饥饿胁迫对福寿螺生长、抗氧化系统及生化物质的影响*
2022-06-10肖泽恒章家恩石兆基张春霞
肖泽恒,甘 甜,秦 钟,2**,章家恩,2**,石兆基,张春霞
(1.华南农业大学资源环境学院 广州 510642;2.广东省生态循环农业重点实验室/广东省现代生态农业与循环农业工程技术研究中心/农业部华南热带农业环境重点实验室 广州 510642)
在生物的生长过程中,由于季节和环境的变化,生物经常面临食物缺乏的情况。在饥饿条件下,生物会减缓生长发育,并消耗储能物质为机体提供能量以维持生命活动。与此同时,生物在饥饿情况下,消化腺会产生大量活性氧,当活性氧的生成速率超过其清除速率时,多余的活性氧会破坏细胞从而导致消化腺组织损伤,威胁生命,这种氧化应激也被认为是饥饿对生物造成负面影响的主要原因。因此,可通过测定生物的生长指标、抗氧化酶活性和生化物质含量来评价其健康状况及对环境的适应状况。
目前,有关福寿螺在饥饿胁迫下的生长和生化物质变化方面的研究已有部分报道: 韩微研究发现,持续的饥饿胁迫致使福寿螺中螺(4.5 ± 0.5) g 体内粗蛋白、粗脂肪、粗糖含量显著低于对照;董胜张等研究发现,间歇性饥饿胁迫下福寿螺的螺重和特定生长率显著低于持续喂食的福寿螺(<0.05)。对于福寿螺在饥饿条件下的抗氧化指标的变化研究则少见报道。本研究在已有工作的基础上,研究了持续饥饿胁迫下福寿螺的形态生长指标及总蛋白、糖原等生化物质的含量变化,同时测定了过氧化物酶活性、总超氧化物歧化酶活性、过氧化氢酶活性等抗氧化指标,以期揭示福寿螺应对饥饿胁迫的生理生态对策,同时为研究其入侵机制及防控策略提供参考。
1 材料与方法
1.1 试验材料
试验所用福寿螺于2020年8月底采集自华南农业大学增城基地同一稻田(113°4′E,23°1′N),为保证试验用螺饥饿程度一致,于华南农业大学土壤生态实验室进行8 d 的雌雄混合暂养,即采用塑料水族箱(45 cm × 35 cm × 35 cm)饲养,并将其放置在有空调的室内阳台,暂养期间温度控制在25 ℃,水深控制在15 cm,每天更换2/3 曝气除氯自来水和投喂足量生菜(L.var.Hort.)。选取表观正常、活力较强、体型较为一致的福寿螺个体用于试验,试验用螺的个体质量为(3.69 ± 0.22) g,壳高为(24.25 ± 0.33) mm。
1.2 试验处理
设置6 个试验组,其中3 个饥饿处理组(STA):饥饿10 d (STA10)、饥 饿20 d (STA20)、饥 饿30 d(STA30),同时设置3 个对照组(CON): 饱食10 d(CON10)、饱食20 d (CON20)、饱食30 d (CON30),每个试验组均设置5 个重复。试验开始前,通过轻微打开福寿螺厣甲检查阴茎鞘的存在与否或根据厣甲的形状来区分雄螺和雌螺,并随机选择5 只雌螺和5 只雄螺放入直径20 cm、高30 cm、容积为9.42 L 的柱状玻璃缸中,作为一个重复,共300 只福寿螺被用于试验。为避免福寿螺死亡带来的密度差异,每个处理组另设2 个重复用于替代试验中死亡的福寿螺,替代用螺120 只。试验开始时,将装有福寿螺的柱状玻璃缸置于25 ℃、14 L∶10 D 下的人工气候箱中,分别在10 d、20 d 和30 d 取出福寿螺进行指标测定。
1.3 日常管理
试验过程中保持水深为15 cm,每天上午用含有饱和碳酸钙的曝气除氯自来水进行换水,换水量为原水量的2/3。饥饿处理组不投喂任何食物,对照组每天早上9:00 投喂足量新鲜生菜并取出前一天未被吃完的生菜。
1.4 测定方法
在第10 d、第20 d、第30 d 随机选取饥饿组和对照组中的福寿螺各10 只(每个重复中取2 只),用毛巾擦干表面水分后,用精确到0.01 g 的天平称量体质量()。用精确到0.01 mm 的游标卡尺测量福寿螺的壳高、壳宽、壳口长和壳口宽。测量后的福寿螺被放回试验玻璃缸中。
两组治疗前FBG、2hPG、HbA1c间无显著差异,治疗后两组血糖相关指标均较治疗前显著降低(P<0.05),且观察组显著低于对照组(P<0.05),见表2。
在第10 d、第20 d、第30 d 随机选取饥饿组和对照组中的福寿螺各5 只(每个重复中随机取1 只,不分雌雄),解剖分离出消化腺组织,用液氮速冻后于-80 ℃冰箱保存。测时按1∶9 的质量体积比加入预冷的生理盐水,冰水浴中用电动高速匀浆器制成10%的组织匀浆液,匀浆液在4 ℃下3000 r·min离心10 min,取上清液用于过氧化物酶(POD)活性、总超氧化物歧化酶(SOD)活性、过氧化氢酶(CAT)活性、还原型谷胱甘肽(GSH)和丙二醛(MDA)含量的测定。
各类酶的活性均采用南京建成生物工程研究所提供的试剂盒进行测定,其中POD 活性采用催化过氧化氢法,定义为37 ℃条件下,每毫克组织蛋白每分钟催化1 μg 底物的酶量为一个酶活力单位。SOD活性采用黄嘌呤氧化酶法测定,其定义为37 ℃条件下,每毫克组织蛋白在1 mL 反应液中SOD 抑制率达50%时所对应的SOD 量为1 个活力单位(U)。CAT活性采用钼酸铵显色法测定,其定义为每毫克组织蛋白每秒钟分解1 μmol HO的量为1 个活力单位。GSH 含量测定采用DTNB 法,单位为mg·g(以prot计)。MDA 含量用TBA(硫代巴比妥酸)法测定,单位为nmol·mg(以prot 计)。
在第10 d、第20 d、第30 d 随机选取饥饿组和对照组中的福寿螺各5 只(每个重复中随机取1 只,不分雌雄)测定其自由水、结合水、甘油、脂肪、糖原和总蛋白的含量。自由水和结合水含量采用质量法测定,甘油含量采用徐淑等的方法测定,脂肪含量采用Folch 法测定;糖原含量采用蒽酮比色法测定;总蛋白含量测定采用考马斯亮蓝法,单位为g·L。
1.5 数据处理
运用Microsoft Excel 软件整理数据,采用SPSS 25.0 统计软件对数据进行独立样本检验,分析比较相同处理时间下的CON 组与STA 组的差异,<0.05代表有显著性差异。文中的数据均表示为平均值±SE。用R 语言的“ggplot2”程序包绘图。
2 结果与分析
2.1 饥饿胁迫对福寿螺生长发育的影响
如图1所示,在饥饿胁迫下,处理组福寿螺的体质量、壳高、壳宽、壳口长和壳口宽在10 d 时均未与对照组形成显著差异。但随时间延长至20 d,壳高和壳口长的生长受到抑制且显著低于对照(<0.05)。饥饿处理30 d 后,福寿螺各形体指标均显著低于对照组(<0.05)。

图1 饥饿胁迫对福寿螺生长的影响Fig.1 Effects of starvation stress on the growth of Pomacea canaliculata
2.2 饥饿胁迫对福寿螺消化腺抗氧化系统的影响
如图2所示,在饥饿胁迫10 d、20 d、30 d 时,处理组福寿螺POD 活性和GSH 含量与对照组均未形成显著差异。在10 d 时,饥饿处理下的福寿螺SOD活性与对照组无显著差异;20 d 和30 d 时,处理组SOD活性(10.97 U·mg和10.77 U·mg)显著高于对照组(10.11 U·mg和9.97 U·mg) (<0.05)。处理组福寿螺CAT 活性在10 d、20 d 时与对照组无显著差异;30 d 时处理组(38.13 U·mg)显著高于对照组(28.60 U·mg) (<0.05)。饥饿组MDA 含量在10 d、20 d时与对照组无显著差异,到30 d 时处理组(1.19 nmol·mg)显著高于对照组(0.93 nmol·mg) (<0.05)。
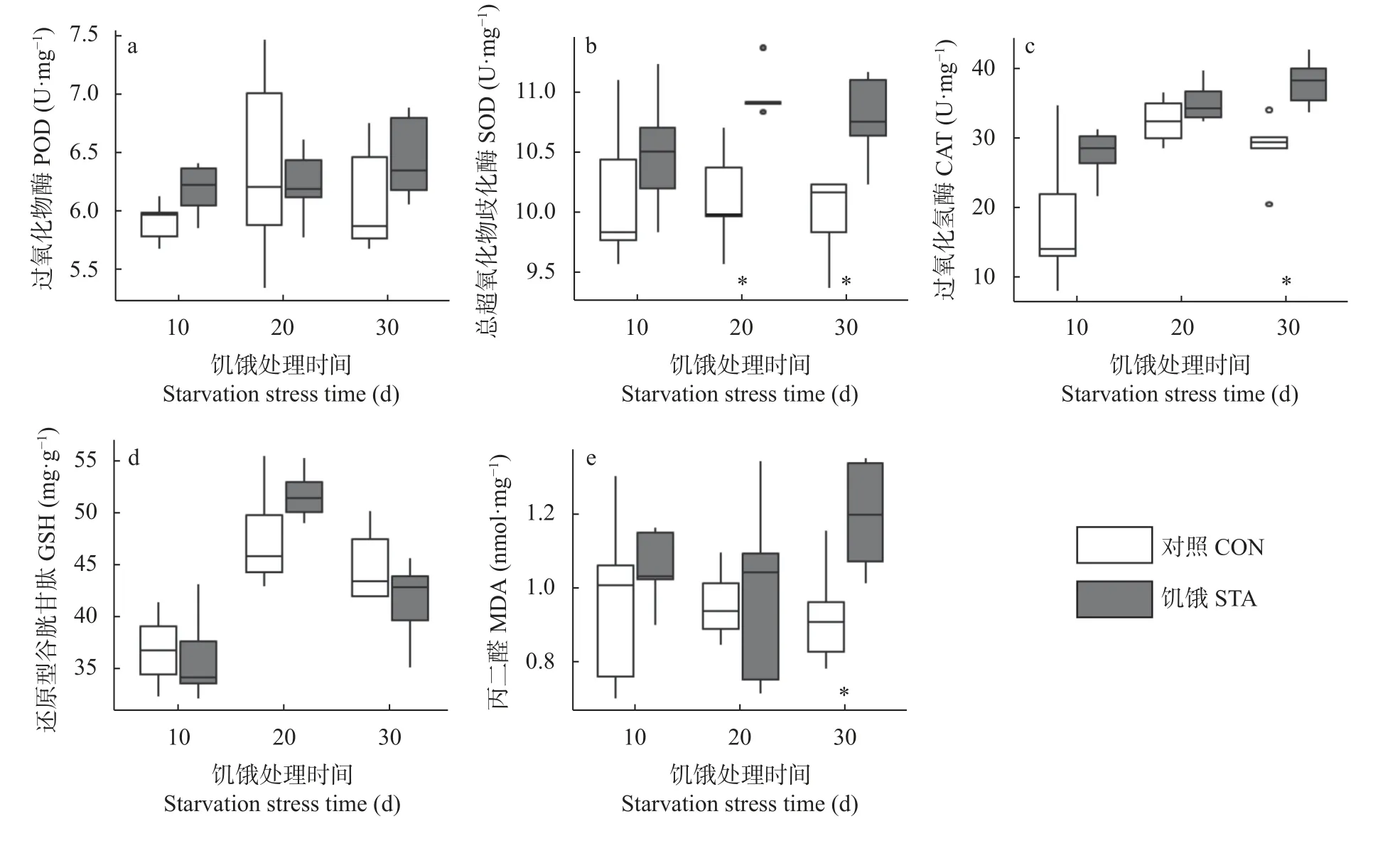
图2 饥饿胁迫对福寿螺抗氧化系统的影响Fig.2 Efects of starvation stress on antioxidant system of Pomacea canaliculata
2.3 饥饿胁迫对福寿螺生化物质含量的影响
饥饿胁迫对福寿螺生化指标的影响如图3所示。在前20 d,饥饿处理下的福寿螺总蛋白含量与对照组无显著差异,但在30 d 时饥饿处理组福寿螺总蛋白含量(85.91 g·L)显著低于对照组(93.64 g·L) (<0.05)。饥饿处理组福寿螺糖原含量在10 d 时与对照组无显著差异,在20 d 和30 d 时显著低于对照组(<0.05)。饥饿处理组福寿螺甘油含量在第10 d 显著高于对照组(<0.05),而在20 d 和30 d 时显著低于对照组(<0.05)。福寿螺脂肪含量在10 d、20 d 和30 d均显著低于对照组(<0.05),且随着饥饿胁迫时间的增加,与对照组的差异越来越显著,30 d 时处理组脂肪含量只有对照组的60.14%。饥饿处理下福寿螺自由水含量在10 d 时与对照组无显著差异,在20 d 和30 d 时处理组福寿螺自由水含量显著高于对照组(<0.05)。饥饿处理组福寿螺结合水含量在整个饥饿期间与对照无显著差异。

图3 饥饿胁迫对福寿螺生化物质含量的影响Fig.3 Effects of starvation stress on the contents of biochemical substances in Pomacea canaliculata
3 讨论
3.1 饥饿胁迫对福寿螺生长和含水量的影响
在饥饿条件下,水生生物无法有效从外界获取能量物质,只能通过消耗自身储能物质和减缓生长发育来维持生命活动,形体指标可以直观地反映生物自身的生长状况。福寿螺在饥饿胁迫30 d 后,体质量显著低于对照组(图1a),这与薛明等对方斑东风螺(Link)的研究结果一致。值得一提的是,试验过程中并没有观察到处理组福寿螺体质量的下降,原因可能是含水量的升高导致,如图3e 所示,处理组福寿螺自由水含量在10 d 时未与对照形成显著差异,但在20 d 和30 d,自由水含量显著高于对照,表明福寿螺主要通过增加自由水含量的方法阻止了体质量的下降。事实上,无脊椎动物普遍存在用水替代组织中损失的有机物的行为。但福寿螺用水替代有机物损失的原因仍不明确,因此需进一步研究福寿螺在饥饿状态下增加自由水含量的原因和潜在作用。动物壳的增长主要来源于钙化过程,软体动物利用外套膜上皮细胞分泌出碳酸钙和壳基质用以构建动物壳。本试验中,福寿螺壳高、壳宽、壳口长和壳口宽在30 d 时均显著低于对照,表明饥饿胁迫下钙化过程受到了抑制。在马氏珠母贝(Dunker)中就观察到饥饿抑制壳生长的现象,但具体抑制机理目前尚不明确,有待进一步研究。
3.2 饥饿胁迫对福寿螺消化腺抗氧化酶系统的影响
SOD 能加速自由基转化为过氧化氢,POD 和CAT 能将过氧化氢转化为水,从而清除了自由基,还原型GSH 既可抵抗活性氧的氧化损伤,同时还是其他抗氧化酶作用的底物。在本研究中,30 d 的饥饿胁迫并未对福寿螺的POD 活性和GSH 含量造成显著影响(图2a,d)。随着饥饿时间的延长,SOD、CAT 活性逐渐上升,第10 d 时SOD、CAT 活性与对照无明显差异(图2b,c),SOD 活性在20 d 和30 d 时显著高于对照。这可能是自由基在SOD 的作用下被还原为HO,随着SOD 活性逐渐升高,HO含量增加,诱导了CAT 的表达增加,致使30 d 时CAT 活性显著高于对照。前期的饥饿胁迫不足以诱导福寿螺消化腺活性氧的大量产生,但随着饥饿时间的延长,机体中产生了大量的自由基,福寿螺受到氧化胁迫,进而调动了福寿螺抗氧化系统的运作。
MDA 是不饱和脂肪酸过氧化的最终产物之一,被认为是自由基介导损伤的良好标志物。MDA产生的数量代表了脂质过氧化的程度,间接反映组织细胞受自由基攻击的程度。MDA 含量在10 d和20 d 时与对照无显著差异,随着时间延长至30 d,MDA 显著高于对照(图2e),这一结果再次证明了前期的饥饿胁迫不能对福寿螺消化腺组织产生氧化损伤,但随着时间延长,脂质过氧化程度加剧,消化腺细胞受到自由基的破坏。MDA 与SOD、CAT 的变化共同说明了福寿螺在饥饿胁迫下的氧化损伤程度和抗氧化系统的运行状态。在管角螺(Gmelin)中也观察到相似的结果。
3.3 饥饿胁迫下福寿螺生化物质含量的变化
饥饿胁迫下,动物会消耗内源性物质以供给机体能量,这些物质的消耗顺序和重要性因物种而异,如无脊椎生物紫贻贝(Linnaeus)和虎纹对虾(Haswell)在食物缺乏的情况下优先消耗蛋白质作为能量来源。方斑东风螺(Link)在饥饿下优先消耗脂肪和糖原,饥饿90 d 后开始消耗蛋白质。高背红螺(Say)在饥饿时首先消耗葡萄糖或糖原,然后利用脂质,最后利用蛋白质。在本研究中,福寿螺在没有食物供应的情况下,脂肪含量于10 d 开始显著低于对照并持续到30 d (图3d),说明10 d 的禁食对福寿螺能量消耗造成影响,随着饥饿时间的延长,糖原含量于20 d 开始显著低于对照(图3b),当饥饿时间延长至30 d 时,福寿螺总蛋白含量显著低于对照(图3a),表明饥饿胁迫下福寿螺可能最优先利用体内存储的脂质为机体提供能量,随后调用糖原和总蛋白。福寿螺的营养消耗策略与腹足纲的方斑东风螺(Link)营养消耗策略类似。
甘油是甘油三酯(脂质的组成成分)水解代谢的最终产物,因此测量甘油含量更能清楚地表明甘油三酯分解的速率。同时,甘油是重要的糖异生前体,生物在长期饥饿状态下会将甘油转变为葡萄糖或糖原用于营养消耗。福寿螺在饥饿10 d 时,甘油含量显著高于对照,表明甘油三酯作为脂质在饥饿前期被大量消耗,甘油三酯的分解速率提高。随着时间延长,福寿螺甘油含量在20 d 和30 d 时显著低于对照,表明福寿螺体内甘油三酯的分解速率下降,可利用的甘油三脂含量降低,福寿螺难以通过甘油三酯的代谢为机体提供充足的能量。
4 结论
本试验探讨了福寿螺面对饥饿胁迫时的生存策略,结果表明,随着饥饿时间的延长,福寿螺自由水含量持续上升,体质量、壳高、壳宽、壳口长和壳口宽的生长受到抑制;总超氧化物歧化酶和过氧化氢酶活性及丙二醛含量随饥饿时间的延长逐渐高于对照;甘油、脂肪、糖原和总蛋白的含量在饥饿30 d后均显著低于对照。福寿螺通过减缓生长、提高抗氧化能力、消耗内源性生化物质等生理机制来缓解胁迫带来的影响。结果将有助于加深我们对福寿螺的环境响应的理解,并为研究其防控策略提供参考。另外,本试验发现福寿螺在饥饿胁迫下会提高螺体内自由水含量,但该行为的具体原因和潜在作用尚未明确,有待进一步研究。
