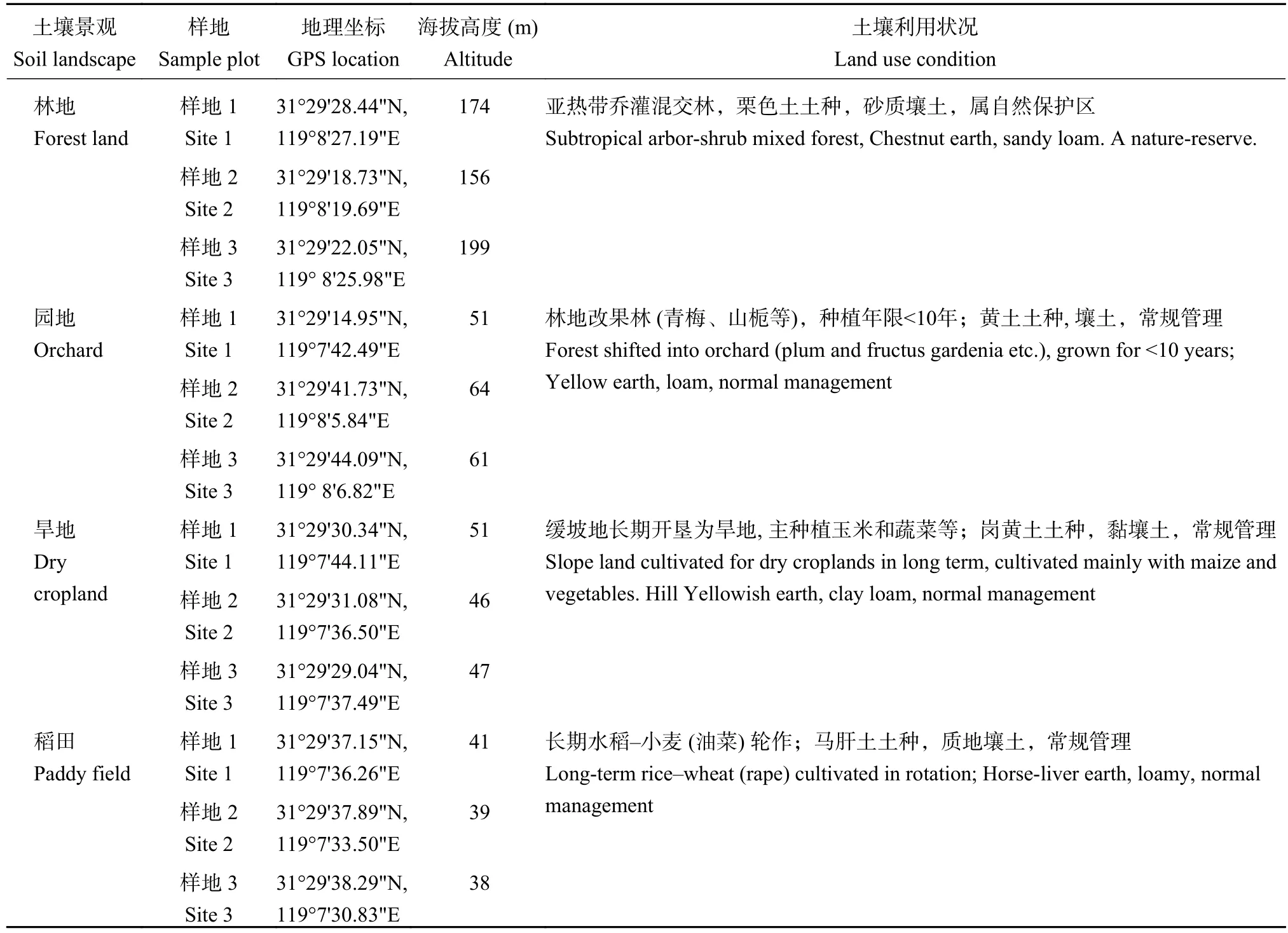
表1 芳山小流域采样点分布Table 1 Locations of the sampling sites within the Fangshan watershed
1.3 分析方法
1.3.1 土壤基本理化性质 土壤样品处理和基本理化性质分析依照鲁如坤[23]的方法进行。土壤容重(BD)和含水量采用不锈钢环刀法野外采样测定;土壤pH采用pH计测定(水土比2.5∶1);土壤可溶性有机碳(DOC)采用Jena碳氮分析仪测定(水土比为5∶1);土壤有机碳采用重铬酸钾容量法—外加热法测定;土壤微生物生物量碳采用氯仿熏蒸—K2SO4提取法测定;土壤全氮采用凯氏定氮法测定;土壤有效磷采用0.03 mol/L NH4F-0.025 mol/L HCl法提取和钼锑抗比色法测定。水稳性团聚体分离采用Cambardella等[24]提出并经过Six等[25]改进的湿筛法,依次分离得到如下团聚体粒径组:宏团聚体(MAC,macroaggregate, 2000~250 μm)、微团聚体 (MIC,microaggregate, 250~53 μm)和粉黏粒 (S-C, silt &clay, <53 μm)。
1.3.2 有机质连续提取 依照Otto等[10]的方法,土壤样品依次进行总溶剂(TSE)提取,碱水解(BHY)提取和氧化铜氧化(CUO)提取,分别主要得到游离脂、结合态脂和木质素酚,采用气相色谱-质谱联用仪(GC-MS)测定生物标志物有机分子丰度。具体方法和步骤如下:
游离态脂类提取:取3 g过0.15 mm筛的土壤样品,超声-超纯水处理,去除极性分子;依次用甲醇溶液,二氯甲烷/甲醇混合液(1∶1, v/v)和二氯甲烷溶液超声提取并离心,合并3次提取的溶液,氮吹后的剩余物质即为游离态脂类。
结合态脂类提取:上步提取后残余样品(约3 g)置入浓度为1 mol/L的甲醇化KOH溶液中,循环加热回流反应3 h。待冷却后离心并收集上清液,加入稀盐酸将上清液pH调为1.0,旋蒸去除溶液中有机溶剂。加入乙酸乙酯和超纯水萃取,将乙酸乙酯相转移至玻璃管中,氮吹后容器底部残留微黄色粉末即为结合态脂类。
木质素酚类提取:碱水解提取后土样转置于圆底烧瓶,加入1 g氧化铜和100 mg六水合硫酸亚铁铵及15 mL NaOH。氮气封闭条件下加热反应3 h。自然冷却后离心并收集上清液。稀盐酸调整提取液pH为1.0,室温下黑箱条件静置1 h (防止肉桂基分解)。加入乙酸乙酯萃取,氮吹后容器底部残留物即为木质素酚类。
1.3.3 提取物的衍生化 采用1 mL二氯甲烷:使用甲醇(1∶1, v/v)对产物复溶,并通过0.22 μm有机滤膜进行过滤。将溶液转移到1.5 mL GC小瓶中,分别加入内标溶液。每种混合溶液在溶剂蒸发系统(Genevac®EZ-2 plus)中去除有机溶剂,与100 μL N,O-双(三甲基硅)三氟乙酰胺(BSTFA)和吡啶(体积比9∶1)在70°C下反应3 h,得到三甲基硅基(TMS)衍生物。冷却后,加900 μL正己烷(HPLC)稀释至1 mL后GC/MS检测。
1.3.4 GC-MS分析条件 衍生化的提取物在气相色谱仪(GC, Agilent 7890B, USA)耦合单四极杆质谱(MSD, Agilent 5977B, USA)进行分析。分析条件为:初始温度65℃ 保持2 min,增温梯度6℃/min增温到290℃,保持20 min。使用Agilent G4513A 自动进样器,进样量为1 μL,分流比为1∶10。载气为氦气,电离源为70eV的EI源,扫描范围为50~650 Da。将所得质谱图与 Agilent Masshunter NIST 17.0 数据库进行匹配,鉴定生物标志物有机分子,根据峰高计算该分子丰度,并采用样品有机质含量换算其相对丰度,其单位表示为 mg/g, SOC。
1.4 数据处理
土壤团聚体平均重量直径(mean weight diameter,MWD)使用下式计算:
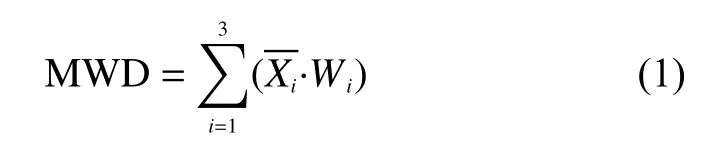
有机碳的团聚体分配(g/kg)=团聚体中有机碳含量(g/kg)×团聚体的质量分数(%) (2)
连续提取—GC/MS分析得到的生物标志物分子检测结果表示为相对丰度(mg/g, SOC),并依据Shannon多样性指数法计算土壤有机质分子多样性:
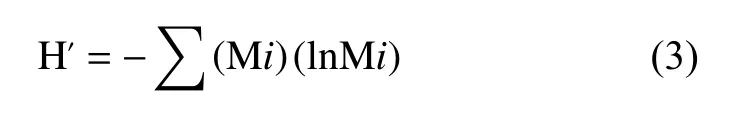
式中,Mi为分子i在分子总丰度中的分数。
数据均以3个重复样品的平均值±标准差表示。数据处理采用Excel 2019和IBM SPSS Statistics 20.0软件。绘图采用Origin 2021。采用单因素方差分析(One way ANOVA)和Duncan检验来检验景观间差异,P<0.05为显著。
2 结果与分析
2.1 土壤理化性状
表2表明,旱地和园地的土壤容重(1.5 g/cm3左右)大于林地和稻田(1.3 g/cm3左右)。山地和坡地为黄棕壤,pH均低于5.0,属于酸性土。稻田因长期水耕熟化,pH提高到5.7。土壤矿质颗粒组成从自然林地到园地、旱地和稻田,砂粒含量降低而粉砂含量提高,黏粒含量也有增高,稻田增加幅度最小。相应地,林地土壤质地为沙壤土,园地和旱地为粘壤土,而稻田则为壤土。土壤阳离子交换量以林地最高,为9.92 cmol/kg,园地(6.09 cmol/kg)和旱地(6.62 cmol/kg)显著降低,而稻田 (8.00 cmol/kg)的降低幅度未达显著水平。林地表土的有效磷含量低于10 mg/kg,而由于施肥,园地、旱地和稻田的有效磷含量高达78 mg/kg以上。不同景观土壤团聚体的平均重量直径差异显著,说明不同景观土壤团聚体的稳定性明显不同,其中林地最高,显著高于园地和旱地,后二者又显著高于稻田。
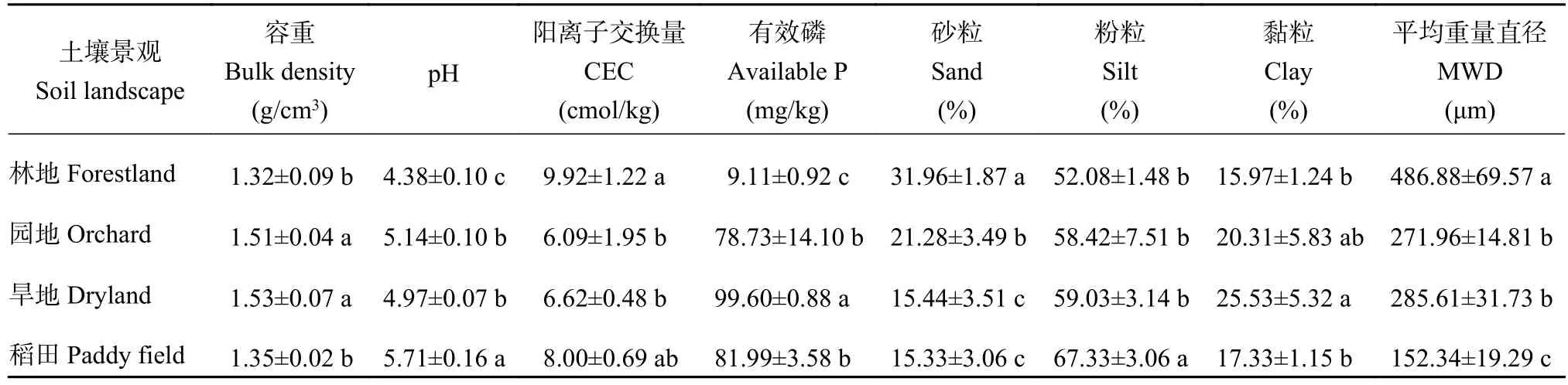
表2 供试小流域不同土壤景观表土(0—20 cm)基本理化性质Table 2 Basic physiochemical properties of topsoil under different landscapes in the watershed
2.2 土壤有机碳含量及形态
不同景观表层土壤总有机碳含量、溶解性有机碳含量及微生物量碳含量均存在较大差异(表3)。与林地相比,园地、旱地和稻田表土本体有机碳含量分别降低70%、57%和51%。土壤全氮含量林地也显著高于其他3个土壤景观,而其他3个土壤景观全氮含量没有显著差异。溶解性有机碳含量和微生物量碳含量林地和稻田接近,均高于园地和旱地。以MBC/SOC计算土壤微生物熵,林地中为1.8%,低于园地(2.7%)、旱地(2.1%)及稻田(3.1%)。
2.3 土壤有机碳的团聚体分布
图2显示不同景观下土壤中三个水稳性团聚体粒径组的比例(质量百分数)。林地中的宏团聚体和微团聚体比例分别为38%和35%,略高于非团聚性的粉黏粒(27%)。与林地相比,园地中的宏团聚体和微团聚体比例分别降低了19和10个百分点,旱地中分别降低了17和15个百分点。与自然状态的林地相比,三种农业景观的土壤结构发生了巨大的变化。土壤团聚体的平均重量直径在林地为486.88 μm,园地和旱地分别为271.96和285.61 μm,稻田最低 (仅 152.34 μm)。
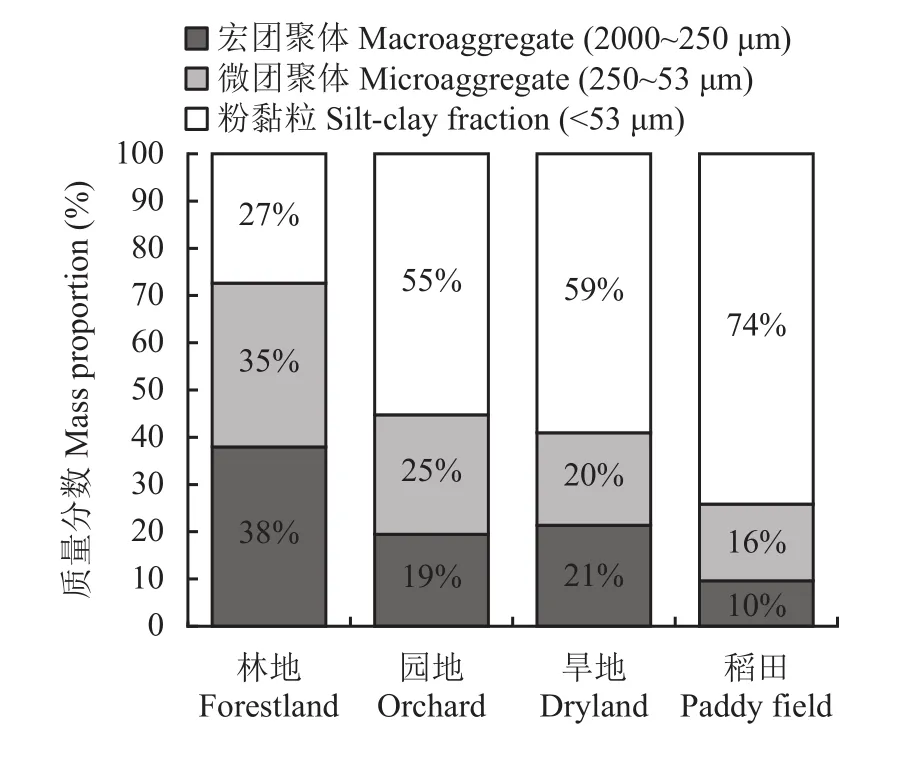
图2 不同土壤景观表土水稳性团聚体粒径组质量分数Fig. 2 Mass proportion of size fractions of water-stable aggregates of topsoil under different landscapes
图3显示,不同土壤景观下不同粒径水稳性团聚体有机碳含量及有机碳分配情况。以团聚体粒径组质量分数及团聚体粒径组有机碳含量而计算出的有机碳团聚体分配,它可以直观的指明宏团聚体、微团聚体、粉黏粒组分对于土壤总有机碳的贡献。首先,有机碳含量总体为宏团聚体高于微团聚体和粉黏粒,仅林地中有机碳含量的团聚体间差异不明显。不同景观间,与林地相比,园地、旱地及稻田宏团聚体结合有机碳含量分别降低了85%、81%和71%,微团聚体结合有机碳分别降低了74%、79%和67%,粉黏粒结合有机碳分别降低了48%、18%和3%。其中,宏团聚体和微团聚体的有机碳含量的变化趋势与土壤总有机碳的变化趋势吻合;而粉黏粒有机碳含量在林地显著高于园地、旱地和稻田中,但后三者间无显著差异。其次,就有机碳的团聚体分配来说,林地中宏团聚体占有40%,微团聚体占33%和粉黏粒占27%。相比之下,林地中宏团聚体、微团聚体及粉黏粒组间分配相对均衡,园地、旱地和稻田中有机碳倾向于粉黏粒(图3右)。园地、旱地和稻田三种农业景观土壤中,宏团聚体有机碳占比分别为23%、27%和18%,微团聚体有机碳占比分别为23%、20%和24%,而粉黏粒有机碳占比分别为54%、53%和58%。与林地相比,微团聚体和宏团聚体结合态有机碳(近似于土壤颗粒态有机碳)减少的幅度在园地、旱地及稻田中分别达到84%、73%和74%。计算团聚体结合态有机碳占土壤有机碳总量的质量分数,林地为73%,有机碳以团聚体结合态为主;而园地、旱地和稻田中分别降低到46%、47%和42%,低于未团聚粉黏粒有机碳(为矿物紧密结合态有机碳)所占的质量分数,在园地、旱地和稻田土壤中有机碳以粉黏粒矿物紧密结合态为主。在本研究中,表土有机碳总量与黏粒和粉粒含量没有显著的相关性,但与团聚体结合态有机碳含量呈显著(P<0.05)的线性关系,与DOC/MBC值有显著(P<0.05)的负相关关系(图4),提示团聚体分配和微生物活动是控制土壤有机碳水平的主导因素。
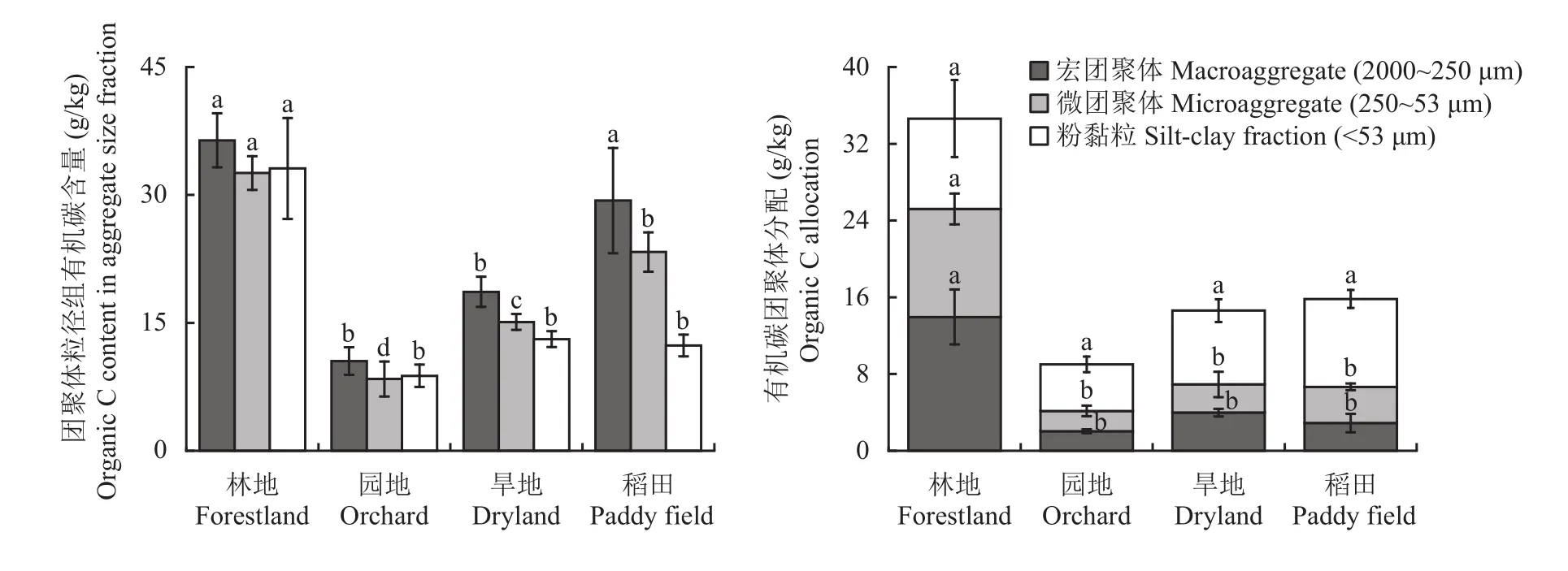
图3 供试不同土壤景观下表土水稳性团聚体粒径组有机碳含量(左)及其有机碳分配(右)Fig. 3 Organic carbon content (left) and the occupation (right) of water-stable soil aggregate size fractions of topsoil under different landscapes
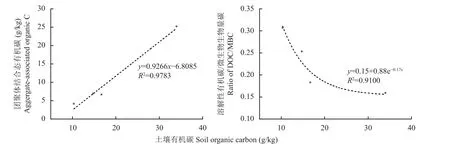
图4 表土总有机碳含量与团聚体结合态有机碳(左)和可溶性有机碳(DOC)与微生物量碳(MBC)比值(右)的关系Fig. 4 Correlation of soil organic C with aggregate-associated C (Left) and the ratio of dissolved organic C/microbial biomass C (DOC/MBC) (Right) under different soil landscapes
2.4 可提取态有机质的丰度及其分子多样性
经3级连续提取得到的不同有机质组分中生物标志物分子数目、总丰度及多样性结果列于表4。所提取有机质的生物标志物分子的相对丰度均在15 mg/g SOC以下,基本上以结合态脂类丰度高于木质素酚类,木质素酚类丰度又高于游离态脂类丰度。生物标志物分子多样性除了林地游离态脂类丰度较高外,与分子丰度表现出类似的规律。另外,总分子丰度与分子多样性间存在一定的相关关系(图5)。
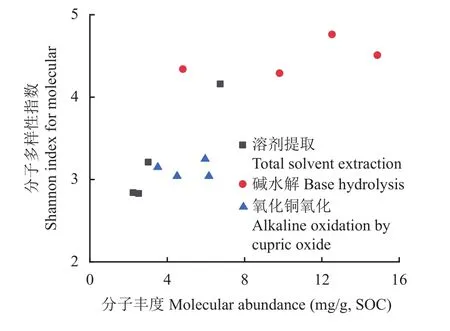
图5 提取有机质组分中生物标志物分子丰度与分子多样性的关系Fig. 5 Shannon index of diversity in relation to abundance of biomarker molecules of soil organic matter extractions
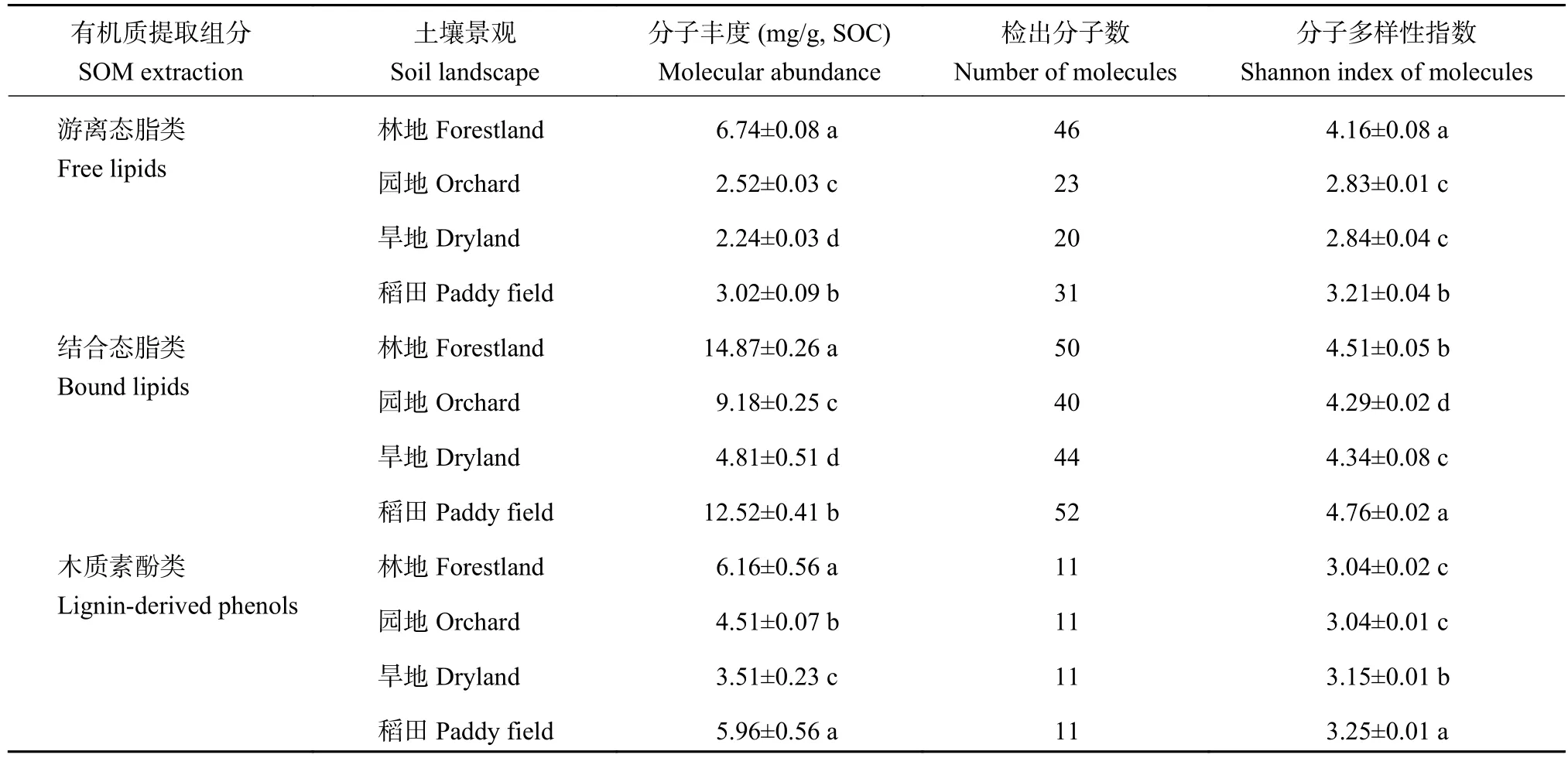
表4 供试不同土壤景观表土提取态有机质生物标志物分子丰度及分子多样性指数(H')Table 4 Biomarker abundance and Shannon index of extracted topsoil soil organic matter (SOM) in different soil landscapes
游离态脂类、结合态脂类和木质素酚类的生物标志物分子总丰度均表现为林地>稻田>旱地>园地。与林地相比,园地、旱地和稻田的游离态脂类相对丰度分别降低了63%、67%和55%,结合态脂类丰度分别降低了38%、68%和16%,木质素酚类丰度分别降低了27%、43%和3%。游离态脂类和结合态脂类的总丰度和全部分子的多样性均与土壤有机碳含量存在显著(P<0.05)的正相关关系(图6)。不过,稻田和旱地木质素酚类分子多样性指数显著高于林地和园地。
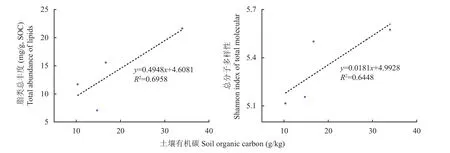
图6 表土提取态脂类分子总丰度(左)和总分子多样性(右)与土壤有机碳含量的关系Fig. 6 Total abundance of lipids (left) and Shannon index of total molecules (right) correlated to organic carbon content
2.5 提取态有机质分子组成
游离脂肪酸是主要的脂类分子(图7),林地中其丰度达1.14 mg/g SOC,而稻田为0.3 mg/g SOC,园地和旱地仅为0.1 mg/g SOC,远低于林地。游离脂肪酸碳链以C14~C18为主,以C16脂肪酸丰度最高,且偶数碳链脂肪酸丰度高于奇数碳链(图8A)。林地的游离脂类分子中,不仅检出了萜类,还检出碳链长度在C18~C30的多个烷醇分子,而园地和旱地仅检出C18烷醇,稻田中仅检出碳链短于C22的烷醇(图7)。这些也进一步证实了林地相较于园地、旱地和稻田的较高的游离态脂类分子多样性。游离态脂类中烷烃组分在园地和旱地明显富集,占游离态脂类的65%以上。所有检测出的烷烃均集中在C20~C27碳链,且C22碳链的分子丰度最高(图8B)。
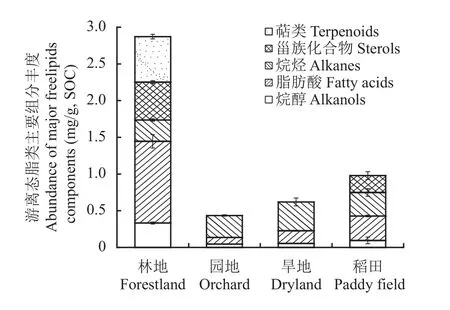
图7 不同土壤景观表土游离态脂类主要生物标志物分子丰度Fig. 7 Abundance of major components of biomarkers in free lipids under different landscapes
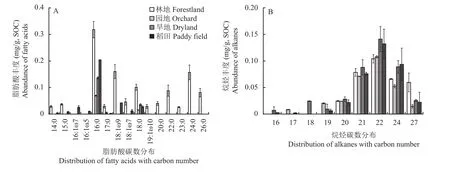
图8 不同土壤景观表土游离态脂类中脂肪酸(A)和烷烃(B)组分的丰度Fig. 8 Fatty acids (A) and alkanes (B) fraction in total solvent extraction components under different landscapes
图9显示碱水解提取的结合态脂类生物标志物分子丰度分布,其中主要包括羟基酸、烷酸及苯甲酸,也检出了丰度较低的烷醇和二羧酸。烷酸系列包括正构烷酸和支链烷酸,其丰度在2.0~6.2 mg/g SOC。正构烷酸由C9~C26的饱和及非不饱和同系物组成。与上述游离脂肪酸相比,偶数链烷酸含量均高于奇数链烷酸。支链烷烃酸主要分布在C13~C19,且丰度较低。羟基酸中特定生物标志物分子可代表植物软木脂及角质,其丰度在林地高达9.3 mg/g SOC,在园地和稻田在2.5~3.0 mg/g SOC,旱地降低至1.1 mg/g SOC。根据Otto和Simpson[10]定义的参数计算来源于软木脂和角质的脂类丰度,稻田中软木脂及角质等生物标志物丰度明显高于林地、园地和旱地等土壤景观中。
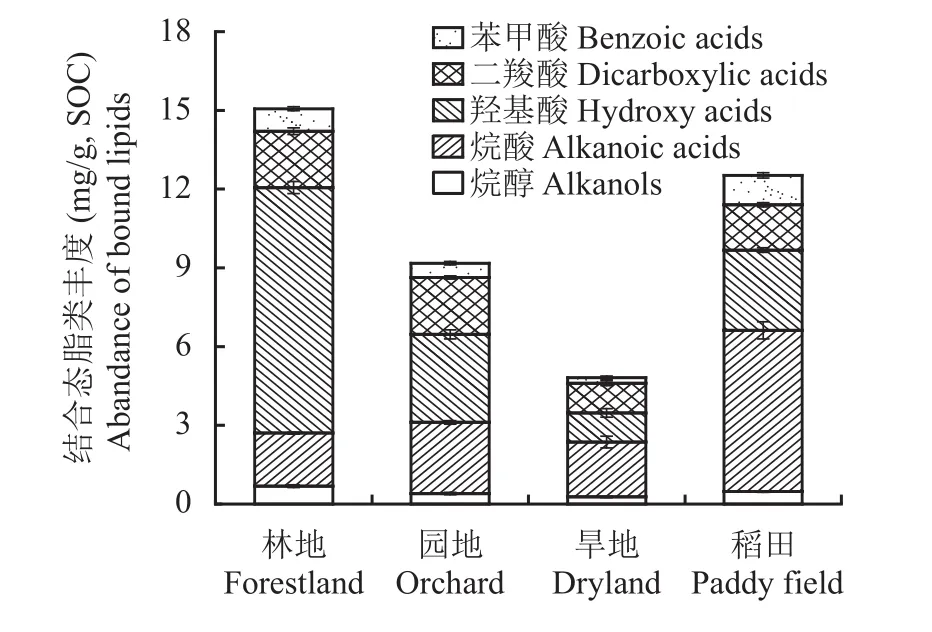
图9 不同土壤景观表土碱水解有机质中结合态脂类的丰度Fig. 9 Abundance of bound lipids in base hydrolyzed soil organic matter of topsoil under different landscapes
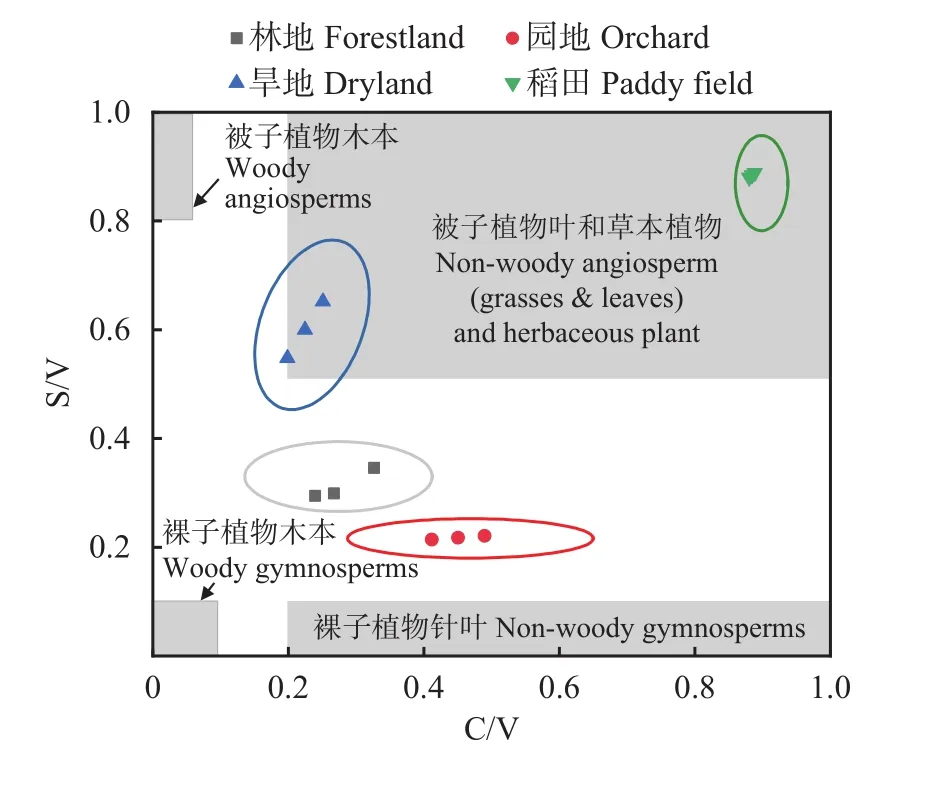
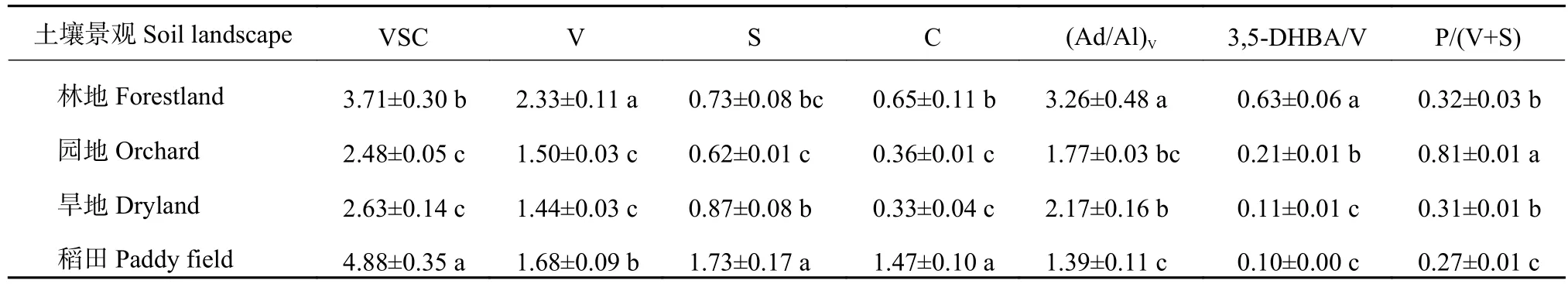
如表5所示,碱提取有机质中长链烷酸(>C20)与短链烷酸( 表5 不同土壤景观表土有机质中不同来源与降解的结合态脂解析Table 5 Sources and degradation indices of bound lipids of topsoil organic matter under different landscapes 氧化铜氧化提取得到有机质中苯甲酸及木质素衍生酚,检出的木质素单体包括香兰基(V)、丁香基(S)和肉桂基(C)等酚类组分(表6)。稻田木质素单体总丰度达4.88 mg/g SOC,显著高于林地(3.71 mg/g SOC)、园地(2.48 mg/g SOC)和旱地(2.63 mg/g SOC)。这主要由于稻田中丁香基和肉桂基单体丰度的显著增加。木质素单体的S/V值在0.31~1.03,C/V值在0.23~0.87,两者在不同景观间的变化趋势大致相同。木质素酚的比值(S/V和C/V)可以指示陆地植被对土壤有机质的相对贡献,而C/V用来区分木本和非木本的输入,S/V用来区分裸子植物和被子植物的输入。如图10所示,稻田和旱地中木质素可以代表作物来源(图中指示为被子植物落叶或草本植物),而林地和园地木质素来源较复杂多样。与前述所有分子丰度的趋势不同,(Ad/Al)V、3,5-DHBA/V、P/(V+S)等比值指示的木质素降解程度,稻田明显低于林地、园地和旱地,显示稻田中木质素降解程度较低,可能是稻田淹水环境木质素难以分解而相对持留。 图10 不同土壤景观表土有机质中木质素酚的植被来源Fig. 10 Vegetation source indicators of lignin phenols of topsoil soil organic matter under different landscapes 表6 不同土壤景观表土有机质中氧化铜氧化提取的木质素衍生酚类的丰度(mg/g, SOC)Table 6 Abundance of lignin-derived phenols in CUO extraction of topsoil organic matter in different soil landscapes 表土有机质是植被来源有机质在土壤中分解持留而形成,当然在农业利用下还可能受到人为有机输入的影响。从土壤形成角度,有机质在土壤中的持留是矿物质与微生物相互作用的结果,这种相互作用以土壤团聚体的形成和更新为媒介[7-8]。在所研究的村域小流域范围内,表土有机碳无论是含量还是形态在不同土壤景观间存在较大变异。与保护的林地相比,因人类耕垦和农业生产管理,小流域内坡地和谷地农田中有机碳总量降低。许多研究报道了自然土壤农业开垦利用后土壤有机碳库的强烈损耗[26-27]。本研究中,伴随着有机碳总量的减少,土壤宏团聚体质量分数下降,团聚体结合态有机碳下降的幅度在园地、旱地和稻田中分别达到84%、73%及74%。也就是说有机碳的损失主要表现为团聚体间结合态有机碳的释放(图3),导致土壤脱团聚化(团聚体稳定性大幅度下降,表2)。这一方面代表了土壤碳释放的“解锁”(unlock)而加剧气候变化[28];另一方面,依附于地理景观,短期的土地利用激活了土壤微生物的分解,在有机碳大量损耗(在园地和旱地中比林地减少了一半以上)的同时,土壤微生物碳利用策略改变[29],有机碳分解能力提高(DOC/MBC比值升高(图3)。土壤微生物生物量代表土壤有机碳中最活跃碳库,与土壤碳氮循环密切相关[30]。土壤微生物商的差异表现出微生物对碳资源竞争的适应。本研究表明,在丘陵山区村域内的小流域中,尽管土壤质地偏沙性,但林地有机质碳含量高达33.99 g/kg,宏团聚体-微团聚体-粉黏粒组成均衡(表3和图2),所以植被保护的林地土壤有机质-团聚体-微生物关系较为协调稳定。反之,山坡地土壤景观因长期耕垦和农业生产管理,土壤中细颗粒比林地增加,土壤团聚体结构退化,微生物生境条件恶化,土壤有机质失去保护而被微生物强烈分解损失,低有机质含量与低团聚体稳定性(平均重量直径)和高的微生物分解能力(DOC/MBC值)代表了失稳的土壤有机质-团聚体-微生物活性关系[30]。既然土壤有机碳强烈损耗是土壤健康变化的主要启动子[31],恢复和增加土壤有机质含量,并促进土壤团聚化是恢复和提高本研究区土壤健康的首要途径。不过,稻田是主要受水分影响下长期水耕培育而成的人为土壤景观,其有机质含量高于园地14%,高于旱地44%,虽然稻田团聚体平均重量直径较小,但稻田保持了与林地相近的土壤微生物生物量,其较低的DOC/MBC值(表3)和团聚体结合碳 (图3左)也说明稻田的有机质-团聚体-微生物关系相对协调,微生物碳资源效率较高,因而稻田是相对土壤固碳和生物活性协调的土壤景观[26,30,32]。也说明,增加土壤有机质碳库可以快速提升土壤生态系统功能[33-34]。 本研究还说明,不同景观下土壤有机质水平与黏粒含量无关,而与土壤颗粒的团聚化有关。未团聚的粉黏粒所持留的有机碳在园地、旱地及稻田3种农业景观土壤间差异较小(图3),黏粒含量最高的旱地(25.5%),土壤保持有机碳反而最少。这可能是Souza等[35]报道的有机碳亏缺时矿物紧密堆积的结果。反过来,不同景观中表土有机碳含量与团聚体有机碳含量呈极显著的正相关关系(图4),这说明土壤团聚化是土壤有机质积累的状态因素,是土壤团聚体化而不是土壤质地本身提供土壤有机质积累的条件。这支持了土壤有机碳积累的上限(即碳保护)是有机碳团聚体物理保护能力所贡献[25,36]。本研究中,林地土壤黏粒含量最低(16%),而保持有机碳最多,主要是宏团聚体的物理保护积累[36]。反之,黏粒含量17%的稻田仍能保持较高的有机碳含量,而黏粒含量分别是20%和25%的园地和旱地,其有机碳含量较低。这可以归因于稻田中宏团聚体和微团聚体中保护的有机碳含量较高,仅次于林地(图3)。在本研究中,林地土壤中是以团聚体保护为主的有机质,而有机碳缺损的旱地中是粉黏粒矿物结合为主的有机质,这里说明了有机碳含量水平与土壤中有机碳结合状态的关联性。也就说,通过土壤团聚体发育而稳定积累有机碳提供了持续固碳的可能性。而增加有机质和促进土壤团聚体化是可以通过土壤管理,特别是有机物循环来实现。这也是最近提出的增进土壤健康,促进食物安全国际计划[37]的战略理念。 本研究表明,有机碳的物理保护是有机碳积累的主要途径,而有机碳损耗对应于微生物分解能力的增强,这从微生物降解与土壤保护两个方面诠释了进入土壤的植物源有机质在不同土壤景观中的存留。TSE溶剂提取和BHY碱水解提取的游离态脂类及结合态脂类代表植物源和微生物源有机质的稳定存留[14]。首先,与林地景观比较,各生物标志物分子丰度在园地和旱地均显著和较大幅度降低,而稻田中又显著高于园地和旱地(表4、图7和图9),这与上述总有机碳的变化趋势吻合。生物标志物相对丰度的变化,反映了这些景观中土壤保护有机物免受分解能力的变化。农业土壤的植物源有机组分贡献一般比自然土壤低[38]。代表高等植物地上部和地下部有机物质输入的角质(cutin)和软木脂(suberin)[39-40],在有机碳含量较高的林地富集,而园地、旱地和稻田中两者丰度都明显下降(表5)。且软木脂与角质的比(S/C)在园地和旱地显著低于林地和稻田,可能提示旱耕促进了地上部更多物质进入土壤。又如,长链脂类(>C20)标志着高等植物蜡质来源[41],而短链脂类( 土壤有机质的酚类分子的提取和鉴定主要是研究稳定有机质中木质素的植被来源和降解状态[47]。香兰基(V)系列单体存在于所有维管植物中,丁香基(S)系列单体主要存在于被子植物中,肉桂基(C)系列单体主要存在于非木质组织中。因此,丁香基与香兰基比值(S/V)和肉桂基与香兰基比值(C/V)常被用来判断不同植物来源的有机物质对土壤有机质的贡献[48]。表6和图10的数据表明,稻田和旱地木质素主要来自被子植物非木本组织的输入,而林地和园地为混合型输入。由于酸是醛氧化分解的产物,酸和醛单体的比值可指示木质素降解程度[49]。此外,3,5-二羟基苯甲酸(3,5-DHBA)及对羟基酚类(P)也是降解的中间产物。因此,3,5-DHBA/V和P/(V+S)也能指示土壤有机质的降解程度[50]。稻田的(Ad/Al)V、3,5-DHBA/V和P/(V+S)等比值低于其他好气土壤景观。淹水下有机质分解较慢,特别是难分解的木质素易于在稻田中持留,这可能是稻田积累有机质的一种表现[51]。 总之,提取态有机质生物标志物分子鉴定结果表明,检测到的植物源生物标志物分子数目和丰度都与土壤有机碳含量有较好的相关性,特别是较为活泼的游离脂类的丰度变化与总有机碳含量变化相吻合(图7)。有报道[52]指出,提取态有机质的分子丰度和分子多样性可以表征不同有机物质输入下的土壤有机质状态。结合前述的不同土壤景观的土壤有机碳在含量、团聚体质量分数和团聚体结合态有机碳分布间的差异,本研究表明,村域小流域内不同土壤景观表土有机质在组成结构上也存在相应的变化格局,生物标志物分子丰度及一些指示性指标可以剖析有机质分子组成的差异。这些差异联系到有机质的保护状态和微生物选择性利用,可能与土壤质量及健康有密切联系,特别是土壤固碳和生物活性[1,5,30]。这应在因地制宜发展乡村农业产业和促进农业碳中和中予以充分重视。同时,本研究也提示,面向土壤固碳和土壤健康,土壤有机质研究中需要更为关注有机质-团聚体-微生物相互作用对土壤有机组分的选择性利用或选择性持留的影响,并结合分子组成研究更深入地表征不同景观条件下土壤有机质的积累状态及其功能[5,7,30]。 供试丘陵山地乡村小流域内不同景观土壤中表土有机碳含量不同,团聚体组成差异也很大,这些差异也反映在提取态有机质的分子组成及丰度上,反映着有机质-团聚体-微生物关系的改变及其对植物源有机质的分解和持留的影响。林地有机质丰富、团聚体间分配均衡、微生物分解性低而有机分子多样性高;在长期耕种影响下,稻田、旱地和园地宏团聚体和微团聚体均趋于破坏,粉黏粒含量增加,植物源有机质强烈分解,有机质以粉黏粒结合态和微生物残留态为主,有机质组分多样性和分子多样性均明显下降。我们认为,增加有机质投入以促进土壤团聚体发育,是提高土壤固碳容量并增进土壤健康的关键途径。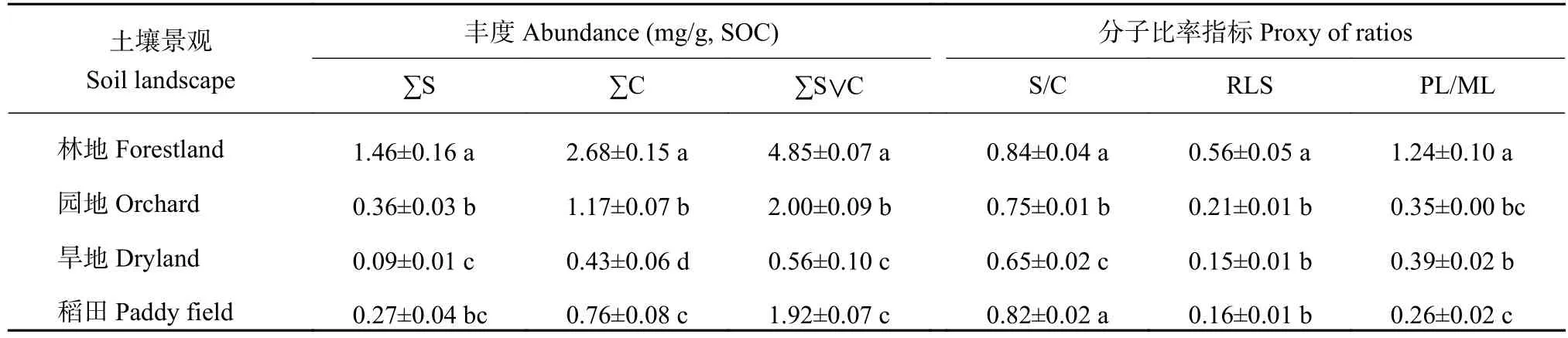
3 讨论
3.1 不同土壤景观下表土有机质变化的数量与库关系
3.2 土壤有机碳变化中的分子组成变化
4 结论
