谷子萌发及苗期生理生化指标对铅胁迫的响应
2022-06-15于雄胜江振岳张文英周国艳王丽芳郭青枝郭春燕
于雄胜 江振岳 张文英 周国艳 王丽芳 郭青枝 郭春燕





摘要:【目的】探究重金屬元素铅(Pb)对谷子萌发及幼苗生理生化特性的影响,为Pb污染背景下谷子的安全生产和品质保障提供理论依据。【方法】以谷子品种晋谷21号为试验材料,采用土培盆栽试验,以自然土壤加水为对照(CK,0 mg/kg),设5个外源Pb浓度处理(50、100、250、500和1000 mg/kg),分析不同Pb浓度处理对谷子萌发及苗期叶片生理生化指标的影响。【结果】随着Pb浓度的增加,谷子的发芽势、发芽率及发芽指数等萌发指标均受到不同程度的抑制,1000 mg/kg处理的相对发芽率仅有52.5%。叶绿素含量随着Pb浓度的增加呈先升高后降低的变化趋势,各Pb胁迫处理均显著高于CK(P<0.05,下同),Pb浓度为500 mg/kg时,叶绿素a、叶绿素b和叶绿素a+b含量均达最大值,较CK分别增加58.3%、69.5%和64.3%。随着Pb浓度的增加,过氧化物酶(POD)活性、质膜透性、丙二醛(MDA)含量及脯氨酸含量均显著增加,其中POD活性对1000 mg/kg高浓度Pb胁迫尤为敏感。可溶性蛋白质含量随着Pb浓度的增加呈先升高后降低的变化趋势,Pb浓度为500 mg/kg时可溶性蛋白质含量达最大值,较CK显著增加54.1%;Pb浓度为1000 mg/kg时,可溶性蛋白质含量急剧下降,与CK相比显著降低14.9%。相关分析结果表明,谷子的发芽势、发芽率和发芽指数均与Pb浓度呈显著负相关,POD活性、质膜透性、MDA含量和脯氨酸含量均与Pb浓度呈极显著正相关(P<0.01)。不同Pb浓度下抗性指标相对值的平均值(敏感性)排序为:POD活性>质膜透性>脯氨酸含量>MDA含量>可溶性蛋白质含量。【结论】谷子在萌发阶段和幼苗时期均会受到Pb胁迫的影响,同时在响应Pb胁迫时,谷子能通过自身生理生化特性的改变提高其适应Pb胁迫的能力,但这种适应性有一定限度。
关键词: 谷子;Pb胁迫;种子萌发;幼苗生理生化指标
中图分类号: S515 文献标志码: A 文章编号:2095-1191(2022)03-0795-08
The germination of foxtail millet seeds and the physiological and biochemical indexes of its seedlings in responses to lead stress
YU Xiong-sheng JIANG Zhen-yue ZHANG Wen-ying ZHOU Guo-yan
WANG Li-fang GUO Qing-zhi GUO Chun-yan
(1Xinzhou Teachers University, Xinzhou, Shanxi 034000, China; 2Maoming Center of Agriculture and Rural Affairs/Maoming Development Center of Fruit Industry, Maoming, Guangdong 525000, China; 3Institute of Dryland
Farming, Hebei Academy of Agriculture and Forestry Sciences, Hengshui, Hebei 053000, China)
Abstract:【Objective】To investigate the effects of heavy metal lead(Pb) on the germination of foxtail millet seeds and the physio-biochemical characteristics of its seedlings so as provide a theoretical basis for the safe production and quality assurance of this type of special rainfed cereal under the background of Pb pollution. 【Method】A pot experiment was conducted using foxtail millet cultivar Jingu 21 as the experimental material to investigate its seed germination and seedling physio-biochemical indexes as influenced by five exogenous Pb concentrations (50, 100, 250, 500, 1000 mg/kg). Soil without added Pb was used as the control (CK, 0 mg/kg). 【Result】The germination energy, germination ratio and germination indexes of Jingu 21 were inhibited with increasing Pb concentrations,the relative germination rate of foxtail millet seeds under Pb treatment of 1000 mg/kg was only 52.5% compared with CK. The contents of chlorophyll increased firstly,and then decreased with increasing Pb concentrations, and all of these kinds of chlorophyll contents were significantly higher than those in the CK(P<0.05,the same below). When Pb concentration was 500 mg/kg,the chlorophyll a,chlorophyll b and chlorophyll a+b contents reached the maximum, which increased by 58.3%, 69.5% and 64.3% respectively compared with CK. As Pb concentration increased,peroxidase (POD) activity,Plasma membrane permeability,malondialdehyde (MDA)content and proline content in leaves of Jingu 21 seedlings increased significantly,among with POD activity was especially sensitive to stress of high Pb concentration. Soluble protein content increased first and then decreased with increa-sing Pb concentrations. When Pb concentration was 500 mg/kg,the soluble protein content reached the highest, which was 54.1% higher than that of CK. When Pb concentration was 1000 mg/kg,soluble protein content decreased sharply, which was 14.9% lower than that of CK. The correlation analysis showed that the germination potential, germination rate and germination index of foxtail millet were negatively correlated with Pb concentrations. The POD activity, plasma membrane permeability, MDA content and proline content were positively correlated with Pb concentration (P<0.01). The average relative value of resistance index (sensitivity) at different Pb concentrations was in the order POD activity>plasma membrane permeabilyty>proline content>MDA content>soluble protein content. 【Conclusion】Millet is affected by Pb stress during both germination and the seedling stage. Within a certain concentration range,foxtail millet can alleviate the damage caused by Pb stress by adjusting its own physiological and biochemical indexes.
Key words: foxtail millet; Pb stress; seed germination; seedling physiological indexes
Foundation items:National Key Research and Development Program of China(2018YFD1000704-6); National Foxtail Millet and Sorghum Industrial Technology System Project During the 14th Five Year Plan Period(CARS-06-14.5-A34);Science and Technology Innovation Project of Colleges and Universities in Shanxi Province(2019L0845,2019 L0840)
0 引言
【研究意义】铅(Pb)为土壤重金属污染“五毒”元素之一,污染普遍、危害严重。2014年全国土壤污染状况调查公报显示,中国土壤Pb污染点位超标率达1.5%(陈能场等,2017)。土壤Pb污染对作物的种子萌发及生长均有一定影响,可造成作物减产、品质降低(Li et al.,2019)。谷子[Setaria italica(L.)Beauv]是起源于中国的传统农作物,具有耐旱、耐瘠等特性,其籽粒小米营养价值丰富(Sharma and Niranjan,2018),为重要的杂粮作物之一。然而,随着土壤污染问题凸显,谷子的产量和品质受到一定程度的威胁,如河北沧州的调查结果显示,小米中Pb含量超标率为20%(李清虹等,2013)。土壤质量直接影响农作物品质和农产品安全,因此,研究土壤Pb污染对谷子萌发及幼苗主要生理生化指標的影响,对谷子Pb污染的防治及品质保障均具有重要意义。【前人研究进展】近年来,已有学者探究了土壤重金属污染对谷子生长和吸收累积的影响,发现重金属污染对谷子的影响与污染元素类型及浓度、研究方法、谷子品种、评价手段等因素相关。张喜文等(2011)采用土培盆栽试验研究谷子幼苗对土壤中Pb、铬(Cr)的生长响应和吸收累积,结果表明,在50~800 mg/kg范围内,Pb、Cr在谷子幼苗地上部和地下部的累积量存在明显差异,谷子幼苗对Pb的吸收富集和转运系数均大于Cr;Pb、Cr不同浓度对幼苗生物量的影响均表现为“低促高抑”的作用。肖志华等(2012)等通过水培试验发现,二价铅离子(Pb2+)对谷子幼苗生理生态的影响和遗传毒害效应均大于二价铜离子(Cu2+),且不同基因型谷子对2种元素的耐性存在差异。张义贤等(2013)从DNA多态性的角度得出,Pb明显影响谷子幼苗细胞中基因组模板DNA的稳定性,DNA多态性变化与Pb浓度存在剂量—效应关系。崔秀秀和张义贤(2015)研究发现,镍(Ni)胁迫可抑制谷子幼苗生长,影响叶片生理指标;Ni胁迫下4种基因型谷子苗期叶片中脯氨酸含量均有不同程度的提高,而可溶性蛋白质含量呈明显下降趋势。王春梅等(2018)以石英砂为基质,通过培养试验研究不同浓度砷(As)对谷子萌发、幼苗生长及抗氧化酶系统的影响,结果表明,低浓度As对谷子萌发和幼苗生长有促进作用,高浓度As则有抑制作用;随着As浓度增加,谷子幼苗超氧化物歧化酶(SOD)活性不断增强,过氧化物酶(POD)和过氧化氢酶(CAT)活性呈先升高后降低的变化趋势。【本研究切入点】Pb胁迫对多种作物的种子发芽率、幼苗生长及生物量均有明显抑制作用,且不同生理生化指标对Pb胁迫的响应与Pb浓度、评价手段等存在一定关系(胡铁柱等,2018;张陆云等,2019;国伟强等,2021)。目前关于Pb胁迫对谷子生理生化指标影响的研究中,缺乏关键抗性指标的敏感性比较分析,也尚不清楚关键抗性指标对不同Pb浓度胁迫响应的差异。【拟解决的关键问题】通过土培盆栽试验,分析不同浓度Pb胁迫对谷子萌发的影响及幼苗关键生理生化指标对Pb胁迫的响应,以期为Pb污染背景下谷子的安全生产和品质保障提供理论依据。
1 材料与方法
1. 1 试验材料
供试谷子品种为山西省优势品种晋谷21号,由山西鑫腾达农业科技有限公司提供。种子种植前,先用5%次氯酸钠消毒15 min,再用蒸馏水冲洗4~5次,然后用滤纸吸干表面水分,待用。供试土壤采自山西省忻州市郊区农田耕层(0~20 cm),为石灰性褐土,自然风干后过2 mm筛,待用。土壤pH 8.43,有机质含量12.7 g/kg,总氮含量0.22 g/kg,最大田间持水量47.0%,土壤Pb本底浓度24.2 mg/kg。试验用试剂均为分析纯。
1. 2 试验方法
试验设5个Pb胁迫处理,土壤外源Pb浓度分别为50、100、250、500和1000 mg/kg。按设定浓度称取不同质量的Pb(NO3)2,分别溶解在200 mL水中,待完全溶解后,分3次加入到称取好的2 kg风干土壤中,混合均匀。对照(CK,0 mg/kg)为自然土壤添加200 mL水。土壤制备好后分别装入同等大小的塑料花盆中(上口径18.0 cm、下口径12.5 cm、高16.0 cm)。所有处理和对照均设3个重复,土壤装盆后放置于室内平衡7 d,备用。挑选经消毒处理后颗粒饱满的谷子种子均匀播种于花盆中,每盆种植35颗。在温室(25 ℃)中培养,采用称重法定期补充水分,并随机更换位置,培养期间自种子露白开始,每天固定时间记录种子萌发情况。待种子萌发完全、生长稳定后间苗,每盆保留8株长势一致的幼苗,待幼苗长至4~5片叶时,选择叶龄相似、形状完好无病害的叶片进行主要生理生化指标测定。
1. 3 测定指标及方法
1. 3. 1 萌发指标计算方法
发芽势(%)=3 d正常发芽的种子数/供试种子数×100
发芽率(%)=7 d正常发芽的种子数/供试种子数×100发芽指数=∑G/D
式中,G为t时间内发芽数,D为相应的发芽天数。
相对发芽率(%)=处理发芽率/对照发芽率×100
1. 3. 2 苗期叶片生理生化指标测定 采用丙酮和乙醇浸提法测定叶绿素含量,采用愈创木酚比色法测定POD活性,采用相对电导率法测定质膜透性,采用硫代巴比妥酸比色法测定丙二醛(MDA)含量,采用茚三酮显色法测定脯氨酸含量,采用考马斯亮蓝G-250染色法测定可溶性蛋白质含量(高俊凤,2006)。各抗性指标相对值计算公式为:
抗性指标相对值=处理指标值/对照指标值
1. 4 统计分析
采用Excel 2016进行数据整理及制图。采用DPS 15.1对数据进行差异显著性检验(LSD法)和简单相关分析(Pearson法)。相关分析中各个指标值以3次重复均值分别与对应的Pb浓度进行相关分析,包含6个Pb浓度梯度。
2 结果与分析
2. 1 不同Pb胁迫处理对谷子萌发的影响
由表1可知,随着Pb浓度的增加,谷子的发芽势、发芽率、发芽指数及相对发芽率均逐渐降低。从发芽势来看,50 mg/kg Pb浓度处理与CK无显著差异(P>0.05,下同),100~1000 mg/kg处理的发芽势较CK显著降低12.7%~47.8%(P<0.05,下同)。发芽率和发芽指数的变化规律与发芽势相似,与CK相比,100、250、500和1000 mg/kg处理的发芽率分别显著降低8.1%、34.4%、42.4%和47.5%,发芽指数分别显著降低11.6%、33.9%、44.3%和48.4%。低浓度(50和100 mg/kg)Pb胁迫下,发芽势比发芽率下降更明显;高濃度(250、500和1000 mg/kg)Pb胁迫下,相对发芽率明显降低,Pb浓度为1000 mg/kg时,相对发芽率仅有52.5%。由此可见,Pb胁迫对谷子萌发有明显的抑制作用。
2. 2 不同Pb胁迫处理对谷子苗期叶片叶绿素含量的影响
由表2可知,不同Pb浓度处理的谷子苗期叶片叶绿素a、叶绿素b和叶绿素a+b含量均较CK显著增加,但叶绿素a/b无显著差异;随着Pb浓度的增加,叶绿素含量呈先升高后降低的变化趋势。其中,50、100和250 mg/kg 3个Pb浓度处理间的叶绿素a和叶绿素b含量无显著差异;Pb浓度为500 mg/kg时,叶绿素a、叶绿素b和叶绿素a+b含量均达最大值,较CK分别增加58.3%、69.5%和64.3%;Pb浓度继续增大到1000 mg/kg时,叶绿素含量较500 mg/kg处理有所降低,但仍显著高于CK及其他处理。
2. 3 不同Pb胁迫处理对谷子苗期叶片POD活性的影响
POD是植物细胞内活性氧清除酶系统中重要的保护酶,是研究植物抗逆反应的重要指标之一(Yang et al.,2015)。由图1可看出,土壤中添加外源Pb后,谷子苗期叶片的POD活性较CK显著增强。在50~250 mg/kg外源Pb浓度范围内,POD活性变化不显著;当土壤外源Pb浓度达500 mg/kg及以上时,POD活性显著增加,500和1000 mg/kg处理的POD活性较CK增加4.0和22.6倍。由此可见,谷子苗期叶片中POD活性对高浓度Pb胁迫尤为敏感。
2. 4 不同Pb胁迫处理对谷子苗期叶片细胞质膜透性的影响
质膜透性是衡量植物细胞质膜受到伤害的重要生理指标之一,其值越大表示质膜受到的伤害越大,而叶片细胞浸提液中相对电导率的变化情况可反映质膜受伤害程度(王马勃等,2019)。由图2可看出,随着Pb浓度的增加,谷子苗期叶片中相对电导率呈逐渐上升趋势。Pb浓度为50 mg/kg时,相对电导率略有增加,但与CK无显著差异;当土壤外源Pb浓度达100 mg/kg及以上时,相对电导率显著增加,较CK增加0.4~1.2倍。表明随着Pb浓度的增加,谷子苗期叶片细胞质膜受到的伤害随之增大。
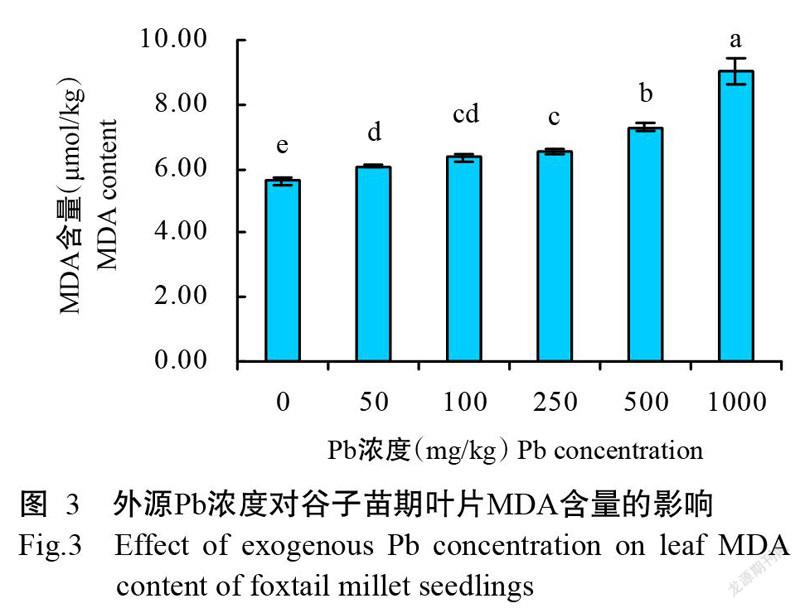
2. 5 不同Pb胁迫处理对谷子苗期叶片MDA含量的影响
MDA是膜脂过氧化最重要的产物之一,其含量可表征植物在逆境胁迫下细胞质膜的受损程度(Khalil et al.,2021)。图3结果显示,不同外源Pb浓度处理的叶片MDA含量均显著高于CK,50~1000 mg/kg处理的MDA含量较CK增加7.8%~60.4%。其中,50和100 mg/kg及100和250 mg/kg处理间无显著差异;Pb浓度为500和1000 mg/kg时,MDA含量显著高于其他处理。
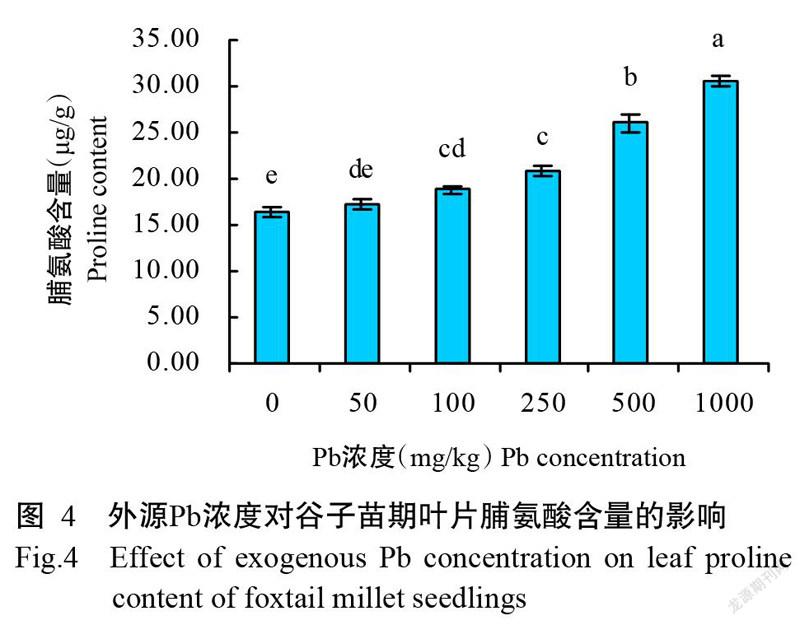
2. 6 不同Pb胁迫处理对谷子苗期叶片脯氨酸含量的影响
脯氨酸是植物适应逆境时重要的渗透压调节物质,具有保持细胞渗透平衡,增加蛋白质的可溶性,进而保护大分子稳定性的功能(Kaur and Asthir,2015)。图4结果显示,当土壤中外源Pb浓度为50 mg/kg时,谷子苗期叶片中脯氨酸含量与CK无显著差异;当Pb浓度达100 mg/kg及以上时,脯氨酸含量开始显著增加,较CK增加0.2~0.9倍,但100和250 mg/kg处理间无显著差异。由此可知,在高浓度Pb胁迫下,谷子苗期叶片中脯氨酸含量会有明显升高。
2. 7 不同Pb胁迫处理对谷子苗期叶片可溶性蛋白质含量的影响
由图5可看出,随着Pb浓度的增加,谷子苗期叶片中可溶性蛋白质含量呈先升高后降低的变化趋势。Pb浓度为50 mg/kg时,可溶性蛋白质含量与CK无显著差异;Pb浓度为100和250 mg/kg时,2个Pb浓度处理间无显著差异,但与CK相比二者可溶性蛋白质含量显著增加,增幅分别为22.1%和32.6%;Pb浓度为500 mg/kg时,可溶性蛋白质含量达最大值,较CK显著增加54.1%;Pb浓度为1000 mg/kg时,可溶性蛋白质含量急剧下降,与CK相比显著降低14.9%。
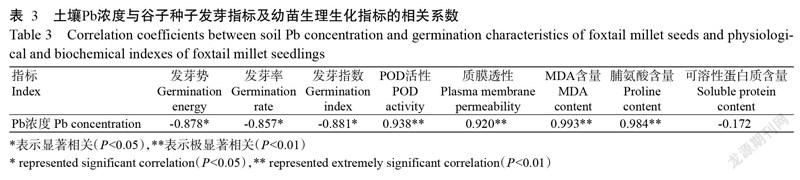
2. 8 土壤Pb浓度与谷子发芽特性及幼苗生理生化指标的相关分析
将土壤Pb浓度与谷子各发芽特性及幼苗生理生化指标进行简单相关分析,结果(表3)表明,谷子的发芽势、发芽率和发芽指数均与Pb浓度呈显著负相关,POD活性、质膜透性、MDA含量和脯氨酸含量均与Pb浓度呈极显著正相关(P<0.01),可溶性蛋白质含量与Pb浓度呈不显著负相关。
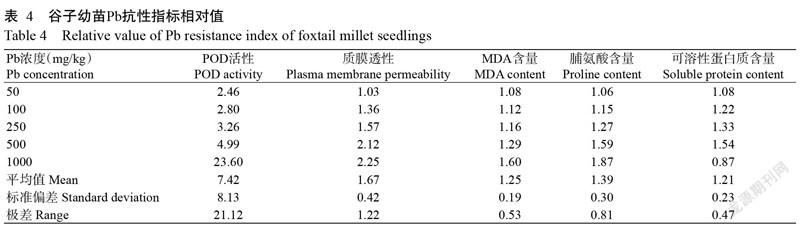
2. 9 谷子幼苗Pb抗性指标敏感性分析
为了更好的评价各个抗性指标的敏感性和适用范围,进一步比较外源Pb胁迫下谷子苗期叶片中POD活性、质膜透性、MDA含量、脯氨酸含量、可溶性蛋白质含量等抗性指标的相对值,其值越大说明该指标敏感性越高。由表4结果可知,不同Pb浓度下抗性指标相对值的平均值(敏感性)排序为:POD活性>质膜透性>脯氨酸含量>MDA含量>可溶性蛋白质含量;标准偏差除可溶性蛋白质含量略大于MDA含量外,其他整體排序与平均值相似。
3 讨论
种子萌发过程是植物响应外界环境变化较敏感的时期,种子萌发情况直接关系到植物幼苗的形态、后期植物的生长发育及作物的产量和品质。因此,种子萌发阶段通常用来研究作物对重金属的耐性和响应机制(张陆云等,2019;刘翰升等,2020)。本研究结果显示,Pb胁迫对谷子萌发具有明显的抑制作用,且低浓度Pb胁迫对谷子发芽势的影响比对发芽率的影响更明显,Pb浓度与发芽势的相关性高于发芽率,表明Pb胁迫对谷子的毒害作用在种子萌发初期就会发生,一直到完成整个萌发过程,且在萌发初期的影响更大,其原因可能是Pb胁迫加速消耗了种子中贮藏的营养物质并破坏了细胞质膜透性。
Pb胁迫不仅对种子萌发有影响,而且对植物幼苗的生理生化指标也有一定影响(王马勃等,2019)。本研究中,谷子苗期叶片中叶绿素含量、POD活性、质膜透性、MDA含量、脯氨酸含量和可溶性蛋白质含量等生理生化指标均与Pb浓度存在一定的剂量—效应关系。叶绿素是植物叶片吸收、转化、利用光能的主要色素,其含量及合成速率是判断植物耐受逆境的重要指标(黄晓霞等,2017)。叶绿素在响应重金属胁迫时,其含量的变化趋势与植物类型、反应时间等均有一定关系。有研究报道,Pb浓度与3种经济树种的叶绿素含量存在显著负相关(刘建华等,2020)。本研究中,叶绿素a、叶绿素b和叶绿素a+b含量均随着Pb浓度的增大呈先升高后降低的变化趋势,在Pb浓度为500 mg/kg时达最大,1000 mg/kg时开始显著下降。与王马勃等(2019)报道Pb胁迫下茅苍术叶绿素含量随处理时间的延长呈先升高后降低的趋势相似。Pb胁迫下植物叶绿素含量的增大可能是植物叶绿素合成系统对重金属的一种应急性反应(黄晓霞等,2017)。此外,叶绿素a/b在各处理中均无显著差异,可能与叶绿素a和叶绿素b对Pb胁迫的响应一致有关。石慧芳等(2021)对黑麦草的重金属元素抗性研究结果也得出了相似结果。
抗氧化酶类可清除植物在逆境条件下产生的活性氧自由基(Kibria et al.,2017)。但当高浓度重金属胁迫达到一定程度时,就会破坏植物正常的酶功能,进而导致酶系统清除活性氧能力下降,引发细胞生理功能紊乱(安婷婷等,2021)。有研究显示,随着土壤Pb浓度的升高,高羊茅的POD活性呈先升高后降低的变化趋势,在1000 mg/kg时达最大值;狗牙根的POD活性则持续升高,在2000 mg/kg时达最大值(熊作明等,2019)。本研究中,随着Pb浓度的增加,谷子幼苗叶片的POD活性显著升高,Pb浓度为1000 mg/kg时,POD活性较CK增加22.6倍,远高于其他处理,该结果与Pb低积累小麦受Pb胁迫下POD活性的变化规律相似(胡铁柱等,2018),究其原因可能主要与不同植物基因型有关,也可知POD活性为谷子关键耐性指标。
相对电导率和MDA含量均可反映质膜系统的受伤害程度(王马勃等,2019;Khalil et al.,2021)。植物体内的可溶性蛋白质主要是一些游离态代谢酶类,可作为渗透调节物质与重金属离子结合,形成一种重金属结合蛋白,减少游离态重金属离子的含量,从而减轻重金属对植物的损伤(谭万能等,2006)。脯氨酸是逆境条件下膜和酶的保护物质及自由基清除剂,可通过改变细胞液浓度及细胞渗透势来防止细胞脱水(Kaur and Asthir,2015)。因此,逆境条件下植物体内脯氨酸含量通常会增加,以增加植物幼苗抗Pb胁迫的能力(Rizvi et al.,2018)。本研究结果表明,相对电导率和MDA含量均随着Pb浓度的增加而显著增加,与张义贤等(2013)的研究结果相吻合。相对电导率的增大和MDA含量的升高,说明在Pb胁迫下细胞内自由基积累增多,膜脂过氧化程度增强,此时植物通过细胞内各种保护机制的增强来缓解Pb胁迫作用。此外,本研究中可溶性蛋白质含量在Pb浓度为500 mg/kg时最高,1000 mg/kg时显著降低且低于CK,其原因可能在于过量的重金属离子可损伤与蛋白质合成相关的细胞器,使植物中蛋白合成酶下降,分解酶活性上升,从而抑制蛋白质的合成,导致可溶性蛋白质含量下降(国伟强等,2021)。同时,植物中MDA和脯氨酸含量增加,质膜透性增大,细胞中的可溶性物质外渗,重金属更易进入细胞并破坏蛋白结构而使其降解,也表现为蛋白质含量的降低,因此可溶性蛋白质含量可作为植物受到重度污染胁迫的指示指标(侯明等,2018)。可溶性蛋白质和脯氨酸含量的改变,提示谷子幼苗在Pb浓度较低时会通过各种生理生化改变来提高自身抗逆性,以适应不利环境的影响,而当这种胁迫超过一定范围后,其自身的胁迫响应防护措施会相应减弱。就谷子幼苗而言,这种减弱作用首先表现在可溶性蛋白质含量上,Pb浓度为1000 mg/kg时可溶性蛋白质含量显著降低,而此浓度下脯氨酸含量还在进一步积累,说明谷子不同生理生化指标在敏感性上存在差异。相关分析结果表明,谷子幼苗叶片中可溶性蛋白质含量与土壤Pb浓度无显著相关性,但研究中发现若去除1000 mg/kg处理后再进行相关分析,可得出可溶性蛋白质含量与土壤Pb浓度呈显著正相关,其原因如前所述,即在重度Pb胁迫下,可溶性蛋白质含量会表现为降低现象,从而导致结果存在差异。
由上述分析并结合抗性指标的敏感性分析结果可知,谷子各生理生化指标对不同Pb浓度的响应存在一定差异。在研究重金属污染对谷子生理生化指标响应时,应重点关注敏感性高的指标。
4 结论
Pb胁迫对谷子萌发存在明显的抑制作用,谷子幼苗各生理生化抗性指标对Pb胁迫的响应存在一定差异,其敏感性顺序为:POD活性>质膜透性>脯氨酸含量>MDA含量>可溶性蛋白质含量,其中可溶性蛋白质含量可作为Pb重度胁迫的指示指标。由此可见,谷子在萌发阶段和幼苗时期均会受到Pb胁迫的影响,同时在响应Pb胁迫时,谷子能通过自身生理生化特性的改变提高其适应Pb胁迫的能力,但这种适应性有一定限度。
参考文献:
安婷婷,黄帝,王浩,张一,陈应龙. 2021. 植物响应镉胁迫的生理生化机制研究进展[J]. 植物学报,56(3):347-362. [An T T,Huang D,Wang H,Zhang Y,Chen Y L. 2021. Research advances in plant physiological and biochemical mechanisms in response to cadmium stress[J]. Chinese Bulletin of Botany,56(3):347-362.] doi:10.11983/CBB20160.
陳能场,郑煜基,何晓峰,李小飞,张晓霞. 2017. 《全国土壤污染状况调查公报》探析[J]. 农业环境科学学报,36(9):1689-1692. [Chen N C,Zheng Y J,He X F,Li X F,Zhang X X. 2017. Analysis of the Report on the national general survey of soil contamination[J]. Journal of Agro-Environment Science,36(9):1689-1692.] doi:10.11654/jaes.2017-1220.
崔秀秀,张义贤. 2015. Ni胁迫对不同基因型谷子幼苗生长及氮素代谢的影响[J]. 生态学报,35(10):3244-3251. [Cui X X,Zhang Y X. 2015. Effect of Ni on the growth and nitrogen metabolism in foxtail millet seedlings of different genotypes[J]. Acta Ecologica Sinica,35(10):3244-3251.] doi:10.5846/stxb201310082415.
高俊凤. 2006. 植物生理学实验指导[M]. 北京:高等教育出版社. [Gao J F. 2006. A guide to plant physiological experiment[M]. Beijing:Higher Education Press.]
国伟强,杜雪,彭玉兰,李源恒,赵春莉,曲同宝. 2021. Cd、Pb复合胁迫对狼尾草种子萌发及幼苗生长的影响[J]. 种子,40(11):91-96. [Guo W Q,Du X,Peng Y L,Li Y H,Zhao C L,Qu T B. 2021. Effects of combined stress of Cd and Pb on seed germination and seedling growth of Pennisetum alopecuroides[J]. Seed,40(11):91-96.] doi:10.16590/j.cnki.1001-4705.2021.11.091.
黄晓霞,匡经舸,李琬婷,管菊,程小毛. 2017. 铅镉胁迫对小叶榕叶片细胞超微结构的影响[J]. 西南林业大学学报,37(6):41-47. [Huang X X,Kuang J G,Li W T,Guan J,Cheng X M. 2017. Effects of Pb2+and Cd2+ stresses on leaf ultrastructure of Ficus parvifolia[J]. Journal of Southwest Forestry University,37(6):41-47.] doi:10.11929/j.issn.2095-1914.2017.06.007.
侯明,王秀丽,杨心瀚,吕志超. 2018. 外源钒胁迫下甜玉米植物可溶性蛋白含量的动态变化[J]. 环境化学,37(6):1217-1222. [Hou M,Wang X L,Yang X H,Lü Z C. 2018. Dynamic changes of soluble protein contents in the sweet corn under exogenous vanadium stress[J]. Environmental Chemistry,37(6):1217-1222.] doi:10.7524/j.issn.0254-6108.2017090104.
胡铁柱,孙海燕,杨靖,游晴晴,茹振钢. 2018. 铅对不同铅积累类型小麦幼苗生理生化特性和根系生长的影响[J]. 河南农业科学,47(11):13-17. [Hu T Z,Sun H Y,Yang J,You Q Q,Ru Z G. 2018. Effect of Pb on physiological and biochemical properties and root growth of two Pb accumulation types of wheat seedlings treated with Pb[J]. Journal of Henan Agricultural Sciences,47(11):13-17.] doi:10.15933/j.cnki.1004-3268.2018.11.003.
李清虹,李清峰,张巍,赵明,王刚,吴海龙. 2013. 沧州市农田土壤及农作物中重金属调查[J]. 环境与健康杂志,30(9):758. [Li Q H,Li Q F,Zhang W,Zhao M,Wang G,Wu H L. 2013. Investigation of heavy metals in farmland soil and crops in Cangzhou[J]. Journal of Environment and Health,30(9):758.] doi:10.16241/j.cnki.1001-5914. 2013.09.013.
刘翰升,赵春莉,刘玥,国伟强,王蕾. 2020. Cd胁迫对波斯菊种子萌发、幼苗耐性及富集的影响[J]. 河南农业科学,49(5):126-133. [Liu H S,Zhao C L,Liu Y,Guo W Q,Wang L. 2020. Seed germination,seedling tolerance and enrichment effect of Cosmos bipinnata under cadmium stress[J]. Journal of Henan Agricultural Sciences,49(5):126-133.] doi:10.15933/j.cnki.1004-3268.2020.05.016.
刘建华,朱济友,姚姜铭,李夏榕,覃国铭,于强. 2020. 土壤重金属胁迫对经济树种的光合响应特征研究[J]. 农业机械学报,51(9):211-216. [Liu J H,Zhu J Y,Yao J M,Li X R, Qin G M,Yu Q. 2020. Photosynthetic response characteristics of economic tree species to heavy metal stress in soil[J]. Transactions of the Chinese Society for Agricultural Machinery,51(9):211-216.] doi:10.6041/j.issn.1000-1298.2020.09.024.
石慧芳,席溢,张茂,裴应杰,黄本用,李金孟,杨克超. 2021. 铅、锌、镉单一及复合胁迫对多年生黑麦草生长及生理特性的影响[J]. 南方农业学报,52(4):1030-1039. [Shi H F,Xi Y,Zhang M,Pei Y J,Huang B Y,Li J M,Yang K C. 2021. Effects of single and combined stresses of lead,zinc and cadmium on growth and physiological characteristics of perennial ryegrass[J]. Journal of Sou-thern Agriculture,52(4):1030-1039.] doi:10.3969/j.issn. 2095-1191.2021.04.021.
譚万能,李志安,邹碧. 2006. 植物对重金属耐性的分子生态机理[J]. 植物生态学报,30(4):703-712. [Tan W N,Li Z A,Zou B. 2006. Molecular mechanisms of plant tole-rance to heavy metals[J]. Chinese Journal of Plant Eco-logy,30(4):703-712.]
王马勃,巢建国,谷巍,陆奇杰,张文明,孙永娣. 2019. 铅胁迫下茅苍术生理指标、光合参数及生物量的变化[J]. 南方农业学报,50(1):32-39. [Wang M B,Chao J G,Gu W,Lu Q J,Zhang W M,Sun Y D. 2019. Changes of physiological indexes,photosynthetic parameters and biomass of Atractylodes lancea(Thunb.) DC. under lead stress[J]. Journal of Southern Agriculture,50(1):32-39.] doi:10. 3969/j.issn.2095-1191.2019.01.05.
王春梅,闫双堆,卜玉山,刘利军,张乃于. 2018. As对谷子萌发、幼苗生长及抗氧化酶系统的影响[J]. 水土保持学报,32(6):352-358. [Wang C M,Yan S D,Bu Y S,Liu L J,Zhang N Y. 2018. Effect of As stress on millet germination,seedling growth and antioxidant system[J]. Journal of Soil and Water Conservation,32(6):352-358.] doi:10.13870/j.cnki.stbcxb.2018.06.051.
肖志华,张义贤,张喜文,李萍. 2012. 外源铅、铜胁迫对不同基因型谷子幼苗生理生态特性的影响[J]. 生态学报,32(3):889-897. [Xiao Z H,Zhang Y X,Zhang X W,Li P. 2012. Effects of exogenous Pb and Cu stress on eco-physiological characteristics on foxtail millet seedlings of different genotypes[J]. Acta Ecologica Sinica,32(3):889-897.] doi:10.5846/stxb201108281256.
熊作明,杨佳欢,王丽楠. 2019. 3种草坪草对土壤重金属铅、镉胁迫的响应[J]. 扬州大学学报(农业与生命科学版),40(6):117-121. [Xiong Z M,Yang J H,Wang L N. 2019. Response of three turfgrasses to soil Pb and Cd stresses[J]. Journal of Yangzhou University(Agricultural and Life Science Edition),40(6):117-121.] doi:10.16872/ j.cnki.1671-4652.2019.06.020.
张陆云,任艳芳,何俊瑜,王伟,崔志文,王羲玥,张黎明. 2019. 不同品种小白菜萌发期对铅胁迫的耐性差异及其生理机制[J]. 植物生理学报,55(10):1467-1480. [Zhang L Y,Ren Y F,He J Y,Wang W,Cui Z W,Wang X Y,Zhang L M. 2019. Tolerance and physiological mechanism of different varieties of pakchoi to lead stress at germination stage[J]. Plant Physiology Journal,55(10):1467- 1480.] doi:10.13592/j.cnki.ppj.2019.0351.
张喜文,张义贤,李萍,肖志华,杨磊. 2011. 谷子幼苗对土壤铅、铬的生长响应及吸收积累的差异性[J]. 植物研究,31(6):739-743. [Zhang X W,Zhang Y X, Li P,Xiao Z H,Yang L. 2011. Differences in growth response to Cr,Pb stress and their uptake and accumulation of foxtail millet seedlings[J]. Bulletin of Botanical Research,31(6):739-743.]
张义贤,付亚萍,肖志华,张喜文,李萍. 2013. 铅胁迫对不同基因型谷子幼苗生理特性及基因组DNA多态性的影响[J]. 农业环境科学学报,32(3):478-484. [Zhang Y X,Fu Y P,Xiao Z H,Zhang X W,Li P. 2013. Effects of Pb stress on physiological characteristics and DNA polymorphism of genome in different geno types foxtail millet[J]. Journal of Agro-Environment Science,32(3):478-484.] doi:10.11654jaes. 2013.03.03.01.
Kaur G,Asthir B. 2015. Proline:A key player in plant abiotic stress tolerance[J]. Biologia Plantarum,59(4):609-619. doi:10.1007/s10535-015-0549-3.
Khalil R,Haroun S,Bassyoini F,Nagah A,Yusuf M. 2021. Salicylic acid in combination with kinetin or calcium ameliorates heavy metal stress in Phaseolus vulgaris plant[J]. Journal of Agriculture and Food Research,5:100182. doi:10.1016/j.jafr.2021.100182.
Kibria M G,Hossain M,Murata Y,Hoque M A. 2017. Antioxidant defense mechanisms of salinity tolerance in rice genotypes[J]. Rice Science,24(3):155-162. doi:10.1016/ j.rsci.2017.05.001.
Li H,Li M,Zhao D,Li J,Li S,Juhasz A L,Basta N T,Luo Y,Ma L Q. 2019. Oral bioavailability of As,Pb,and Cd in contaminated soils,dust,and foods based on animal bioassays:A review[J]. Environmental Science & Technology,53(18):10545-10559. doi:10.1021/acs.est.9b03567.
Rizvi A,Khan M S. 2018. Heavy metal induced oxidative damage and root morphology alterations of maize(Zea mays L.)plants and stress mitigation by metal tolerant nitrogen fixing azotobacter chroococcum[J]. Ecotoxicology and Environmental Safety,157:9-20. doi:10.1016/j.eco-env.2018.03.063.
Sharma N,Niranjan K. 2018. Foxtail millet:Properties,processing,health benefits,and uses[J]. Food Reviews International,34(4):329-363. doi:10.1080/87559129.2017.12 90103.
Yang H J,Mu J Y,Chen L C,Feng J,Hu J L,Li L,Zhou J M,Zuo J R. 2015. S-Nitrosylation positively regulates ascorbate peroxidase activity during plant stress responses[J]. Plant Physiology,167(4):1604-1615. doi:10.1104/pp.114.255216.
(責任编辑 王 晖)
