植物源miRNA跨界调控动物机体健康研究进展
2022-06-10彭永佳
黎 梦 彭 涵 彭永佳 张 瑾
(嘉兴学院生物与化学工程学院,嘉兴314001)
在维持动物健康和高效生产过程中,关注点往往集中于食物中传统的营养素及其他对动物营养调控基因表达的调节。植物性食物中还含一种丰富的功能性分子——miRNA,约含22个核苷酸(nt)的单链非编码的小RNA。最早发现miRNA可在动植物等体内细胞转录与翻译蛋白质的mRNA转录物反向互补,通过改变mRNA稳定性及其翻译进程等抑制mRNA的翻译,从而调节动植物的生理活动[1]。近年来研究表明,miRNA的调节作用不仅仅局限于细胞内水平,还可以在细胞间进行传递[2],甚至在物种间发生互作[3-4]。Zhang等[4]报道,食物中的大米miR168a可以被吸收进入小鼠的肝脏和血液中,抑制肝脏的低密度脂蛋白受体连接蛋白1(low density lipoprotein receptor adaptor protein 1,LDLRAP1)的表达,从而减缓肝脏对血液中低密度脂蛋白(low density lipoprotein,LDL)的清除。此后相继有研究证实,植物源miRNA可以通过饮食进入机体并发挥调控功能[5]。由此看来,这些植物源miRNA可能是一种新型的食物功能成分。在畜牧生产上,植物源miRNA通过饲料发挥跨物种调节作用的研究进展,有利于中草药或者植物饲料的开发利用,对畜禽生长发育和健康至关重要。表1中总结了主要的植物源miRNA跨界调控研究,这些研究结果可为理解植物源miRNA如何吸收转运并发挥跨界调控功能提供参考。本文从植物源miRNA跨界调节动物健康的稳定性、转运方式、吸收规律、作用机制和代谢等方面进行综述,这些研究具有重要的参考价值,为植物源miRNA参与调控动物健康以及疾病发生的机制提供试验依据,其对疾病的治疗及其在畜牧业中的应用具有重要意义。
1 植物源miRNA的稳定性
植物源miRNA作为一个重要的功能性核酸小分子,它通过采食进入机体发挥调节作用的前提是保持完整的结构和功能,这需要它具有一定的稳定性。miRNA作为短的单链核酸片段,最初被认为是极不稳定的。然而,自首次报道在人和小鼠的肝脏和血液组织中检测到大米miR168a以来[4],又陆续有研究报道在人和动物体内检测到植物源miRNA。Zhang等[4]研究证明,小鼠肝脏及大米miRNA在体外模拟的胃肠道环境中(pH 2.0,37 ℃)至少可以稳定保存6 h,且相对于动物的miRNA或者未甲基化的植物miRNA,大米的各种miRNA降解更加缓慢。这意味着植物源miRNA可抵抗胃肠道的酸碱环境和体内各种酶的降解,通过饮食进入机体组织发挥作用。此外,植物源miRNA还可以抵抗高温和强氧化作用。miR2911在经高温煎煮的金银花汤剂中仍然保持完整,并可通过饮用抑制流感病毒的复制,减轻小鼠流感症状[15]。与动物源miRNA相比,经过高碘酸盐氧化后植物源miR168a和miR156只有少量被破坏[4]。这种稳定性可能与miRNA存在形式和分子修饰有关。研究发现,miRNA经采食进入机体后由外泌体包裹并释放到循环中后,进入肝脏等目标组织与细胞中的Argonaute2蛋白(AGO2)结合,促进其与靶基因结合形成RNA诱导沉默复合物发挥功能作用[4]。植物中的miRNA还可能以植物外泌体样纳米颗粒(exosome-like nanoparticles,EPDENs)被肠道干细胞或者巨噬细胞吸收[16,23-24]。血液中游离的miRNA可以和高密度脂蛋白(high density lipoprotein,HDL)结合转运到受体细胞[25]。miRNA由外泌体包裹或与蛋白质结合等可以保护其不被降解。此外,植物源miRNA 3′端碱基戊糖2位O存在甲基化修饰,可防止miRNA氧化[4]或保护miRNA不被尿苷化,在一定程度上抵抗外源酶的降解作用[26],增强其稳定性。植物源miRNA的甲基化还可抑制转移酶和聚合酶的活性,保护miRNA不受核酸外切酶的降解[27];或抑制植物源miRNA以自身为引物并以mRNA为模板合成更多的miRNA长序列,保持其完整性和发挥调控作用的功能[28]。植物源miRNA对强酸、强碱、高温和体内多种酶等极端条件的抵抗作用,有利于其进入动物机体并在体内转运以及发挥调节作用。

表1 植物源miRNA跨界调控研究
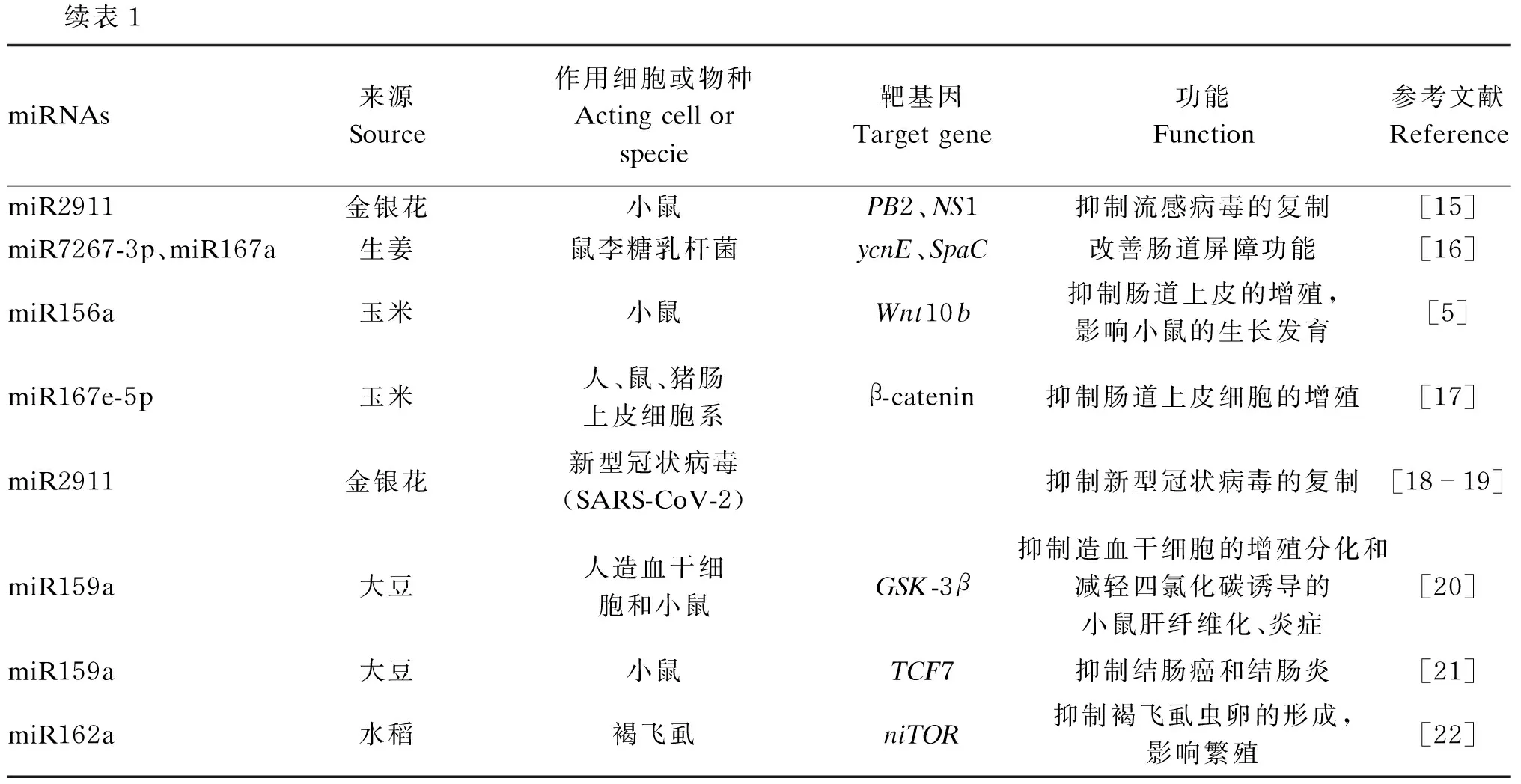
续表1miRNAs来源Source作用细胞或物种Acting cell or specie靶基因Target gene功能Function参考文献ReferencemiR2911金银花小鼠PB2、NS1抑制流感病毒的复制[15]miR7267-3p、miR167a生姜鼠李糖乳杆菌ycnE、SpaC改善肠道屏障功能[16]miR156a玉米小鼠Wnt10b抑制肠道上皮的增殖,影响小鼠的生长发育[5]miR167e-5p玉米人、鼠、猪肠上皮细胞系β-catenin抑制肠道上皮细胞的增殖[17]miR2911金银花新型冠状病毒(SARS-CoV-2)抑制新型冠状病毒的复制[18-19]miR159a大豆人造血干细胞和小鼠GSK-3β抑制造血干细胞的增殖分化和减轻四氯化碳诱导的小鼠肝纤维化、炎症[20]miR159a大豆小鼠TCF7抑制结肠癌和结肠炎[21]miR162a水稻褐飞虱niTOR抑制褐飞虱虫卵的形成,影响繁殖[22]
2 植物源miRNA进入动物机体的转运方式
食物中植物源miRNA要进入动物机体细胞内发挥作用,需要经消化道细胞特定的识别或由特定的转运载体摄取后,由血液循环系统转运到达特定组织细胞完成调控基因的表达。
早前在线虫的研究中发现,外源RNA可以依赖转运蛋白载体系统性RNAi(systemic RNA interference defective,SID)-2蛋白通过线虫消化道[29],随后经各个组织的SID-1蛋白转运到周围组织[30]。在高等动物机体中,外源miRNA可能穿过肠道屏障进入血液和组织[9]。Chen等[31]研究发现,SID-1跨膜蛋白(SID1 transmembrane protein 1,SIDT1)缺陷的小鼠,体液中饲料来源的植物源miRNA含量明显减少,且口服植物来源miR2911对肝纤维化的延缓作用也在SIDT1缺陷小鼠中被消除。研究证实,植物源miRNA在进食后可通过机械消化释放到胃,胃酸可以促进成熟的miRNA被胃小凹细胞的SIDT1吸收,由外泌体包裹,再分泌到循环系统并输送到其他组织和器官,最终进入受体细胞的miRNA被释放,并以序列特异性互补的方式调控靶基因[31]。
进入血液循环的植物源miRNA可能由外泌体通过直接融合、网格蛋白/小窝蛋白依赖的胞吞途径、巨胞饮、吞噬及脂筏介导的内吞途径等不同方式被其他组织细胞摄取[32-33]。因此,植物源miRNA在组织的吸收会受到外泌体大小、受体细胞上的识别蛋白如小窝蛋白、网格蛋白等表达的影响。同时,酸性环境可以促进外泌体表面小窝蛋白-1表达,增强受体细胞对外泌体的摄取[34-35],进而影响植物源miRNA的组织分布。
3 植物源miRNA进入动物组织的吸收规律
植物源miRNA经过胃肠消化道吸收已被认为是正常的生理现象,并呈现一定的吸收动力学规律[36],但可能存在一定的选择偏向性。因为植物miRNA的丰度和种类与进入动物机体的含量和种类并不是一致的,同一个miRNA在不同动物组织中的含量也不相同。有研究报道,在猪的心脏、肝脏、脑、肺脏、脂肪、肌肉等12个组织中检测到玉米不同丰度的18个miRNA含量丰富,其中心脏中miRNA含量最高,脑和肺脏次之,胰脏最少;在脂肪组织中玉米中丰度miR164a-5p和miR162-5p及高丰度miR168a-5p和miR167g-3p含量最高[9]。随后,王宇豪[37]分别给育肥猪灌喂人工合成的玉米miR164a-5p、miR167e-5p和miR168a-5p,24 h后经检测发现,miR164a-5p在眼肌和肝脏中含量最高,背部皮下脂肪中含量次之;miR167e-5p在心脏中含量最高,背部皮下脂肪中含量最低;miR168a-5p在心脏中含量最高,其余组织中含量均较低。通过对植物源miRNA与拟南芥miRNA数据进行保守性分析和碱基含量分析发现,植物源miRNA的吸收规律可能与miRNA第1和第3个碱基位点存在特定碱基有关[38],但目前仍未有相关的研究证明。此外,植物源miRNA还可能依赖于进入机体后的存在形式以及摄取的组织和细胞种类。
4 植物源miRNA跨界调控作用机制
在动植物中,miRNA在进化上均高度保守,其通过与靶基因mRNA部分互补或完全互补的方式调节基因表达,但执行这种调控作用的机制却不尽相同。动物源miRNA大多通过种子序列与动物靶基因mRNA 3′非编码区(3′ untranslated region,3′ UTR)互补,抑制或降解靶基因;植物源miRNA则通过与自身靶基因mRNA的开放阅读框(open reading frames,ORF)以几乎完全互补的方式结合,对靶基因进行切割和降解[39]。
植物源miRNA在行使跨界调节功能的过程中,与动物源miRNA调节自身基因表达具有一定的相似性。例如,玉米的miR156a通过种子序列靶向连接分泌蛋白分子Wnt10b基因的3′ UTR,抑制Wnt10b的表达,从而抑制肠上皮细胞的增殖[5]。此外,植物源miRNA也可以靶向动物基因3′ UTR以外的序列,如miR168a与LDLRAP1外显子4高度结合,降低小鼠肝细胞LDLRAP1蛋白的表达水平,但不影响LDLRAP1 mRNA的表达水平[4]。植物源miRNA还能以增强依赖序列的方式,在受体细胞中发挥作用[40]。有证据表明,植物源miRNA可通过结合树突状细胞的Toll样受体3(Toll-like receptor 3,TLR3)来抑制炎症;甲基化和未甲基化的植物源miRNA均具有免疫调节作用,但甲基化的植物源miRNA作用更强[40]。因为植物源miRNA 3′末端的甲基化修饰会增加miRNA 3′端与AGO2降解复合物结构域PAZ(argonaute like subfamily,PAZ domain)的亲和力,使其更好与mRNA结合[41],从而使它在功能上更加活跃。这种机制也确保较低含量植物源miRNA进入动物机体后能稳定发挥调控作用。
5 植物源miRNA进入受体细胞后的代谢
目前,植物源miRNA跨物种调节动物靶基因表达的说法已逐渐得到认可,而这些miRNA在发挥作用后又是如何进行代谢的仍需要进一步研究。有报道推测,跨界进入受体细胞中的RNA分子可以随受体细胞生长而被稀释,从而使得功能逐步降低;有的RNA在发挥调节作用之前就被受体细胞识别和降解;跨界小RNA也可以形成次级小RNA并实现更复杂的功能;而跨界传递的mRNA或长链非编码RNA也可以通过被剪切成小RNA发挥作用[42]。如菟丝子在寄生于拟南芥过程中可通过吸收转运miRNA,导致宿主筛子分子阻断氨基端蛋白1(SEOR1)等基因mRNA分裂,次级产生siRNA,减少mRNA和miRNA的积累[14]。当动植物miRNA与靶基因完全匹配或只与3′端少量不匹配时,miRNA会发生进一步降解,以减少自身的积累[43]。
6 植物源miRNA对动物机体健康的影响
6.1 植物源miRNA对动物免疫功能的影响
随着miRNA研究的深入,越来越多的证据表明植物源miRNA可跨界参与调控人和动物健康及疾病的发生。其中,植物源miRNAs在慢性炎症相关疾病的预防中具有治疗性应用的潜力。体外研究表明,植物源的miRNA和sRNA可通过抑制T细胞的增殖来调节树突细胞对炎症因子的反应能力来抑制炎症;这些植物源miRNA发挥的免疫调节作用并非完全与序列相关,而是通过普遍的在TLR3上结合抑制Toll/IL-1受体结构域适配器(Toll/IL-1 receptor domain-containing adaptor,TRIF)信号通路活性调节炎症[40]。将植物sRNA提取物注射给试验性自身免疫性脑脊髓炎小鼠后,小鼠表现出不同的T细胞亚群干扰素-γ(interferon-γ,IFN-γ)和白细胞介素-17(interleukin-17,IL-17)含量减少,进一步证实了植物sRNA通过以不依赖调节性T细胞(regulatory T cells,Treg)的方式抑制辅助性T细胞1(T helper cell 1,Th1)和辅助性T细胞17(T helper cell 17,Th17)的反应和抑制树突状细胞迁移,从而缓解小鼠免疫性脑脊髓炎的发病严重程度[40]。在动物妊娠免疫调节中,母体胎盘来源的外泌体可以改善母体对持续妊娠的适应,并促进胎儿存活[44];李青芝[45]证实,植物源miRNA可以通过母体传递给胎儿,这些植物源miRNA很可能在其中发挥免疫调节作用调控母体代谢稳态及子代健康。这预示着植物源miRNA有潜力作为一种新型的饲料成分来改善动物的免疫力。
6.2 植物源miRNA对动物肠道健康的影响
早在2013年,Ju等[24]发现葡萄的EPDENs可以被小鼠肠道干细胞吸收诱导其增殖,从而缓解小鼠因葡聚糖硫酸钠(DSS)诱导的结肠炎。随后证实,包括葡萄的4种可食用植物中分离出EPDENs富含miRNA等物质,这些EPDENs可以通过口服被肠巨噬细胞和干细胞吸收;生姜中EPDENs优先诱导小鼠巨噬细胞表达抗氧化基因、血红素加氧酶-1和抗炎细胞因子中白细胞介素-10(interleukin-10,IL-10)等;葡萄柚、生姜和胡萝卜EPDENs则促进核因子的激活维持肠道稳态平衡;而这些EPDENs均能激活小鼠隐窝中肠道稳态相关的重要通路Wnt/转录因子(TCF)4转录机制,表明从各种水果和蔬菜中摄入包含miRNA的EPDENs有利于维持肠道内稳态[23]。Teng等[16]进一步研究发现,植物的EPDENs还可以被肠道微生物群吸收改变菌群组成,影响宿主的肠道健康,其中生姜中EPDENs的mdo-miR7267-3p可介导靶向鼠李糖杆菌单氧酶ycnE,使吲哚-3-羧醛(I3A)产量增加,以诱导肠道白细胞介素-22(interleukin-22,IL-22)的产生,改善由DSS诱导的肠上皮损伤,缓解小鼠结肠炎。本实验室研究发现,减少小鼠饲粮中的玉米sRNA,小鼠肠道的发育明显优于正常饲料饲喂的小鼠,体重明显增加;在猪肠道上皮细胞系证明植物源miR156a可以通过靶向Wnt10b基因抑制Wnt信号通路的活性,从而抑制肠道上皮细胞的增殖,并在小鼠活体上灌服合成的植物源miR156a进一步证实了这个结论[5]。此外,本实验室在体外试验研究证明,miR167e-5p可以通过靶向β-连环蛋白(β-catenin)抑制人、小鼠和猪肠道上皮细胞的增殖,而miR167e-5p的抑制作用在同时加入其抑制剂后被相应抵消[17]。小鼠强饲大豆miR159a和大豆RNA也能通过靶向TCF7抑制Wnt信号通路的活性抑制结肠癌和肠道炎症,抑制miR159a时这种效应消失[21]。植物源miRNA调节肠道菌群平衡和肠道健康的研究,对提高肠道对营养物质的消化吸收、保持畜禽的健康生长和提高畜禽生长性能具有指导意义。
6.3 植物源miRNA对相关疾病的影响
植物源miRNA在代谢综合征[4]、抑制病毒[15,18-19]、癌症[10,21]和心血管疾病[12]等方面的也有相应的研究。金银花miR2911可抑制甲型流感病毒(influenza a virus,IAV)包括H1N1、H5N1和H7N9亚型mRNA的表达和阻断蛋白翻译而抑制病毒的复制;口服富含miR2911可以防止H5N1病毒引起的IAV感染减少引起的小鼠死亡[15]。新型冠状病毒(SARS-CoV-2)感染者在口服金银花汤剂后,体内新型冠状病毒复制明显受到抑制;临床试验证明,口服富含miR2911的金银花汤剂志愿者7 d转阴率高达83.3%,口服无miR2911的传统中药混合物的志愿者7 d转阴率仅达26.1%,表明金银花miR2911可增加感染患者的治愈可能,防止病毒的传播扩散[19]。miR2911在埃博拉病毒和非洲猪瘟病毒等多个病毒的基因组上均存在相应的靶点,它可靶向埃博拉病毒编码的基质蛋白(matrix protein,VP40)和刺突糖蛋白(spike glycoprotein,GP)基因,降低GP蛋白的表达量从而来减少小鼠的感染死亡[46]。小鼠口服合成的miR159可抑制编码Wnt信号转录因子的TCF7抑制增殖,从而抑制小鼠异种乳腺肿瘤的生长抑制[10]。基于这些研究,解析植物源miRNA的吸收、调控和代谢过程,有助于解释中草药或植物提取物的分子机制,为各种疾病提供预防策略和更安全、更“自然”的治疗方法,为中草药在维持动物健康生长奠定基础。植物源miRNA对健康和疾病的潜在影响仍需要大量深入研究。
6.4 植物源miRNA对动物生长的影响
各种植物或者中草药已被广泛用于畜禽饲料中以提高养殖水平,追求健康高效畜禽生产。根据前期的研究表明,膳食中的植物源miRNA可作为新的功能成分发挥作用。有报道称,猪可以通过采食获取玉米中的miRNA,并在各个组织包括影响肉品质的关键组织——脂肪和肌肉检测到丰富的植物源miRNA[9]。本实验室通过给小鼠灌服植物源miRNA发现小鼠脂代谢显著增加,其中肌肉比例显著增加,附睾脂肪沉积显著减少[47]。根据前期研究提示,植物源miRNA很有可能通过采食参与肌肉和脂肪形成的调节。研究植物源与肉质形成有关的miRNA,对提高猪的瘦肉率和减少体内脂肪沉积、改善胴体品质和肉的风味具有重要意义。花粉中miR162a可以靶向果蝇和蜜蜂的果蝇雷帕霉素靶蛋白(Drosophilamelanogastertarget of rapamycin,dmTOR)抑制身体和卵巢的发育,延缓蜜蜂的发育[11];水稻miR162a可以抑制褐飞虱的褐飞虱雷帕霉素靶蛋白(Nilaparvatatarget of rapamycin,nlTOR),抑制褐飞虱虫卵的形成,影响繁殖[22];母体胎盘来源的外泌体在母子信息传递中起免疫调节作用[44],这些研究为提高畜禽繁殖力和产仔率提供参考。因此,植物源miRNAs在提高畜禽生产水平和产品品质等方面均具有重要价值,对畜牧业的发展起着重要的作用。
6.5 植物源miRNA在畜禽饲料作物中的应用
尽管自然界中天然存在的跨界抗病RNA干扰(RNA interference,RNAi)在近年来才被证明,但是RNAi技术已经被广泛应用于作物抗病育种中。近年来发现,黄萎病菌细胞可以吸收宿主棉花miR166和miR159,分别靶向性降解致病基因2个编码钙离子依赖半胱氨酸蛋白酶-1(cysteine protease-1,Clp-1)和异木霉菌素C15羟化酶(hydroxylase isotrichotin C15,hic15)的基因,减弱黄萎病菌致病性[48]。由此可确定新的宿主植物防御策略:植物通过特定输出miRNA来诱发对致病真菌的跨界基因沉默,并以此获得抗病能力。在此基础上,由Zhang等[49]成功建立了跨界RNAi介导的抗黄萎病棉花种植新技术系统。2018年Nature和Science杂志相继报道自然界中天然存在植物-真菌跨界抗病RNAi[13-14]。拟南芥细胞外泌体可携带sRNA被灰葡萄球菌摄取,其TAS1c-siR483和TAS2-siR453可以靶向的致病基因表达,减弱真菌致病性[13];诱导菟丝子通过吸器把miRNA传递给拟南芥,共同下调SEOR1和F-box/RNI样超家族蛋白(TIR1)等的表达,发挥毒力因子的作用[14]。随着RNAi和其他分子学领域的深入研究和应用,miRNA作为RNAi途径中重要的非编码小分子RNA,在饲料作物病虫防治、提高饲料作物的产量和营养价值等方面的作用将得到更广泛深入的关注。
7 小结与展望
植物源miRNA作为一类功能性小RNA分子,本质是核苷酸,但在跨物种机体中的吸收转运和功能调节机制过程中发挥独特作用。虽然在动物体内检测到的植物源miRNA可能是由污染[50-51]、技术人工制品[51]和确认偏差[52]等原因所造成,但越来越多的研究已证实植物源miRNA的确存在跨界调控功能。因此,在进行相关研究时,需要对外源miRNA检测方法、进入体内有效生物浓度、试验对象和膳食miRNA的来源等进行标准化评估。同时还应考虑植物源miRNA在跨物种机体中吸收、分布、代谢和消除等方面存在的变异性,以及不同食物摄入来源,均会导致血液或组织特定miRNA的浓度不同。植物源miRNA的吸收是一个持续累积的过程,除了miRNA的浓度外,发挥功能的程度还取决于miRNA与其靶位点的互补性以及活跃性。
综上所述,植物源miRNA的跨界调控存在着巨大的潜力和重要的研究意义,有必要进一步探究其对健康和疾病的潜在影响以及在畜牧、农业生产中的开发利用。相信随着科学研究不断推进和植物源miRNA跨界调控的深入探索,将会推动miRNA技术在疾病、农业、畜牧业生产等领域的应用。
