海拔梯度对贺兰山岩羊主要活动区植物群落特征的影响
2022-06-11赵朋波邱开阳谢应忠刘王锁李小伟陈林王继飞孟文芬黄业芸李小聪杨浩楠
赵朋波,邱开阳*,谢应忠,刘王锁,李小伟,陈林,王继飞,孟文芬,黄业芸,李小聪,杨浩楠
(1. 宁夏大学农学院,宁夏 银川 750021;2. 宁夏大学生态环境学院,宁夏 银川 750021;3. 宁夏大学西北土地退化与生态恢复省部共建国家重点实验室培育基地,宁夏 银川 750021;4. 宁夏贺兰山国家级自然保护区管理局,宁夏 银川 750021)
贺兰山地处阿拉善高原和银川平原之间,是我国重要地理分界线,区隔着我国季风区与非季风区、温带草原与荒漠草原[1−2],也是青藏高原、蒙古高原及华北平原植物区系的重要枢纽[3],是干旱内陆地区为数不多的富有植物多样性的地区之一[4]。贺兰山作为我国西北地区一道重要的生态屏障,既阻碍了腾格里沙漠的东进,又拦截了来自乌兰布和沙漠的风沙,有效地保护了银川平原[3]。因此,贺兰山对于维护中国西北乃至整个西部地区的生态安全具有重要的战略意义。
在贺兰山,由于雪豹等天敌消失[5]、禁牧措施等原因导致种间竞争减弱[6],使得岩羊(Pseudois nayaur)数量快速增长[6],这种单一物种种群快速增长诱发了诸多问题,造成了与同等生态位上其他物种的关系失衡[7],严重影响到植被的恢复和其他动物的发展[8]。岩羊作为植物−岩羊−雪豹这一食物链中重要的一环,一旦超过自然界的承载能力,势必引起贺兰山生态环境的破坏,如草场面积锐减、土地沙漠化加剧等问题[9]。
贺兰山自然保护区的植被垂直分异特征明显,其植被类型沿着海拔上升依次为荒漠草原、山地疏林草原、山地针叶林和高山−亚高山灌丛草甸[4,10−11]。贺兰山东坡是岩羊的主要分布区[6]。贺兰山岩羊主要分布在海拔为1100~2800 m 的地区,这些地区多是以灰榆(Ulmus glaucescens)等乔木为主的山地疏林草原,或无高大乔木,或接近水源,并远离人类活动干扰,距离裸岩2~5 m,位于半阴半阳坡的下坡位[12−13]。另外还有高山−亚高山灌丛草甸[14−15]也是岩羊偏爱的取食地和栖息地。
在山地生态系统中,海拔对其生态−水文过程有着强烈的影响,这也导致了植物群落特征随海拔变化而有所不同[16]。同时,与海拔密切相关的气候的变化也对植物群落有制约作用,也是植物多样性变化的主要驱动因子[17]。现有研究表明,山地植物物种丰富度的模型主要有两种:一是物种丰富度随海拔升高而升高[18],二是随海拔的升高,物种丰富度先升高后降低[19]。这是与海拔相关的水分、温度等环境因子的变化所导致。在不同海拔地区,水分主导了植物的生长,水分对植物的限制性随着海拔的降低而增强。总的来说,海拔变化囊括了水分、温度、光照、湿度和土壤等环境因子的变化,对植物群落的多样性存在显著影响[20]。
为了探明贺兰山岩羊主要活动区的植物群落特征,进而为理清岩羊与植被的平衡关系奠定基础,以期加强对贺兰山地区岩羊种群的合理管护,从而维护贺兰山生态平衡,本研究以位于贺兰山东坡的岩羊主要活动区植被群落为对象,通过对不同海拔的岩羊主要活动区进行植被调查,研究其物种组成、物种多样性变化,试图说明:1)贺兰山岩羊主要活动区不同植被类型的物种组成;2)植物物种多样性沿海拔梯度的变化规律。
1 材料与方法
1.1 研究区概况
研究区位于宁夏贺兰山国家级自然保护区苏峪口。贺兰山横亘于阿拉善高原和银川平原之间,东临黄河,西靠腾格里沙漠,属于干旱半干旱地区。地理坐标为38°13′−39°30′ N,105°41′−106°41′ E,是我国季风区与非季风区、内流区与外流区的地理分界线。贺兰山大体为西南−东北走向,东西宽20~40 km,南北约180 km,东坡山势陡峭,西坡相对平缓,从山麓到山顶垂直分化明显,高度相差2000 m 以上,主体海拔大多在2000~3000 m 之间,主峰敖包疙瘩高3556.1 m。目前建立了宁夏贺兰山国家级自然保护区(东坡)和内蒙古贺兰山国家级自然保护区(西坡)。
贺兰山属于典型的大陆性季风气候,冬季漫长且寒冷,夏季短暂而炎热,气候较为干燥,全年盛行西北风,年平均风速7.5 m·s−1,大风天数达157.7 d,最大风速38.7 m·s−1。贺兰山的水热条件随着海拔上升有着明显的差异,东坡年平均气温8.2 ℃,年平均降水量183.3 mm,西坡年均气温7.6 ℃,年均降水量430 mm,降水主要集中在6−9月,按水热条件,贺兰山地区可被划分为4 个垂直气候带:温暖干旱草原荒漠气候带(<1500 m)、较温凉半干旱草原气候带(1600~2000 m)、温寒半干旱偏湿润森林气候带(2000~3000 m)、寒冷半湿润高山−亚高山气候带(>3000 m)。主要植被类型随着海拔升高逐渐由山地草原带(1000~1600 m)、山地疏林草原带(1600~2000 m)过渡到山地针叶林带(2000~2500 m)和高山−亚高山灌丛、草甸带(>2500 m)[21]。
1.2 样地设置
于2020年8月在贺兰山国家级自然保护区苏峪口开展试验。结合以往在贺兰山开展的与岩羊相关的研究[6,12]及实地勘察,并根据植被类型情况,确定贺兰山岩羊主要活动区4 个,从山麓到山顶分别为荒漠草原、山地疏林草原、山地针叶林、高山−亚高山灌丛草甸(表1)。在每个活动区设置3 个样地,每个样地布设具有代表性的套样方,即3 个乔木样方(20 m×20 m)内布设灌木样方(10 m×10 m)5 个,每个灌木样方内各设置1 个草本样方(1 m×1 m),样方均设置在阳坡。运用植物群落学调查技术,调查植物种类、高度、盖度、频度、多度以及地上生物量,灌木和乔木植物补充测定其冠幅、胸径等[22]。
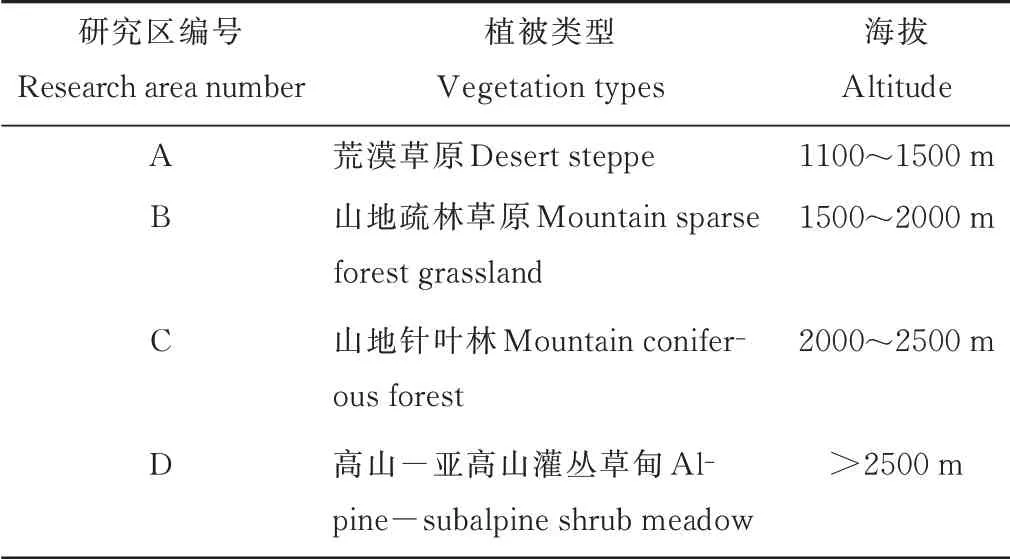
表1 研究区植物群落类型及海拔高度Table 1 Plant community types and the altitude of research areas
1.3 数据分析
物种多样性指数采用物种丰富度指数(R)、Simpson 优势度指数(D)、Shannon−Wiener 多样性指数(H′)和Pielou 均匀度指数(E),根据以下公式计算不同植物群落数量特征:

式中:S为群落中物种总数;Pi为种i的相对重要值,i=1,2,……,S。
本研究首先使用Excel 2019 对数据进行初步整理、计算各研究区内植物的重要值和多样性指数,然后利用SPSS 19.0 进行单因素分析(One-way ANOVA)和回归分析,对不同海拔的数据进行差异显著性检验、多重比较和回归分析等,所有图表均在Excel 2019 中完成。
2 结果与分析
2.1 贺兰山岩羊主要活动区植被组成及区系特征
贺兰山岩羊主要活动区共记录到30 科62 属87 种植物。其中被子植物84 种,裸子植物3 种。贺兰山岩羊主要活动区所有植物中有3 个科所含种的数量在10 种以上(表2),其中含种数量最多的是禾本科(Poaceae),有20种,占总种数比例为22.98%,其次是蔷薇科(Rosaceae)(11 种)和菊科(Asteraceae)(10 种),两科合计占总种数比24.14%,然后依次是豆科(Fabaceae)(7 种)、石竹科(Caryophyllaceae)(4 种)、伞形科(Apiaceae)(3 种)、玄参科(Scrophulariaceae)(3 种)、苋科(Amaranthaceae)(3 种)、石蒜科(Amaryllidaceae)(2 种)、旋花科(Convolvulaceae)(2种)、毛茛科(Ranunculaceae)(2 种)、唇形科(Lamiaceae)(2 种)。另外还有18 个单种科,包括远志科(Polygalaceae)、鸢尾科(Iridaceae)、忍冬科(Caprifoliaceae)、茜草科(Rubiaceae)、榆科(Ulmaceae)、小檗科(Berberidaceae)和松科(Pinaceae)等,所有单种科数占总科数的60.00%。
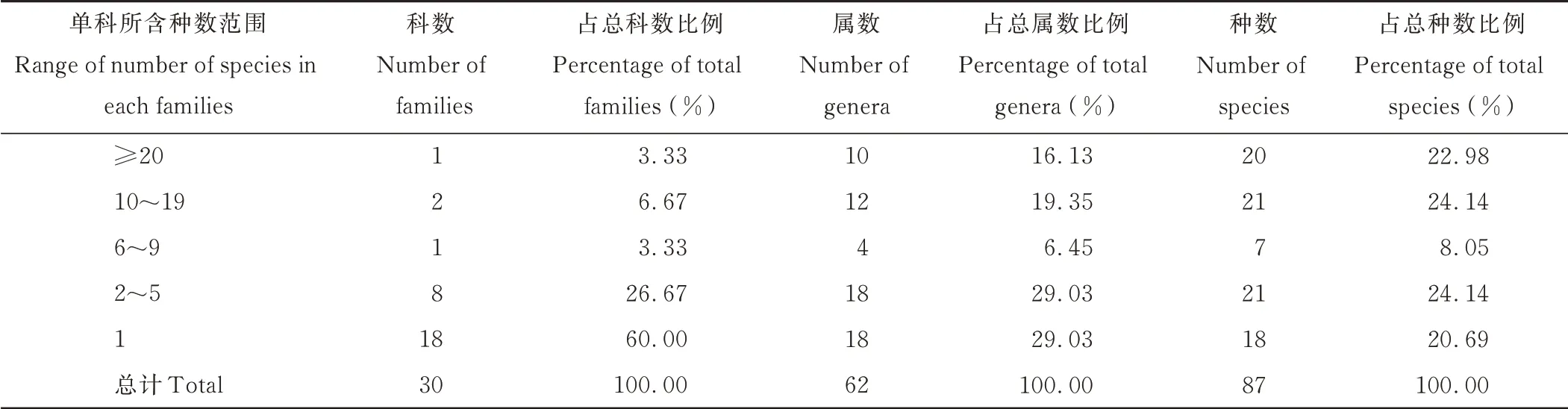
表2 贺兰山岩羊主要活动区植物群落物种数量Table 2 Species number of plant community in main active areas of P.nayaur in Helan Mountains
在贺兰山岩羊主要活动区植物群落的物种中,包括一年生草本、多年生草本、半灌木、灌木、乔木和木质藤本6 种生活型的植物。其中,草本63 种,一年生草本4 种,多年生草本59 种;木本23 种,灌木12 种,半灌木7 种,乔木4 种,藤本1 种,为木质藤本(图1)。
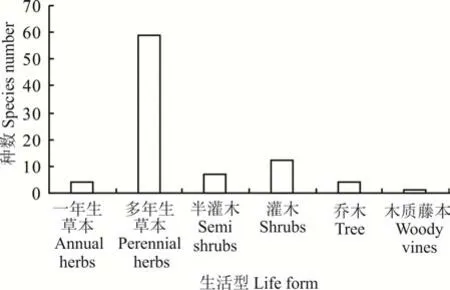
图1 贺兰山岩羊主要活动区植物生活型Fig. 1 Plant life forms of the main active areas of P. nayaur in Helan Mountains
2.2 贺兰山岩羊主要活动区不同海拔的植物群落组成与特征
不同海拔植物群落组成各有差异。海拔在1100~1500 m 的研究区科数最少,与其他海拔研究区的科数具有显著差异(P<0.05),而其他海拔研究区之间不存在显著差异(P>0.05);在属水平上,2500 m以上海拔的研究区属数最多,和其他海拔研究区之间存在显著差异(P<0.05),最少的是1100~1500 m 海拔处的研究区,与其他海拔研究区的属数具有显著差异(P<0.05);在种水平上,海拔在2500 m 以上的研究区植物种数最多,和其他海拔研究区之间存在显著差异(P<0.05),最少的是1100~1500 m 海拔处的研究区,与其他海拔的种数具有显著差异(P<0.05)(表3)。
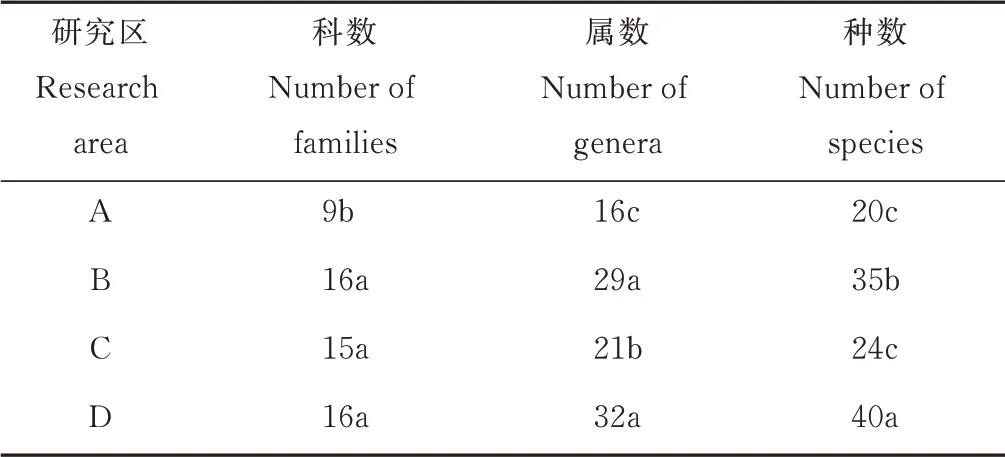
表3 贺兰山岩羊主要活动区不同海拔植物所含科、属、种的数量Table 3 Number of families,genera,and species of plants at different altitudes in main active areas of P. nayaur in Helan Mountains
在1100~1500 m 的研究区,主要植被类型为荒漠草原,共有9 科16 属20 种植物,其中以禾本科为主,有7 种,其次为苋科、旋花科、豆科、菊科等,主要属有针茅属(Stipa)、旋花属(Convolvulus)、隐子草属(Cleistogenes)和碱猪毛菜属(Salsola)等;海拔在1500~2000 m 的山地疏林草原,共有16 科29 属35 种,其中禾本科最多,有8 种,其次为蔷薇科(7 种)、菊科和豆科(各4 种),主要属有针茅属、早熟禾属(Poa)、绣线菊属(Spiraea)、桃属(Amygdalus)等;海 拔 在2000~2500 m 的 山 地 针 叶林,共有15 科21 属24 种植物,主要植物为禾本科(5种)、蔷薇科(3 种)和菊科(3 种),主要属为蒲公英属(Taraxacum)、披 碱 草 属(Elymus)和 鹅 观 草 属(Roegneria);海拔在2500 m 以上的高山−亚高山灌丛草甸中共有16 科32 属40 种植物,主要科为禾本科(7 种)、菊科和蔷薇科(各6 种)、石竹科(4 种)、豆科(3种)等,主要属有委陵菜属(Potentilla)、披碱草属、早熟禾属等(图2)。

图2 贺兰山岩羊主要活动区不同海拔植物群落各科所含种数Fig. 2 Species number of each family in plant communities at different altitudes in main active areas of P. nayaur in Helan Mountains
2.3 不同海拔梯度植被优势种及其重要值
贺兰山岩羊主要活动区植被类型沿海拔由低到高依次为荒漠草原、山地疏林草原、山地针叶林和高山−亚高山灌丛草甸,岩羊主要活动区植物物种重要值排序结果显示,贺兰山岩羊主要活动区植物优势种以禾本科草本植物和蔷薇科灌木植物为主(表4)。
在草本层,低海拔荒漠草原的优势种为短花针茅(Stipa breviflora)(重要值为0.332,下文括号中数值同为重要值)、无芒隐子草(Cleistogenes songorica)(0.200)等,中低海拔处的山地疏林草原优势种为针茅(Stipa capillata)(0.327),中高海拔处的山地针叶林的优势种为苔草(Carexspp.)(0.428)、真穗披碱草(Elymus gmelinii)(0.183)、小红菊(Chrysanthemum chanetii)(0.126)等,高海拔的高山−亚高山灌丛草甸的优势种为苔草(0.226)等(表4)。

表4 贺兰山岩羊主要活动区不同植物群落类型优势种及其重要值Table 4 Dominant species and their importance values of different plant community types in main active areas of P. nayaur in Helan Mountains
在灌木层,低海拔荒漠草原的优势种为刺旋花(Convolvulus tragacanthoides)(0.462)、松叶猪毛菜(Salsola laricifolia)(0.434)等,中低海拔山地疏林草原优势种为蒙古扁桃(Amygdalus mongolica)(0.407)、小叶金露梅(Potentilla parvifolia)(0.175)和猫头刺(Oxytropis aciphylla)(0.109)等,高海拔的高山−亚高山灌丛草甸的优势种为小叶金露梅(0.508)、银露梅(Potentilla glabra)(0.402)等,而中高海拔山地针叶林没有灌木植物生长。
2.4 贺兰山岩羊主要活动区不同海拔的植物物种多样性变化
贺兰山岩羊主要活动区植物群落多样性方差分析和多重比较结果显示,在群落水平上,草本层的物种丰富度、Shannon−Wiener 多样性指数和Simpson 优势度指数均大于灌木层,而均匀度指数小于灌木层。不同研究区之间的草本层植物的物种丰富度指数、Pielou 均匀度指数和Shannon−Wiener 多样性指数都存在显著差异(P<0.05),而Simpson 优势度指数之间没有显著差异(P>0.05);不同研究区之间灌木层的丰富度指数、Shannon−Wiener 多样性指数和Simpson 优势度指数都存在显著差异(P<0.05),而Pielou 均匀度指数没有显著差异(P>0.05)(表5)。
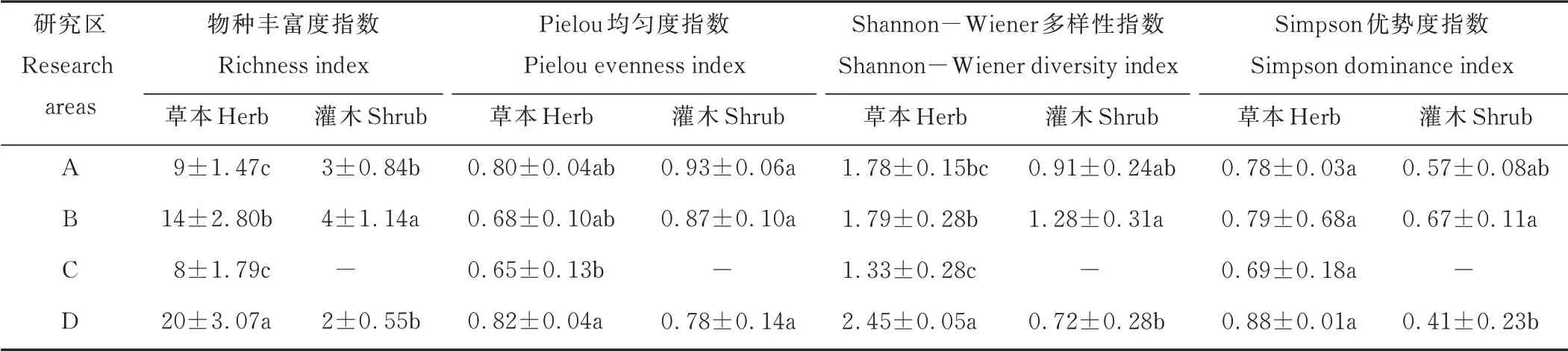
表5 贺兰山岩羊主要活动区植物群落多样性对海拔梯度的响应规律Table 5 Response of vegetation community diversity to altitudinal gradient in main active areas of P.nayaur in Helan Mountains
回归分析结果显示,草本层植物的物种丰富度随着海拔升高而升高;Pielou 均匀度指数、Shannon−Wiener 多样性指数、Simpson 优势度指数均与海拔高度相关不显著,呈现出先降低再升高的特征。灌木植物的物种丰富度随海拔升高而降低;Pielou 均匀度指数与海拔高度之间呈显著相关,随海拔升高而降低;Shannon−Wiener 多样性指数和Simpson 优势度指数均与海拔呈显著相关,随海拔的增加先升高后降低(图3)。
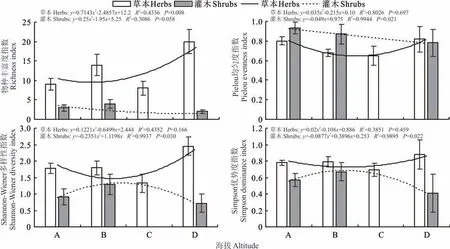
图3 植物物种多样性与海拔高度的关系Fig.3 Relationship between plant species diversity and altitude
草本层植物的物种丰富度、Pielou 均匀度指数、Shannon−Wiener 多样性指数和Simpson 优势度指数最高的均为海拔在2500 m 以上的高山−亚高山灌丛草甸,指数最低的均为海拔在2000~2500 m 的山地针叶林,不同海拔处的研究区草本植物物种多样性指数由高到低依次是高海拔区、中低海拔区、低海拔区和中高海拔区。
灌木层植物的物种丰富度、Shannon−Wiener 多样性指数和Simpson 优势度指数最高的均为海拔位于1500~2000 m 的山地疏林草原,而Pielou 均匀度指数最高的为位于低海拔处的荒漠草原;各个指数最低的均为2500 m 海拔以上的高山−亚高山灌丛草甸。不同海拔处的研究区灌木植物物种多样性指数由高到低依次是中低海拔区、低海拔区和高海拔区。
3 讨论
3.1 贺兰山岩羊主要活动区植物区系特征及其与岩羊等食草动物的关系
贺兰山位于蒙古高原南缘、黄土高原西北处,又与青藏高原东北部邻近,因此这里汇聚了蒙古高原、华北平原、青藏高原的植物成分,使得贺兰山的植物地理成分复杂,而具有明显的过渡性特征[23−24],又因其高度差较大、水热分布差异,从而形成了明显的垂直地带性植被[25]。根据相关研究,贺兰山西麓的植物物种以禾本科居多,菊科、豆科、蔷薇科等次之[26]。而本研究在贺兰山岩羊主要活动区植被调查共记录到87 种植物,隶属于30 科62 属,禾本科包含的植物种类最多,其次是蔷薇科、菊科和豆科等。岩羊主要活动区植物生活型组成有灌木、半灌木、多年生草本、一年生草本、木质藤本,这与胡天华[23]和段河等[26]在宁夏贺兰山国家级自然保护区的研究结果相符。
在贺兰山,荒漠草原以短花针茅为优势种,群落内植物多在10 种左右[25];山地疏林草原主要是灰榆、蒙古扁桃、短花针茅等小乔木和灌木稀疏分布[27];山地针叶林主要生长着油松(Pinus tabuliformis)和青海云杉(Picea crassifolia);海拔在2500 m 以上的高山−亚高山灌丛草甸植物种类繁多,草本植物主要有苔草、早熟禾及其他杂类草[26]。本研究关于岩羊主要活动区的植物区系特征印证了这些结果。
贺兰山岩羊所采食的植物以禾本科植物为主[28],还包括蔷薇科,豆科、菊科等植物,其中针茅是岩羊的大宗食物,早熟禾、灰榆、冰草等植物也是岩羊的主要采食植物[29]。本研究植物区系特征结果与岩羊主要采食的植物相符合,说明贺兰山岩羊在贺兰山能够获得生长发育所需的食物,同时,植物组成也可能是岩羊选择其活动区的关键因素。
有研究表明,即使食草动物的数量很少,也会是保护和恢复目标的障碍,因为它们对一些植物物种有着持续压力[30]。在贺兰山,岩羊数量持续增长,且分布并不均匀[31],其生境已经呈破碎化状态,部分生境内的植被因为过度啃食已经出现严重退化的现象,已经阻碍了植被的生长和恢复,对贺兰山植被的保护、生物多样性和群落结构的恢复及协调发展造成了巨大的威胁。
因此,植物群落组成及多样性与贺兰山岩羊等食草动物之间有着深刻的联系。食草动物对生态系统功能具有至关重要的作用,一些关键的生态系统过程受到它们的影响,如养分的周转和种子的传播[32−33]。而且它们还调控着植物的多样性和生产力[34],食草动物的捕食压力下,植被幼苗死亡率高,且其高度的生长率甚至可以忽略不计,从而导致植物成活率极低,而食草动物数量的减少会促使大多数植被的幼苗增长[35]。食草动物对植物群落的多样性和组成有着强烈的影响[36]。植物物种多样性会随着食草动物采食强度的增强而降低,而且在食草动物的采食行为中具有较强的选择性,这种行为则可能会加剧植物物种多样性的降低[37]。
同时,植物多样性对食草动物具有积极的调节作用[38]。植物多样性的增加通常会导致植物生物量和栖息地多样性的增加,这两者都有利于食草动物。植物多样性的增加可以通过提供更广的范围和更丰富的食物来促进食草动物数量的增加。食草动物对植物多样性的响应则反映了栖息地和营养效应之间的平衡状态[39]。
3.2 贺兰山不同海拔的岩羊主要活动区植物物种多样性的影响因素
能够对物种多样性产生影响的因素多种多样,如降水、温度、湿度、阳光、土壤条件、海拔和人为干扰等。本研究结果显示,海拔在2500 m 以上的高山−亚高山灌丛草甸中的草本层植物的物种丰富度和Pielou 均匀度指数都高于其他研究区,而2000~2500 m 海拔处的山地针叶林研究区的草本植物物种丰富度和Pielou 均匀度指数最低。这也与岩羊在高山草原活动较频繁[40]相印证。造成这种结果最主要的因素是水分、温度等气候因素[41−42],而海拔高度能够对水、热、湿度等条件产生直接决定作用[43]。宁夏贺兰山处于中温带干旱地区,这一气候区域使得热量和水分条件成了植物生长的限制因子,气候格局从而呈现出气温随海拔升高而降低,降水随海拔升高而增多[44]。在中低海拔区,相对来说温度高而降水量少,因此其蒸散量和干旱程度高于高海拔地区,从而这一区域的植物生长受到水分的限制作用,而在高海拔地区降水量相对增多,蒸散量相对减少,水分不再是限制植物生长的主导因素,加之人为干扰减少,因此物种丰富度逐渐增加[45]。高山−亚高山灌丛草甸研究区海拔位于2500 m 以上,降水量多,受到太阳直射时间较长,光照使得温度对植物的限制相对降低,因此这一研究区的草本植物物种多样性最高。山地针叶林研究区海拔处于2000~2500 m,降水量多,但这一研究区因高大乔木郁闭度较高,导致阳光不能直射至林下,空气湿度大,气温低,这可能是导致这一研究区草本层植物物种丰富度最低的原因,而该研究区没有灌木植物生长也与这一气候条件限制有关,灌木的生长喜光且不耐涝,这一地区林下既没有阳光直射,湿度又大,不利于灌木的生长。相反在中低海拔区,由于灰榆等小乔木生长稀疏,冠层较小,使太阳光能够照射至灌木层,又因这一区域温度较高导致蒸散量和干旱程度高,降水量相对较少,符合灌木的生长习性,因此在中低海拔区的灌木层植物的物种丰富度和Pielou 均匀度指数相对较高,这也与Jiang 等[4]的研究结果一致。但灌木也存在死亡率较高的现象[46],这是由于当种间和种内对于光照、水分、养分和空间等因素的竞争激化时,灌木的生长受到抑制所致。
已有研究表明,草本植物的Shannon−Wiener 多样性指数随海拔升高而增加[47]。本研究的结果与上述规律基本一致,除了山地针叶林研究区,其他研究区的Shannon−Wiener 多样性指数沿海拔梯度的变化印证了这一结果。其中海拔2500 m 以上的高山−亚高山灌丛草甸研究区与其他研究区之间的多样性指数存在显著差异,产生这种差异的原因主要是不同生境之间的差异使得各研究区生长的物种产生差别,进而使植物群落在结构和功能方面都产生了差异[48−49]。本研究通过对各研究区之间的Shannon−Wiener 多样性指数进行比较,发现山地针叶林研究区的Shannon−Wiener 多样性指数最低,Jiang 等[4]研究发现贺兰山以青海云杉(Picea crassifolia)为主的山地针叶林Shannon−Wiener 多样性指数最低,Liu 等[50]发现岩羊对山地针叶林植物的食用量低,而在本研究中的Shannon−Wiener 多样性指数最低的同样是以青海云杉为主的山地针叶林,这也说明了本研究与前人的结果一致。
Simpson 优势度指数和植物群落中的物种数量有关,优势度越大,植物群落不同种类的植物数量分布越不均匀。在本研究中,草本层植物的Simpson 优势度指数在0.69~0.88 之间,灌木层的优势度指数在0.34~0.67 之间,灌木层和草本层之间优势度的差异,可能是由于在岩羊主要活动区内灌木植物之间的种内和种间竞争激烈,从而导致灌木植物物种数量较低,进而使得灌木层优势度指数低于草本层。灌木在海拔2500 m 以上的高山−亚高山灌丛草甸研究区的优势度指数最小,可能是由于该研究区海拔高,人类活动少,但风力大,风蚀严重,不利于灌木的生长发育[16]。
物种多样性受到多因素的调节,而植物也能根据环境等因素有形态适应、物候适应、生理生态适应的多样性[10]。贺兰山的植物种类丰富、区系地理复杂、沿海拔垂直带变化多样共同造成了其物种多样性。
4 结论
根据植被调查结果,构成岩羊主要活动区的草本植物主要为禾本科植物,灌木以蔷薇科植物为主。海拔梯度对贺兰山岩羊主要活动区的植物物种多样性变化有一定影响,草本层植物物种多样性在高海拔地区达到峰值,而灌木植物由于其自身喜光但不耐涝的特性,在中海拔地区物种多样性最大。整体来看,草本层植物物种多样性随着海拔的升高在岩羊主要活动区呈现出先降低后升高的趋势,而灌木层植物物种丰富度、Pielou 均匀度指数呈现出沿海拔上升而降低的特征,Shannon−Wiener 多样性指数和Simpson 优势度指数呈现出先增加后降低的特征。因此,建议在不同海拔对贺兰山岩羊采取不同的管护措施,以达到防止植被退化和促进岩羊种群健康发展的效果。
