基于相对和绝对定量同位素标记技术分析2种成熟度桑叶对家蚕脂肪体蛋白质组的影响
2022-06-10贾漫丽孙志超李季生
谢 岩 王 晖 李 娜 贾漫丽 黄 露 孙志超 李季生
(承德医学院蚕业研究所,河北省高校特产蚕桑应用技术研发中心,承德067000)
家蚕(Bombyxmori)为寡食性昆虫,通过与其宿主桑树(MorusalbaL.)的自然选择和长期协同进化,对桑叶表现出偏好的采食性和桑叶营养成分消化吸收的适应性[1-3]。人工饲料可以克服桑树生长期对蚕业生产季节性的限制,通过人为调配营养成分来提高生产的标准化程度[4-5],现已成为蚕业科学发展的一个研究热点。但由于家蚕的摄食性和营养需求受到品种、地域、饲养环境的诸多影响,难以形成统一人工饲料用于大规模推广和全龄期饲用,而且全龄人工饲料饲育效果难以达到桑叶的饲育效果[6]。因此,桑叶仍然是家蚕的最适宜食物来源。桑叶营养成分含量直接影响家蚕的生长和经济性能[7-8],适宜的优质桑叶的选择是家蚕健康生长和蚕茧生产的保障。桑叶成熟度是桑叶营养物质变动的重要因素,其与家蚕生长和经济性状的关系研究较多[9-12],但鲜有深入揭示其分子机制的研究,难以为实际生产应用提供准确的理论指导。家蚕是重要的经济昆虫,也是基础生物学研究的优良模式动物[13-14]。全基因组计划的完成和遗传图谱的构建奠定了其在现代生命科学领域的优势地位,并促进了家蚕功能基因组学研究的发展。蛋白质组学是功能基因组学的重要组成部分,通过研究参与特定生理或试验处理状态的所有的蛋白质种类及其变化阐述生命活动机理,相对和绝对定量同位素标记(isobaric tags for relative and absolute quantitation,iTRAQ)技术因其高通量、高重现性、高灵敏性的特点成为蛋白质组定性和定量研究的有效工具,近年来被广泛应用于蛋白质功能注释、生物标记筛选、药物效应验证等生命科学领域[15-16],已在家蚕免疫、性别分化和滞育等机制的研究中发挥重要作用。
家蚕5龄食桑量占全龄食桑量的80%以上[17],该龄期饲喂不同成熟度桑叶,蚕体会发生明显差异[10,18-19],家蚕脂肪体具有类似哺乳动物肝脏和肾脏的功能,存在大量与合成、代谢、免疫和繁殖相关的蛋白[20-21],是研究家蚕对摄食物质响应的关键器官[22-23]。本试验在测定秋季老、嫩桑叶营养和活性物质含量的基础上,采用iTRAQ结合质谱检测技术高效捕获5龄蚕脂肪体对2种成熟度桑叶营养物质变动反应的差异表达蛋白,并应用生物信息方法对差异表达蛋白进行功能聚类和互作分析,以揭示桑叶成熟度与其营养物质含量的联系,摄食2种成熟度桑叶的家蚕脂肪体蛋白质组差异程度,蚕体摄食2种成熟度桑叶后其脂肪体细胞内差异表达蛋白的功能分类和表达模式,蚕体对摄食物质质量变动作出响应的关键蛋白,筛选出的桑叶主要差异营养物质和蚕体响应功能蛋白,为进一步揭示单一营养物质变动对蚕体生长和生产影响的关联机制奠定基础,以期丰富饲养家蚕营养组分的合理选择和调配的理论依据。
1 材料与方法
1.1 试验设计及样本采集
家蚕品种:菁松×皓月。1~4龄家蚕幼虫相同条件(25 ℃,同质叶片)下饲喂,5龄幼虫眠起,随机挑选200头分为2组(每组100头),测定总重并分别饲喂9月桑树老叶(春伐抽条基部4片叶)和嫩叶(春伐抽条顶部完全展开叶片4片),每天定量饲喂3次,次日清晨饲喂前(排出肠道粪便)称重并记录。
连续饲喂3 d后,解剖蚕体采集脂肪体组织,每个组设置2个重复,每个重复为6个个体(雌雄各占1/2)脂肪体混合样品,共计4个样品(C1、C2、T1、T2),C1、C2为老叶组(C组)样品,T1、T2为嫩叶组(T组)样品。
1.2 桑叶物质含量测定
饲蚕用老、嫩新鲜桑叶真空低温冻干,在45 ℃烘箱内烘至恒重,测定失水量,失水量和鲜叶重的比值作为水分含量值。冻干样品研磨成粉末,过80目筛。粗脂肪和粗蛋白质含量分别采用索氏抽提法和凯氏定氮法测定(青岛科创质量检测有限公司承担)。可溶性糖含量采用植物可溶性糖含量检测试剂盒(北京索莱宝科技有限公司,货号:BC0030)测定,还原性糖含量采用3,5-二硝基水杨酸(DNS)比色法测定,多糖含量采用蒽酮-硫酸比色法测定,总黄酮含量采用Al(NO3)3-NaNO2-NaOH比色法测定,总多酚含量采用福林酚比色法测定,上述指标均由紫外可见分光光度计(普析通用,中国)检测。1-脱氧野尻霉素(1-DNJ)含量采用高效液相色谱(HPLC)法,使用LC-2030高效液相色谱仪(岛津,日本)测定。除水分外,其余物质均采用测定物质重量和干物质重量的比值作为含量值。每种成熟度均采集3个生物学重复样品用于物质测定,数据用平均值±标准差(mean±SD)形式表示,含量数据用SPSS 19.0软件进行描述统计量计算和单因素方差分析。
1.3 家蚕样品总蛋白提取
取适量样品加入液氮研磨,采用三氯乙酸(TCA)/丙酮沉淀法去除杂质,预冷纯丙酮洗涤沉淀3~5次。裂解液[8 mol/L尿素、30 mmol/L 4-(2-羟乙基)-1-哌嗪乙磺酸(HEPES)、1 mmol/L苯甲基磺酰氟(PMSF)、2 mmol/L乙二胺四乙酸(EDTA)、10 mmol/L二硫苏糖醇(DTT)]重悬蛋白后,进行还原烷基化处理。再加入4倍体积的预冷丙酮,沉淀蛋白,用含0.1%(质量体积分数)十二烷基磺酸钠(SDS)的50%(质量体积分数)三乙醇胺(TEA)复溶后离心,保留上清。分别使用Bradford法和十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)对提取的蛋白进行定量和完整性检测。
1.4 家蚕样品总蛋白iTRAQ标记和液相色谱串联质谱(LC-MS/MS)定性和定量分析
样本总蛋白的iTRAQ标记与质谱分析委托华大基因科技有限公司进行。每个样品取100 μg,加入等量适宜的胰蛋白酶(trypsin)将蛋白酶解为肽段,进行标记,标记方法和步骤参照iTRAQ®Reagent-8 Plex Multiplex Kit(应用生物系统公司,美国)说明书,1个样品标记1种同位素标记,真空抽干标记肽段用于后续试验。
各标记肽段样品等量混合后通过Agilent 1100高效液相色谱仪(安捷伦,美国)和SCX 100强阳离子交换柱(飞诺美,美国)进行预分级,通过梯度洗脱分离30个组分,合并为16个,再使用Acclaim PepMap C18 100(赛默飞,美国)反相色谱除盐。脱盐组分依次通过纳升液相系统Dionex ultimate 3000 nano LC system(赛默飞,美国)的Strata-X C18 300色谱柱(艾杰尓,美国)进行梯度洗脱分离。质谱检测采用Q-Exactiv质谱仪(赛默飞,美国),检测参数:正离子模式,母离子扫描范围350~2 000 Da,二级分辨率17 500,离子源电压1 800 V,毛细管温度320 ℃,碎裂模式为高能碰撞解离(higher collision energy dissociation,HCD)。
将质谱扫描的原始文件输入到Proteome Discoverer 1.3软件(赛默飞,美国)进行提取和筛选质谱谱图[母离子质量范围:350~6 000 Da,二级质谱图中最小峰数:10,信噪比(S/N)域值:1.5]。提取后的谱图用Mascot 2.3.0软件在Bom_mori_uni数据库中进行蛋白质的定性和定量分析。
1.5 差异表达蛋白的筛选、功能富集分析和互作网络构建
筛选差异表达倍数(fold change,FC)>1.25或<0.80,即|log2FC|>0.33且P<0.05的蛋白进行功能信息注释、富集分析和互作分析。采用蛋白索引号(UniProtKB AC)在UniProt(http://www.uniprot.org)获取蛋白名称和基本功能,无信息的蛋白注释其氨基酸序列在AmiGO2(http://wiki.geneontology.org/index.php/AmiGO_2)和NCBI(http://www.ncbi.nlm.nih.gov/)中比对到最高同源序列(相似性大于80%)的信息;通过QuickGO(http://www.ebi.ac.uk/QuickGO/)和WEGO2.0(http://wego.genomics.org.cn)对差异表达蛋白进行GO注释和功能富集分析,通过KEGG(https://www.kegg.jp/)获取差异表达蛋白参与的代谢通路;使用STING(https://string-db.org/)和Cytoscape 3.9.1软件绘制差异表达蛋白互作关系网络图。
2 结果与分析
2.1 家蚕体重变化
分别以老、嫩桑叶定量饲喂100头5龄眠起蚕,每天调查1次蚕体总重和存活数,共投食3 d,测定数据4次(包括眠起采食前的测定,即第0天),其中老叶组家蚕4次平均体重分别为0.66、1.37、1.61、2.13 g,嫩叶组家蚕4次平均体重分别为0.65、1.38、1.72、2.37 g。如图1所示,饲喂不同成熟度叶片第2天开始,嫩叶组家蚕体重明显高于老叶组,并且在第3天差异趋势愈加明显。

图1 家蚕饲喂老、嫩桑叶的体重增长趋势
2.2 老、嫩桑叶物质含量差异分析
依次测定老、嫩桑叶营养成分含量:水分、粗蛋白质、可溶性糖、粗脂肪和还原性糖;活性物质含量:多糖、总黄酮、总多酚、1-DNJ。其中糖类均采用无水葡萄糖配制梯度浓度标准品,总黄酮、总多酚和1-DNJ分别采用芦丁、没食子酸、1-DNJ标准品配制适宜梯度浓度标准品,绘制标准曲线,各测定成分标准曲线线性程度良好,R2均高于0.99,结果见表1。除水分和多糖外,老、嫩桑叶的各物质含量均存在显著或极显著差异(P<0.05或P<0.01)。嫩桑叶的粗脂肪、粗蛋白质、还原性糖、总黄酮、总多酚、1-DNJ含量均高于老桑叶,可溶性糖含量则低于老桑叶。
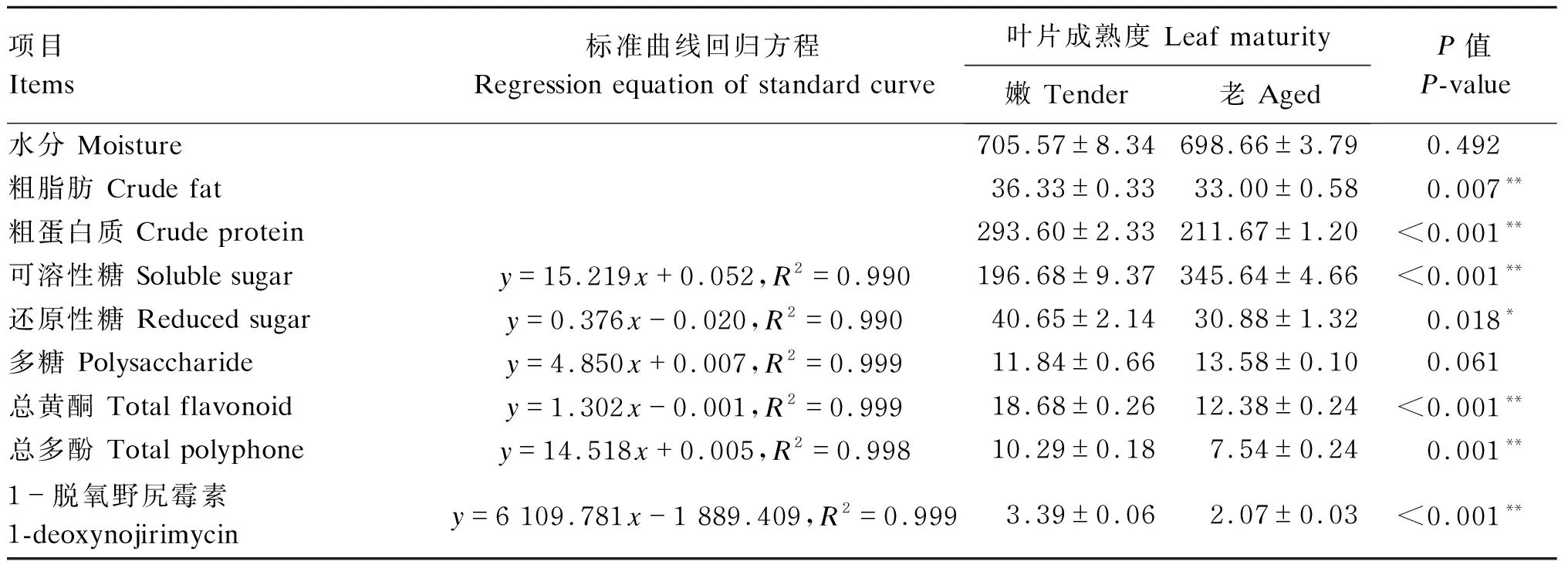
表1 桑叶物质含量
2.3 总蛋白提取质量
Bradford法绘制的标准曲线为y=0.183x-0.023,R2=0.993,提取的总蛋白浓度为1.09~1.52 μg/μL,总量为654~912 μg,取等量各样品进行SDS-PAGE,结果如图2,蛋白质电泳条带清晰完整,无降解。

1~4泳道分别为家蚕脂肪体样本C1、C2、T1、T2,泳道M为蛋白质marker(14~94 kDa)。
2.4 家蚕脂肪体蛋白质谱鉴定及定量结果
通过质谱扫描、PD筛选和Mascot 2.3.0匹配,共鉴定到:总谱图数344 774,匹配谱图数81 713,肽段种类数16 170和蛋白数2 172。设置FC>1.20或<0.83且P<0.05为筛选条件,共获得173个差异表达蛋白,见表2,相对于老叶组,嫩叶组上调表达蛋白为85个,下调表达蛋白为88个。
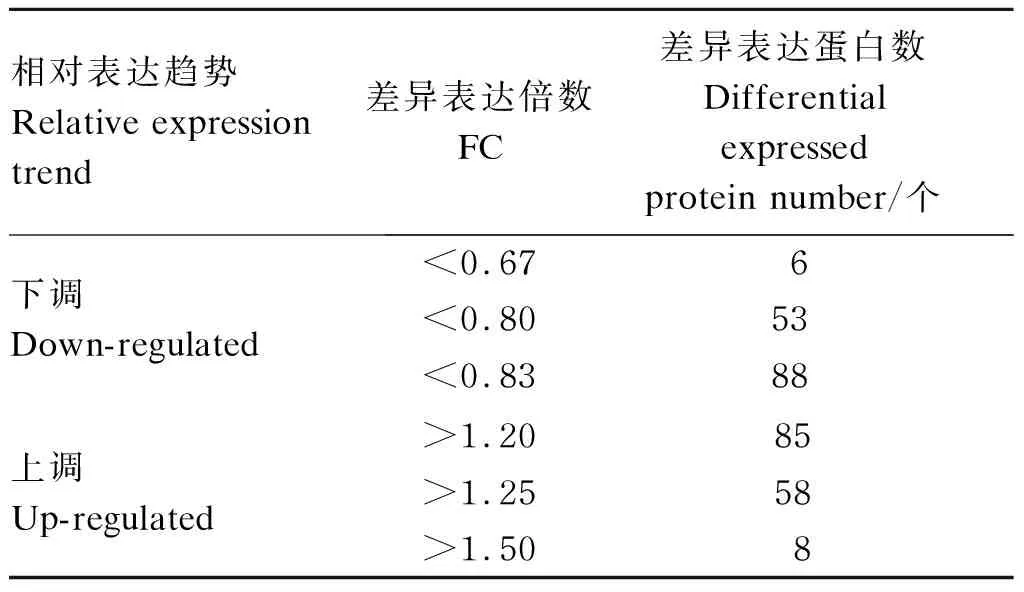
表2 家蚕脂肪体差异表达蛋白数(嫩叶组相对于老叶组)
2.5 家蚕脂肪体差异表达蛋白功能注释和聚类结果
选取FC>1.25或<0.80的58个上调蛋白和53个下调蛋白用于功能聚类分析,通过UNIPROTKB、NCBI和STRINGS等获取的综合信息对上述蛋白的基本信息和功能进行注释,除9个蛋白外,102个蛋白均能注释到蛋白名称、基因名称和功能概述中的至少1条信息。
差异表达蛋白在细胞组分、生物学过程和分子功能3个方面的GO分析,共注释到38个上调表达蛋白和43个下调表达蛋白,见图3。分析发现:1)差异表达蛋白参与9种细胞组分,细胞内组分多于细胞膜和细胞外区,组成细胞内细胞器的蛋白最多;2)差异表达蛋白参与14个分子功能,绑定、离子绑定、蛋白绑定、催化活性富集到的蛋白较多;3)差异表达蛋白参与16个生物学过程,参与细胞过程、代谢过程、有机物代谢过程、初级代谢过程、氮化合物代谢过程的蛋白数量明显高于其他过程,其次富集到蛋白较多的过程为生物过程的调节、发展过程、对刺激的反应、对应激的反应。

图3 家蚕脂肪体差异表达蛋白GO注释
111个差异表达蛋白中的53个通过KEGG的BlastKOALA得到通路注解,共有34个蛋白映射到84个三级通路,三级通路根据功能相似性归纳为30个二级通路,进一步概括为6个一级功能集合:代谢、遗传信息过程、环境信息过程、细胞过程、机体系统和人类疾病,见图4。其中代谢、人类疾病和机体系统注释到的蛋白和通路最多,分别为11个蛋白、38个通路,12个蛋白、20个通路和15个蛋白、13个通路。差异表达蛋白具体聚类到KEGG各个三级通路和功能信息见图5。

图4 家蚕脂肪体差异表达蛋白KEGG功能分类

图5 家蚕脂肪体差异表达蛋白KEGG通路注释
结合GO和KEGG分析富集到参与物质代谢、免疫、发育、繁殖、对刺激反应的功能相似蛋白集合,根据基因功能注释信息,进行汇总和分类统计,结果见图6。在功能聚类中按差异表达蛋白数量由多到少依次为:物质能量代谢、发育过程、对刺激的反应、免疫和感染类疾病、信号分子转导和互作、细胞死亡和循环、繁殖。同时还统计到一些结构和功能相似的家族蛋白,如表皮蛋白、低分子30K蛋白、核糖体蛋白、氧化或还原蛋白和热应激蛋白。注释的代谢相关蛋白有46个,占参与分析的差异表达蛋白总数(111个)的41.44%,包括已知的糖、脂和能量代谢蛋白:苹果酸脱氢酶1(MDH1)、ATP结合转运蛋白G超家族成员(ABCG)、酸性脂肪酶1(PGLIP1)、天门冬酰胺合成酶活性蛋白(ASNS)等。注释的发育相关蛋白有23个,占参与分析的差异表达蛋白总数的20.72%,包括表皮、肌肉、神经、寿命等组织发育相关蛋白:CPR4、CPR5、EDG91、CPR68、肌钙蛋白T(TNNT)、肌侵蛋白(MTPN)、肌球蛋白-2必需轻链(MLC-C)、脑星形细胞源性神经营养因子(MANF)、神经前体细胞表达发育下调蛋白8(NEDD8)、GLAZ/NLAZ[24]等。注释的刺激响应相关蛋白有23个,占参与分析的差异表达蛋白总数的20.72%,包括应激和刺激感受蛋白:热应激相关的热应激蛋白20.8(HSP20.8)和热应激蛋白20.4(HSP20.4)、氧化应激相关的γ-干扰素诱导溶酶体硫醇还原酶2(GILT2)和铜伴侣抗氧化剂1(ATOX1)、化学感受蛋白7(CSP7)等。注释的免疫、感染相关蛋白有12个,占参与分析的差异表达蛋白总数的10.81%,主要包括天然免疫相关蛋白和抗菌肽:蛋白质二硫键异构酶(ERP57)、钙网蛋白(CALR)、GILT2、巨噬细胞迁移抑制因子(MIF)、免疫诱导蛋白(B6TOX)、抗菌肽(ATTA)等。

图6 家蚕脂肪体差异表达蛋白功能注释
2.6 家蚕脂肪体差异表达蛋白的互作关系
111个差异表达蛋白通过STINGDE的多序列搜索(multiple sequences search)获得49个蛋白,组成78对蛋白质-蛋白质互作(protein-protein interaction,PPI)网络,该网络的PPI富集P值为7.61×10-10。将互作信息文件导入到Cytoscape中构建可视网络图和计算各节点(网络中各个蛋白)特征,结果如图7所示。通过MCODE插件的k核解析设定阈值为4获取2个子网络:7个节点[核糖体蛋白S17(RPS17)、核糖体蛋白L23(RPL23)、核糖体蛋白L4(RPL4)、核糖体蛋白L24(RPL24)、核糖体蛋白S28(RPS28)、信号识别颗粒蛋白19(SRP19)、NEDD8]的21对互作关系;4个节点[网格蛋白轻链(CLC)、促进海藻糖转运蛋白(TRET1)、脂蛋白受体1(LPR1)和脂蛋白受体2(LPR2)]的6对互作关系。通过CentiScape插件3个统计量——点度中心性(degree centrality)、接近中心性(closeness centrality)和中介中心性(betweenness centrality)对各节点中心度进行分析,获得12个高中心度的关键蛋白——脂肪酸绑定蛋白(FABP)、ERP57、钙网蛋白(CALR)、10 kDa热应激蛋白(HSP10)、ATP合酶亚基O(ATPsynO)、RPS17、RPL23、RPL4、RPL24、NEDD8、CLC、紫色酸性磷酸酶(PAP)。子网络分析和网络中心度分析均显示RPS17、RPL23、RPL4、RPL24和NEDD8处于蛋白网络中心,与其他蛋白互作关系多。
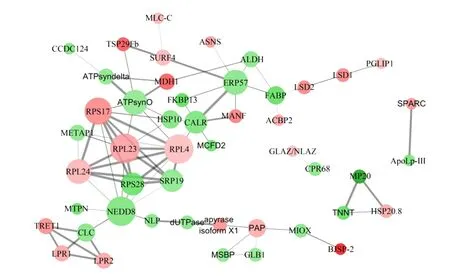
MLC-C:肌球蛋白-2必需轻链 myosin Ⅱ essential light chain;TSP29Fb:四次穿膜蛋白29Fb tetraspanin 29Fb;ASNS:天冬酰胺合成酶 asparagine synthetase;CCDC124:含有卷曲螺旋结构域蛋白124 coiled-coil domain containing 124;SURF4:过量位点蛋白4 surfeit locus protein 4 homolog;ALDH:醛脱氢酶 aldehyde dehydrogenase;ERP57:蛋白质二硫键异构酶 protein disulfide-isomerase;FABP:脂肪酸绑定蛋白 fatty acid binding protein;LSD1/2:脂质储存液滴表面结合蛋白1/2 lipid storage droplets surface-binding protein 1/2;PGLIP1:酸性脂肪酶1 acid lipase 1;ACBP2:乙酰辅酶A结合蛋白2 acyl-CoA binding protein 2;ATPsyndelta:ATP合酶delta亚基 ATP synthase delta subunit;ATPsynO:ATP合酶亚基O ATP synthase subunit O;MDH1:苹果酸脱氢酶1 malate dehydrogenase 1;MANF:中脑星形细胞来源的神经营养因子 mesencephalic astrocyte-derived neurotrophic factor;FKBP13:肽基脯氨酰异构酶 peptidylprolyl isomerase;RPS17:核糖体蛋白S17 ribosomal protein S17;HSP10:10 kDa热应激蛋白 10 kDa heat shock protein;CALR:钙网蛋白 calreticulin;MCFD2:多重凝血因子缺乏蛋白2 multiple coagulation factor deficiency protein 2;METAP1:甲硫氨酸胺基肽酶1 methionine aminopeptidase 1;RPL23:核糖体蛋白L23 ribosomal protein L23;RPL4:核糖体蛋白L4 ribosomal protein L4;RPL24:核糖体蛋白L24 ribosomal protein L24;RPS28:核糖体蛋白S28 ribosomal protein S28;SRP19:信号识别颗粒蛋白19 signal recognition particle protein 19;MTPN:肌侵蛋白 myotrophin;NEDD8:神经前体细胞表达发育下调蛋白8 neural precursor cell-expressed developmentally down-regulated protein 8;NLP:核质蛋白类似蛋白 nucleoplasmin-like protein;dUTPase:脱氧尿苷三磷酸酶 deoxyuridine triphosphatase; apyrase isoform X1:三磷酸腺苷同型X1;PAP:紫色酸性磷酸酶 purple acid phosphatase;MIOX:肌醇加氧酶 inositol oxygenase;BJSP-2:基础幼年激素抑制蛋白2 basic juvenile hormone-suppressible protein 2;TRET1:促进海藻糖转运蛋白1 facilitated trehalose transporter 1;CLC:网格蛋白轻链 calreticulin;LPR1/2:脂蛋白受体1/2 lipophorin receptor 1/2;MSBP:膜类固醇绑定蛋白 membrane steroid binding protein;GLB1:β-半乳糖苷酶1 beta-galactosidase 1;GLAZ/NLAZ:载脂蛋白D同源蛋白Glial Lazarillo/Neural Lazarillo apolipoprotein D homolog protein Glial Lazarillo/Neural Lazarillo;CPR68:假定表皮蛋白68 putative cuticle protein 68;MP20:肌肉蛋白20 muscle protein 20;TNNT:肌钙蛋白T troponin T;HSP20.8:热应激蛋白20.8 heat shock protein 20.8;SPARC:分泌蛋白、酸性、富含半胱氨酸 secreted protein, acidic, cysteine-rich;ApoLp-Ⅲ:载脂蛋白III apolipophorin Ⅲ。
3 讨 论
3.1 老、嫩桑叶物质含量差异与家蚕体重增长的联系
为了明确食入组分变化和家蚕体重增加的联系,我们测定了老、嫩桑叶的主要营养成分和活性物质含量。在营养成分上,嫩桑叶中粗脂肪、粗蛋白质含量高于老桑叶,尤其是粗蛋白质。李瑞等[18]采用秋季不同成熟度叶片饲喂5龄蚕,检测到嫩桑叶粗蛋白质含量高于老桑叶,盛食期蚕体重与饲喂桑叶的成熟度呈显著负相关。Alebiosu等[10]用不同成熟度(嫩、成熟、老、混合)桑叶饲喂2个品种家蚕,分析确定嫩桑叶的粗蛋白质含量显著高于其他成熟度叶片,粗蛋白质含量与幼虫体重显著正相关。这些研究结果与我们测定的物质含量和家蚕体重变化趋势一致。此外,我们测定出老桑叶中可溶性糖的含量显著高于嫩桑叶,但还原性糖含量则相反,与吕顺霖等[25]发现单糖在嫩桑叶中含量高于老桑叶结果一致。可溶性总糖包括单糖、双糖、寡聚糖、部分多糖,除蔗糖外,水溶性单糖和双糖都有还原性,为可溶性还原糖。单糖和双糖更易于5龄家蚕吸收,吸收效率在92%~94%,并且单糖>双糖[25],可为细胞生长快速供能。结合本试验结果与相关研究报道表明,桑叶成熟度显著影响其粗蛋白质和低分子糖类含量,是家蚕体重增长的原因之一。另外,给家蚕添饲适量的药用植物提取物或特定黄酮、多酚物质会提高家蚕的体重和寿命[26-29],嫩桑叶本身黄酮、多酚、1-DNJ等活性物质的含量高也是家蚕体重增加的重要因素。因此,秋季嫩桑叶的物质含量特征促进壮蚕期家蚕增重。
3.2 摄食2种成熟度桑叶的家蚕脂肪体差异表达蛋白功能聚类
综合以往的饲养研究来看,大蚕期建议饲喂适熟桑叶(第6~7叶位叶片或生长期为30~35 d的叶片),而嫩桑叶或老桑叶饲喂蚕在发育速度、抗逆性、存活率、茧量、茧重量、茧丝率、产卵量和良卵率等部分指标弱于适熟桑叶的饲喂效果[10,18-19,30]。相对于摄食适熟桑叶的蚕,一般摄食嫩桑叶的蚕体重和蚕茧生产性能高,而健康程度和存活率低,原因为:嫩桑叶相对蛋白质含量高而糖含量低使蚕体氨累积而中毒,嫩桑叶含水多干物质少使得蚕摄食营养不足[18,31]。相对于摄食适熟桑叶的蚕,摄食老桑叶的蚕则发育、抗逆性低,产卵量和良卵质量高,蚕茧生产性能低,原因为:老桑叶含水少而粗纤维含量高导致蚕体消化利用率低,总糖含量高而蛋白质含量低导致蚕体糖供给充足而蛋白质供给短缺[16]。因此,桑叶的成熟度与其物质含量差异密切相关,从而影响家蚕发育、抗病、茧质和繁殖等多方面性能。
在本研究中,食入秋季嫩桑叶的家蚕脂肪体相对于摄入老桑叶的蚕:1)细胞周期和凋亡相关蛋白死亡相关蛋白1(DAP1)[32]、硫磺内酯(ENDOS)[33]、NEDD8[34]、含有卷曲螺旋(coiled-coil)结构域蛋白124(CCDC124)[35]、MIF[36]下调,而细胞增殖促进蛋白ASNS[37]、富含天冬氨酸的TNF同源物(TNFSF13)[38]上调,由此可见,摄入适宜营养物质后蚕体通过抑制细胞凋亡、调节细胞周期和促进细胞增殖增加细胞体积和数量导致体重增长;2)营养储存蛋白TRET1、低分子30K蛋白、载脂蛋白、贮存蛋白1(SP1)、芳基蛋白质亚基β、碱性保幼激素抑制蛋白-2(BJSP-2)均上调[39-41],家蚕蛹化后不再进食,其所需物质能量均来源于幼虫,营养物质的增加储备会影响家蚕的产丝和产卵成绩;3)氧化应激蛋白肌醇加氧酶(MIOX)[42]下调,抗氧化应激相关蛋白ERP57[43]、醛脱氢酶(ALDH)[44]、ATOX1[45]下调,抗氧化应激保护性蛋白HSP20.4、HSP20.8[46]、GLAZ/NLAZ[47]、ATP结合盒转运蛋白G家族成员(ABCG)[48]上调,动物体的生命活动都涉及到物质和能量利用,嫩叶组家蚕氧化应激水平低于老叶组,免疫水平也是如此,说明机体应激和免疫消耗降低,可以将更多物质和能量用于生长发育。综上所述,家蚕对短期(3 d)食入的不同成熟度的桑树叶片在脂肪体内会做出复杂和综合性反应,可发掘出一批与家蚕体与摄入桑叶物质成分变化联系的候选蛋白,可为发掘有利于家蚕生长、经济性状改善的物质和分子标记奠定基础。
3.3 家蚕体对营养物质变动的关键响应蛋白挖掘
通过蛋白质互作中的子网络分析和网络中心度分析,显示NEDD8和4个核糖体蛋白RPS17、RPL23、RPL4、RPL24与其他蛋白的联系度高,说明它们处在蚕体对营养物质变化响应上的核心位置。
核糖体蛋白组成核糖体,在细胞内与RNA结合行使蛋白质翻译合成功能,此外,核糖体蛋白展示出多样的核糖体外功能[49]。本研究共发现11个核糖体蛋白——RPS17、RPS18、RPS21、RPS28、RPS30、RPL4、RPL7Ae、RPL6、RPL23、RPL24、RPL34在嫩叶组和老叶组间存在显著差异,除了RPS28、RPS30、RPL7Ae在嫩叶组下调外,其余在嫩叶组均上调。核糖体蛋白-MDM2-P53通路在感应细胞应激、整合营养压力方面起关键作用[50-51]。摄入营养的质量改变核糖体生物发生,引起核糖体蛋白作为营养变动的二级信使,通过与小鼠双微粒体2(MDM2)结合使细胞发生物质和能量代谢的适应性变动[52]。已发现14个核糖体蛋白可以绑定到MDM2酸域(central acidic domain,CAD),负向调控MDM2对P53蛋白泛素化,起到活化或稳定P53蛋白的作用[53]。真核生物存在80种基本核糖体蛋白,不断有新的核糖体蛋白与MDM2-P53互作被证实,如RPL4[54]、RPL6[55]和RPL34[56]。P53是总新陈代谢分子网络中的重要节点[57-58],可以在非静态或低胁迫条件下改变细胞代谢程序[59],在本研究中,饲喂不同成熟度的桑叶也是一种温和的、持续的营养物质差异刺激,PRL23、RPL4、RPL6、RPL34的蛋白量变化与组间处理的联系,说明它们可能通过核糖体蛋白-MDM2-P53通路发挥代谢调控作用。非单一的核糖体蛋白变动可能是由于不同的刺激诱导特定核糖体蛋白与MDM2结合,核糖体蛋白需要形成多聚复合物才能有效调节MDM2,核糖体蛋白之间存在互作等原因[53]。
NEDD8是类泛素蛋白的一种,通过类泛素化调控靶定基因发挥生物学作用。在本研究中,NEDD8在摄取嫩叶组下调。NEDD8和泛素蛋白参与脂肪细胞分化和脂肪代谢[34,60]。Park等[63]发现分化中的3T3-L1脂肪细胞,过氧化物酶体增殖物激活受体γ(peroxisome proliferator-activated receptor γ,PPARγ)(机体物质能量代谢的重要调控因子)[61-62]的含量正向依赖于NEDD8的表达量,NEDD8类泛素化通过与泛素化竞争抑制PPARγ2降解,从而调控PPARγ2靶定基因的转录。于洁[64]在2017年克隆到家蚕NEDD8基因,在家蚕BmN细胞中筛选和鉴定到28个NEDD8修饰底物蛋白,其中24%的蛋白富集到代谢功能,5个蛋白注释到氨基酸代谢通路,因此,家蚕NEDD8蛋白量变化可从调节氨基酸和脂肪合成或代谢蛋白质的功能和效应方面调控蚕体物质代谢。
本研究和相关文献初步支持核糖体蛋白-MDM2-P53通路蛋白和类泛素蛋白NEDD8在蚕体对持续的、温和的营养物质质量差异响应上敏感,是研究营养物质变动和动物机体变化机制的切入点,具体摄入营养物质含量差异和特异蛋白的确定联系、响应机制和效应通路有待于进一步发掘和验证。
4 结 论
本研究通过分析秋季老、嫩2种成熟度的桑叶基本物质含量差异和短期(3 d)采食它们的5龄家蚕脂肪体总体蛋白质相对变化水平,揭示了家蚕体重增长与桑叶内物质组成的联系和作用机制,发现:秋季嫩桑叶中粗蛋白质、低分子糖、总黄酮、总多酚、1-DNJ含量高,促进蚕体增重;摄食2种成熟度桑叶的家蚕脂肪体细胞共鉴定出173个差异表达蛋白,功能涉及代谢、发育、健康、抗性、生产和繁殖等生理作用;差异表达蛋白互作网络分析筛选出NEDD8和核糖体蛋白是蚕体对营养物质变动响应的关键候选蛋白。这些发现为进一步探索营养物质配比对家蚕生长发育的调节和控制的分子机制提供了数据。
