嘉陵江中游蓬安段鱼类群落功能多样性研究
2022-06-09向伶俐鲍江辉张朝硕米湘媛
张 倩 曾 燏 肖 瑾 向伶俐 鲍江辉, 张朝硕, 米湘媛, 段 明
(1. 西华师范大学生命科学学院, 国家淡水渔业工程技术研究中心(武汉)西南分中心, 南充 637002;2. 中国科学院水生生物研究所, 武汉 430072; 3. 中国科学院大学, 北京 100049)
群落功能多样性 (Community functional diversity) 是生物多样性的重要组成部分[1,2], 是联系物种与生态系统功能的桥梁, 能有效预测生态系统功能变化[3], 近几十年来成为生物多样性研究的热点之一。环境变化和人类活动对淡水生物多样性的威胁日益加剧, 而鱼类群落是水生生态系统中的顶级群落, 其群落结构和生物多样性指数是影响群落功能多样性的重要内在因素[4—6], 如Baptista等[7]研究了蒙德戈河口1988—2012年内3个关键时期的鱼类群落功能多样性, 结果表明人为干扰和工程建设(大坝建设、边缘路堤修建和河道整治等)导致的气候变化和极端天气使该河口发生了严重的水文变化, 鱼类群落结构也随之发生显著改变, 导致功能丰富度由38.00下降到22.00。另外, 鱼类群落功能多样性变化不仅能有效反应水生生态系统功能, 同时也是评价水生生态系统健康与否的重要依据[8—10]。
鱼类群落功能多样性主要关注与群落功能密切相关的物种功能性状, 包括形态性状如体长、体型和口的位置, 生态性状如食性、垂直分布、生长系数、脆弱性、卵黏性和生活史类型等, 其功能多样性的计算则是通过对所选取的功能性状的赋值和量化来实现[11—13]。早期, 生态学家对功能多样性的评估方法中因缺乏对物种丰度的考虑而难以准确反映生态系统功能变化。2008年Villéger等[14]在对前人的方法进行研究和优化的基础上, 提出了3个独立的功能多样性指数, 即功能丰富度、功能均匀度和功能离散度, 这三类功能多样性在植物学和动物学领域得到广泛应用, 如薛倩妮等[15]对山西五鹿山森林群落木本植物功能多样性的研究; Shaffer等[16]研究了大型水坝拆除对近岸鱼类群落功能多样性的影响; Docherty等[17]研究了燃烧稀树草原对鸟类物种丰富度和功能多样性的影响等。但目前关于生物群落功能多样性的研究主要集中于植物和微生物群落[18—20], 而关于鱼类群落功能多样性的研究相对较少[21—23]。
嘉陵江是长江水系中面积最大的支流, 由陕西发源流经多地, 最后由重庆汇入长江。蓬安段位于嘉陵江中游, 是连接嘉陵江上游和下游的重要枢纽,其地形结构复杂而独特, 为多种鱼类提供繁殖、育幼和栖息的场所[24—26]。蓬安段位于嘉陵江中游, 鱼类种类丰富, 约有57种[24]。但近年来由于人类活动的影响(梯级水利工程和城市化发展等), 水文生态环境(温度、流速、水位和溶解氧)发生较大改变[27—29]。据史料记载, 在20世纪60年代以前蓬安段洪水期与枯水期水位差距较小, 而当前洪水期与枯水期水位差距明显, 平均水位差可达15 m[30]。本文依据嘉陵江中游蓬安段洪水期和枯水期的鱼类资源调查数据, 结合11项鱼类功能性状类型, 运用Villéger等[14]的多维功能多样性计算方法, 探究了嘉陵江中游蓬安段鱼类群落功能多样性对水位洪、枯变化的响应及其与水环境因子之间的关系。
1 材料与方法
1.1 样本采集
本研究调查于嘉陵江中游蓬安段, 选取了分布于金溪电站和马回电站之间的S1—S6共6个采样点(图 1), 洪水期采样时间为 (6、7和8月) 和枯水期为(11、12和1月), 不同的采样点均采用随机采样方法进行捕捞作业, 2014—2019年洪、枯期共搜集鱼类资源总量57959尾。由于嘉陵江鱼类资源丰富, 所捕获的渔网规格也有差异, 本研究的主要捕捞方式包括流刺网(内网目3 cm, 外网目30 cm)、撒网(网目3 cm)和地笼(网目1.5 cm)等。每月租用当地渔民渔船进行为期3d捕捞跟踪调查,每天上午和下午各2次撒网, 每月共计12网次, 每次收网时现场进行物种鉴定和数量统计及常规的生物学特征测量, 鱼类标本用10%福尔马林溶液保存并带回西华师范大学生命科学学院实验室。每次放网前均测量水中pH、水温、透明度、流速及溶氧量等环境数据, 其中水位数据从蓬安水文监测站获取, 年际变化趋势如图 2所示, 总磷和总氮数据从蓬安生态环境监测站获取。本研究将水环境变量分为化学环境因子(pH、溶氧、总氮和总磷) 和物理环境因子(流速、水位、透明度和温度) 两类进行研究(表 1)。

表1 嘉陵江中游蓬安段2014—2019年洪、枯期水环境因子(平均值±标准差)Tab. 1 Water environmental variable in Peng’an section of the middle reaches of Jialing River during flood and dry seasons from 2014 to 2019 (mean±SD)

图1 嘉陵江中游蓬安段鱼类采样点分布图Fig. 1 Distribution of fish sampling sites in Peng’an section of the middle reaches of Jialing River

图2 嘉陵江中游蓬安段2014—2019年水文特征变化趋势Fig. 2 Variation trend of hydrological characteristics in Peng’an section in the middle reaches of Jialing River from 2014 to 2019
1.2 功能性状选取
为探究嘉陵江中游蓬安段鱼类群落功能多样性, 参考Shaffer等[16]、Pool等[31]和张晓妆等[32]的研究方法, 选取了与鱼类摄食能力(食性、营养级和口的位置)、游泳能力(最大体长和体型)、生态适应能力(栖息水层、适应水温和流速喜好)、群落动态(脆弱性指数和生长系数K)及生活史策略(周期主义、均衡主义和机会主义)相关的11项鱼类功能性状类型。上述功能性状均从鱼类标本和《四川鱼类志》[33]、鱼类数据库(Fishbase)及参考文献中获得[34](表 2), 2014—2019年蓬安江段鱼类名录及其功能性状见表 3。

表2 嘉陵江中游蓬安段鱼类功能类别、性状与类型Tab. 2 Functional category, functional trait, and type of functional traits of fish species in Peng’an section of the middle reaches of Jialing River

表3 2014—2019年嘉陵江中游蓬安段鱼类名录及其功能性状Tab. 3 List of fishes and their functional traits in Peng’an section of the middle reaches of Jialing River from 2014 to 2019
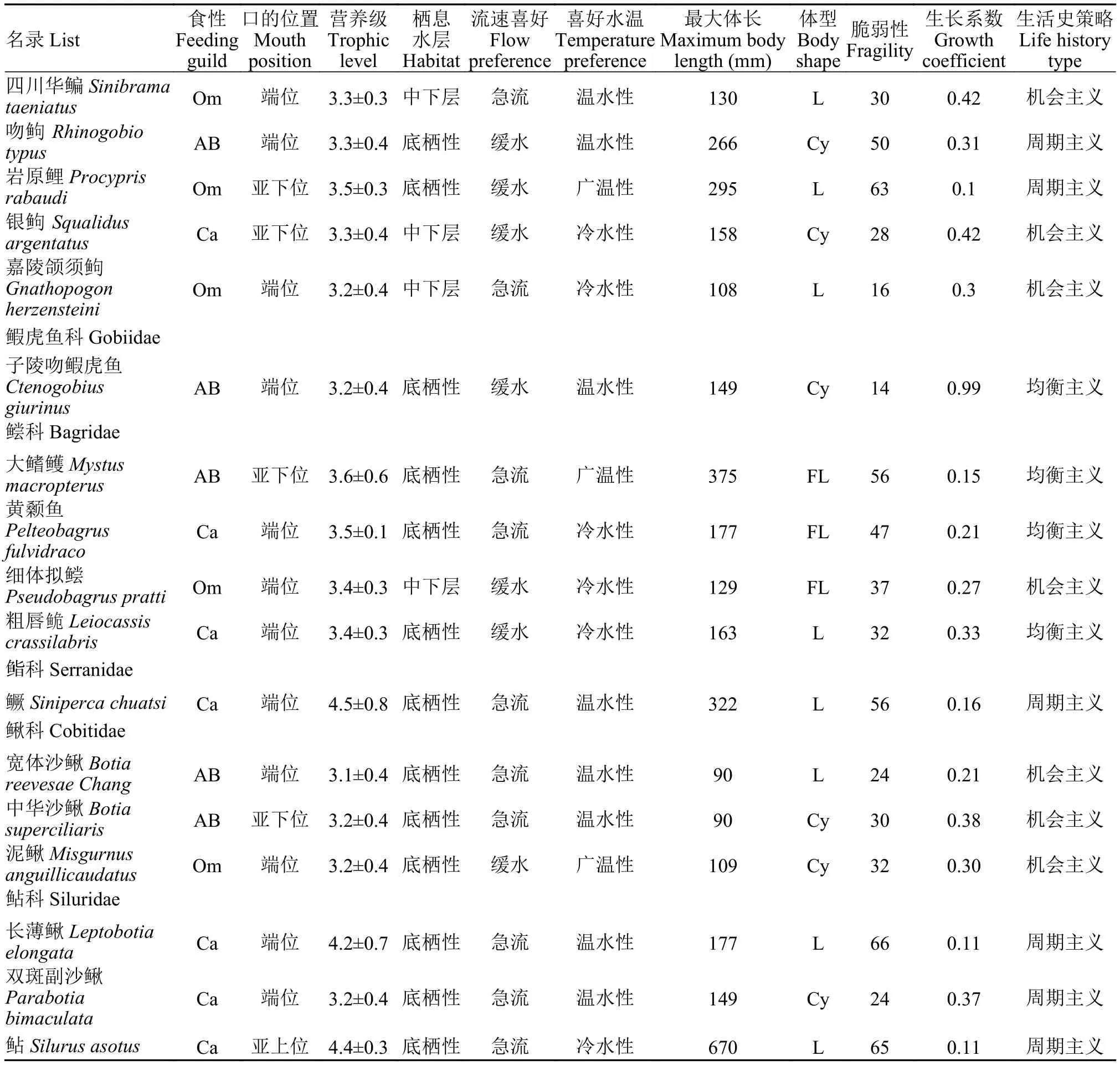
续表3
1.3 数据分析与处理
生物多样性指数本研究选取物种丰富度指数SA、香农威纳指数H、Pielou均匀度指数和Simpson多样性指数来反映嘉陵江中游蓬安段鱼类群落在物种水平上的多样性。其分别的计算方法如下:

蓬安江段鱼类群落优势种由于优势种对鱼类群落结构具有明显的控制作用, 故本研究采用Piankas[35]的相对重要性指数(Index of Relative Importance,IRI)来衡量蓬安江段鱼类群落洪、枯期及年际生态优势度, 其计算公式为:

式中,N%为某一种鱼类捕获数量占总捕获数量百分比;W%为某鱼类物种重量占所捕获鱼类总重量百分比;F%为某鱼类物种在调查时的出现频率。IRI>1000为优势种, 100<IRI<1000为重要种。
鱼类群落功能多样性指数本研究选取将物种相对丰度和多种功能性状均考虑在内的功能丰富度Functional richness (FRic)、功能均匀度Functional evenness (FEve)及3个功能离散度[功能离散度Functional divergence (FDiv)、功能分散度Functional dispersion (FDis)[14]和二次熵指数Quadratic entropy (Rao)[36]。
(1) 功能丰富度FRic: 由功能性状空间内形成的最小凸多边形体积来计算[14]。
(2) 功能均匀度FEve公式如下:
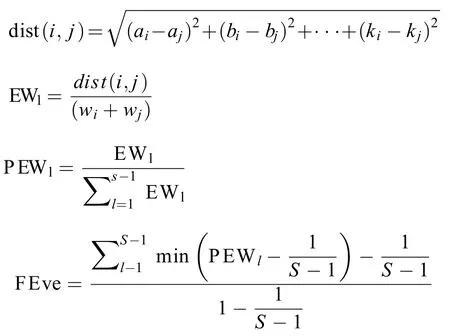
式中,a—k表示多维性状空间中物种i和j的11个功能性状; dist(i,j)表示物种i和j之间的欧式距离;EWl为分支长度;wi和wj分别为物种i和j的相对丰度;PEWl为分支长权重;S为物种数。
(3) 功能离散度FDiv计算公式如下:

式中,S为物种数,T为功能性状数目,gk为性状k的重心,xik为物种i性状k的值,wi为物种i的相对丰度,为物种i的功能性状值距离重心的欧氏距离,Δd为以相对丰度为权重的离散度。
(4) 功能分散度FDis计算公式如下:

式中,aj为物种j的相对多度,xij为物种j第i个性状值。zj是物种j到重心c的加权距离。
(5) RAO的二次熵指数计算如下:

式中,pi为物种相对多度, 则S为物种数,dij为物种i和j的相异程度, 变化于 (0, 1) 之间, 该值为0时表示两物种具有完全相同的功能性状, 该值为1时表示两物种具备完全不同的功能性状。
数据处理相对重要性指数用Excel 2019计算; 所有功能性状值经过标准化后再进行功能多样性计算, 功能多样性指数用R 3.5.1软件dbFD程序包计算; 用Past软件进行SPIMPER分析; 用Prism软件进行图形绘制; 并用SPSS 25.0软件分析以下数据:(1)独立样本t检验分析各功能多样性指数洪、枯期的差异; (2)用单因素方差分析和多重比较(LSD)分析各功能多样性指数间年际差异; (3)用皮尔逊相关性分析, 探明各功能多样性指数与环境变量之间的关系; (4)功能多样性结果图形绘制。
2 结果
2.1 2014—2019年蓬安段鱼类群落生物多样性指数
总体来看, 2014—2019年洪水期的4类生物多样性指数均高于枯水期(图 3)。
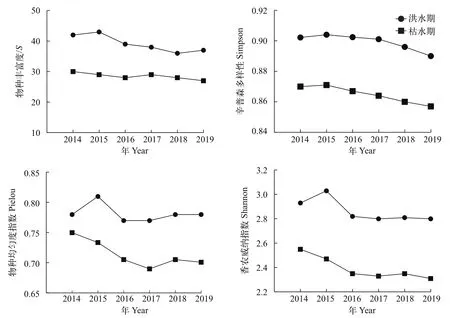
图3 2014—2019年蓬安段鱼类群落生物多样性变化Fig. 3 Fish community biodiversities in Peng’an section from 2014 to 2019
2.2 蓬安段洪、枯期鱼类群落物种组成及优势种
2014—2019年洪水期和枯水期在蓬安段共调查到鱼类57959尾, 分属分属于3目, 6科, 38属45种。其中鲤科鱼类洪水期和枯水期物种数差异明显, 洪水期41种, 枯水期为34种。其中峨眉鱊Acheilognathus omeiensis、华鳈Sarcocheilichchthys sinensis、宽体沙鳅Botia reevesae Chang、似Pseudogobio vaillanti、四川华鳊Sinibrama taeniatus、银鲴Xenocypris argentea和中华沙鳅Botia superciliaris仅在洪水期出现, 贝氏䱗Hemiculter bleekeri仅在枯水期出现。由表 4可知, 䱗Hemiculter leucisculus、蛇Saurogobio dabryi、鲫Carassius auratus、鲤Cyprinus carpio和黄尾鲴Xenocypris davidi为各年份洪、枯期均出现的优势种鱼类。
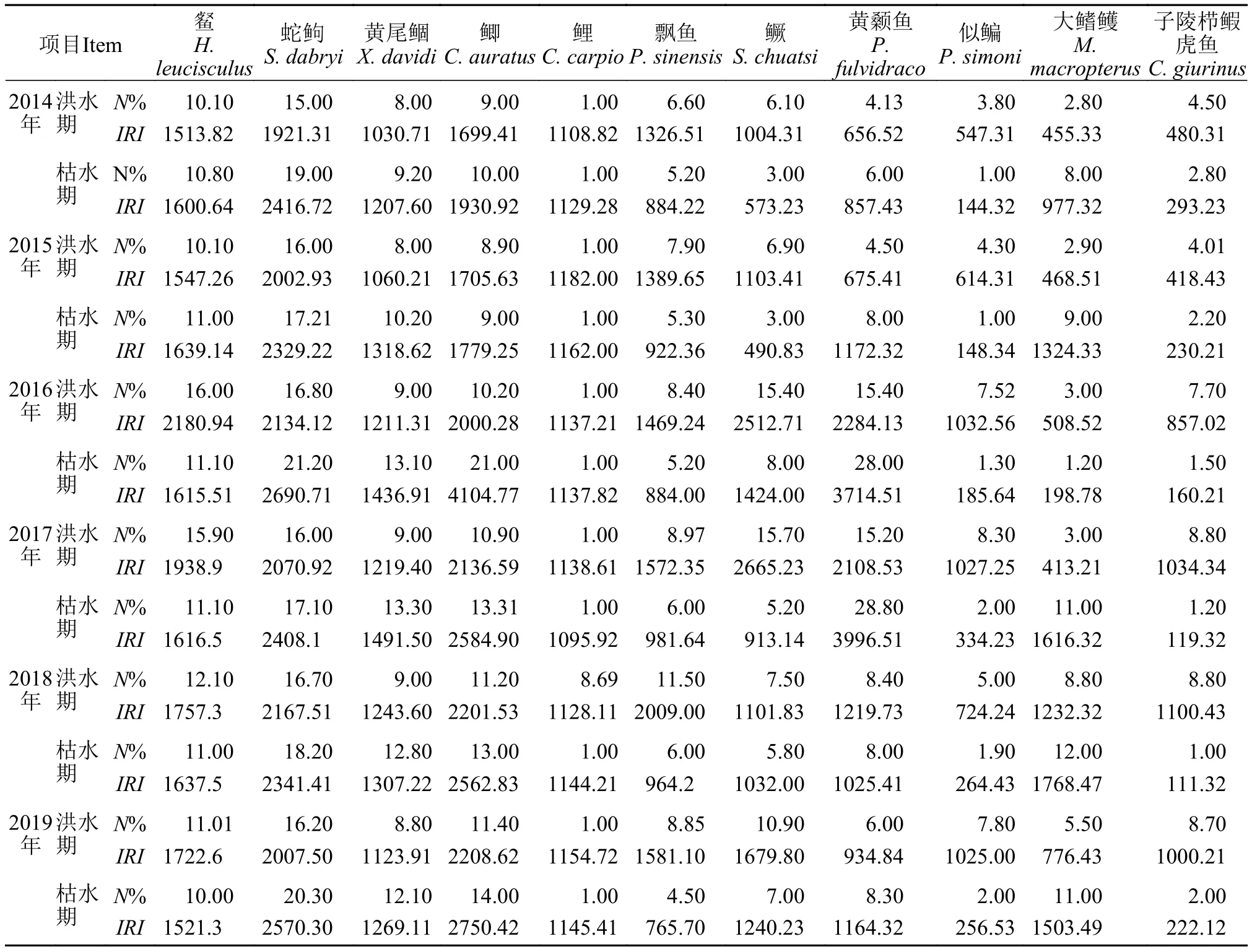
表4 蓬安段2014—2019年洪、枯期相对重要性指数IRI大于1000的优势种鱼类及其相对丰度Tab. 4 Dominant species with IRI greater than 1000 and their relative abundance in flood and dry seasons of Peng’an section from 2014 to 2019
本研究将2014—2019年鱼类平均相对丰度做SIMPER分析, 将累积贡献率超过90%的鱼类视为群落结构主要差异贡献种, 分析结果显示(表 5), 大鳍鳠、子陵吻鰕虎鱼、似鳊、黄颡鱼、黄尾鲴、鳜、飘鱼、蛇、鲫和䱗是造成洪水期和枯水期鱼类群落结构差异的主要贡献种。

表5 嘉陵江蓬安段主要鱼类物种对2014—2019年洪水期和枯水期鱼类群落结构差异的贡献率Tab. 5 Contribution rates of major fish species to the difference of fish community structure during flood and dry seasons from 2014 to 2019 in Peng’an section of the middle reach of Jialing River
2.3 2014—2019年蓬安段鱼类群落功能多样性
独立样本t检验表明(图 4), 各年洪水期的FRic、FDiv、FDis和Rao指数均为洪水期显著高于枯水期(P<0.05); FEve指数2014、2018和2019年无显著性差异, 2015、2016和2017年均为洪水期显著高于枯水期。

图4 嘉陵江中游蓬安段2014—2019年洪、枯期鱼类群落功能多样性指数Fig. 4 Functional diversity indices of fish community in Peng’an section of the middle reaches of Jialing River during flood and dry seasons from 2014 to 2019
方差分析(ANOVA)结果显示, 2014—2019年的FRic洪水期呈现逐年降低趋势, 枯水期无显著年际变化; FEve洪水期表现为先上升后下降的趋势,枯水期为先下降后上升的趋势; FDiv洪、枯期均为先上升后下降的趋势; FDis和Rao为先上升后下降的趋势。
2.4 蓬安段鱼类群落功能多样性与环境因子的相关性分析
将功能多样性指数与所调查的洪、枯期环境因子进行Pearson相关性检验结果显示 (表 6), 蓬安段水域物理和化学环境因子均是影响功能多样性的重要因素。洪、枯期均表现为, FRic、FDiv、FDis和Rao与温度、水位和流速均显著正相关; 各功能多样性指数与透明度均显著负相关(P<0.01)。FEve与pH显著正相关; FRic和FEve与总氮、总磷和溶氧显著正相关(P<0.01)。此外, 基于相关系数强度差异标准[37], 洪、枯期功能多样性指数与环境因子相关性程度呈现较大的差异, 如洪水期的FEve与流速中度相关, 枯水期的FEve与流速则呈现弱相关关系; 枯水期的FDis和Rao与pH呈中度相关, FDiv和Rao与总磷中度相关, 洪水期则均呈现弱相关关系。
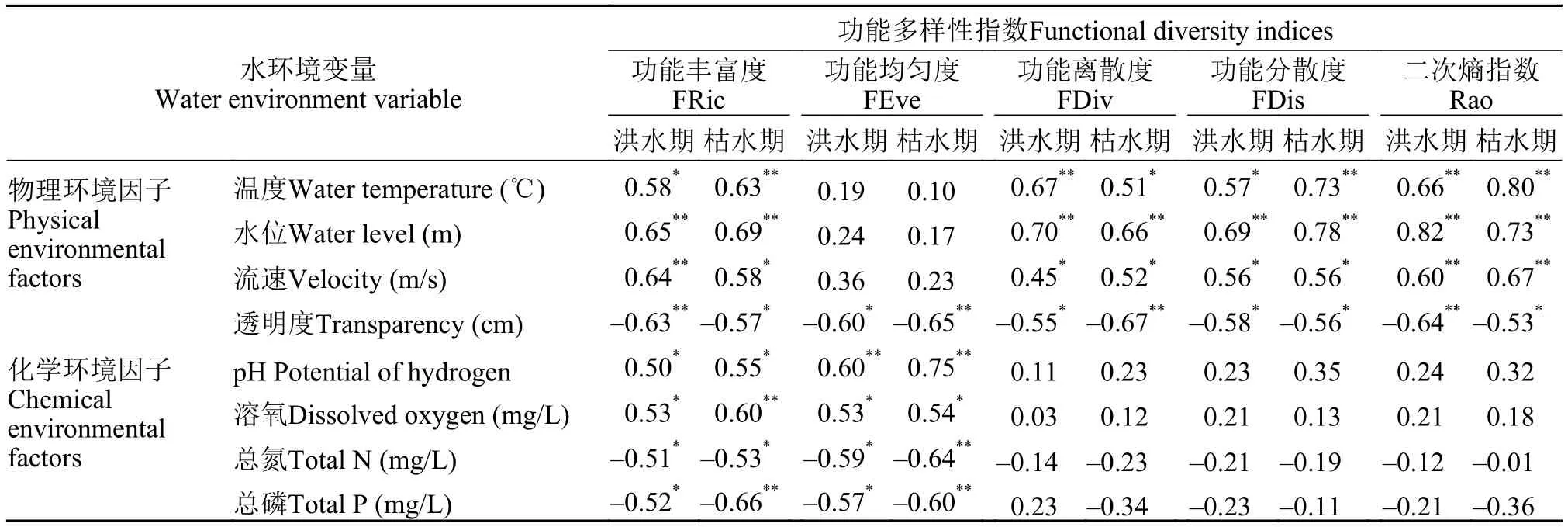
表6 嘉陵江蓬安段群落功能多样性指数与环境因子的Pearson相关性分析Tab. 6 Pearson correlation coefficients between functional diversity indices and environmental factors in Peng’an section of the middle reaches of Jialing River
3 讨论
3.1 嘉陵江中游蓬安段功能多样性指数变化
本研究结果显示, 鱼类群落功能多样性指数和生物多样性指数具有相似的变化规律, 即洪水期显著高于枯水期, 表明蓬安段枯水期水生生态系统功能较脆弱[38]。可能原因是随着蓬安段水域的洪、枯期更替使得鱼类栖息环境改变, 难以适应当前水环境的鱼类会暂时性地进入相连通的周边小生境(支流、小水体、独立湿地和江心岛等)中避难场所, 而能适应当前水环境的鱼类就成为优势类群,导致生物多样发生改变, 进而改变了鱼类群落结构和功能[39]。相似的研究结果在其他流域也有表现,如苏国欢等[40]发现大坝截流后金沙江观音岩水电站鱼类群落的物种丰富度和功能多样性指数都呈现显著下降的情况, 主要原因是大坝截流后, 物种丰富度指数呈下降趋势, 表现为拥有特定功能特征的鱼类减少或消失, 降低了该流域的功能多样性。
功能丰富度FRic是群落生态位空间填充的最佳预测指标, 该指数与物种丰富度具有显著正相关关系[15], 不同物种对生态系统功能的贡献不同[21]。如张晓妆等[32]研究发现春季海洲湾功能丰富度显著低于秋季, 原因是竹筴鱼(Trachurus japonicus)和鳓(Ilisha elongate)等7种鱼类仅在秋季出现, 缩小了春季鱼类群落的生态位空间。本研究结果与此相似, 蓬安段水域枯水期FRic较洪水期显著降低, 结合生物多样性指标季节变动分析结果来看(图 3),枯水期随着水温、水位和流速等生境因子的下降,可能会导致一些不适应该环境的鱼类退缩到周边的一些支流和小水体内, 如峨眉鱊(A. omeiensis)、华鳈(S. sinensis)、宽体沙鳅(B. reevesae)、似(P.vaillanti)、四川华鳊(S. taeniatus)、银鲴(X. argentea)和中华沙鳅(S. superciliaris)等7种鱼类的消失,出现枯水期物种丰富度指数下降, 从而降低了功能性状空间的范围和数值, 致使枯水期鱼类群落部分生态位空间缺失[41]。
功能均匀度FEve通常衡量群落在所占据性状空间分布是否均匀和资源利用是否充分, 进而作为群落生产力、抵抗力稳定性评价指标[15,19]。如向琳等[19]发现高海拔地区鹿角杜鹃 (Rhododendron latoucheae) 群落灌木层植物FEve较低, 原因是植物群落在高海拔环境下随着温度的下降资源的利用率降低, 功能性状空间未被完全利用, 导致群落抵抗力稳定性较脆弱, 遭到破坏后难以恢复。本研究蓬安段2015、2016和2017年FEve枯水期较低, 表明该时期鱼类群落生产力和抵抗力稳定性降低, 其中一个原因可能是枯水期的Pielou均匀度指数较低,导致群落中物种在数量上分配不均匀[42], 造成枯水期鱼类群落对资源利用不充分。另一个原因可能是枯水期鱼类群落中高生长系数、低脆弱性指数的优势种鱼类相对丰度下降(表 3和表 4), 如飘鱼(P. sinensis)等, 再加上枯水期蛇(S. dabryi)和黄尾鲴 (X. davidi)等机会主义鱼类相对丰度反而显著增高, 导致枯水期鱼类生命周期普遍较短, 个体较小, 鱼类群落功能脆弱性增加[41,43]和游泳能力较弱进而导致鱼类寻找食物资源能力减弱[44], 导致2015、2016和2017年枯水期鱼类群落对饵料资源的利用率下降[16,31]。
功能离散度(FDiv、FDis和Rao)是衡量群落内物种功能性状相似性及群落竞争强弱的指标, 功能性状相似的物种越多群落竞争越激烈, 生态系统功能越不稳定[45]。如张晨[46]对宜昌江段建坝前后鱼类群落功能多样性变化研究发现, 在建坝后, 鱼类群落功能离散度上升, 原因是杂食性且位于中上层的鱼类飘鱼(P. sinensis)和䱗(H. leucisculus)等食性较广的鱼类相对丰度增加, 使群落中与摄食能力有关的功能性状相似性减少, 鱼类对食物和栖息地的竞争减弱, 生态系统功能较稳定。本研究枯水期功能离散度显著低于洪水期, 原因可能是枯水期水流速度相对较慢且水位相对较低, 从而导致鱼类的饵料资源交换率降低和栖息地空间缩小[47], 一些资源利用方式相似的物种如以营底栖生活和以底栖无脊椎动物为食的大鳍鳠(M. macropterus)、鲫(C.auratus)和蛇(S. vdabryi) 等鱼类相对丰度升高(表 3和表 4), 摄食生态位和栖息地空间重叠效应增强, 群落对饵料资源和栖息地空间的争夺剧烈导致生态系统稳定性下降[3,11]。
鱼类群落功能多样性的年际波动是鱼类群落功能的不稳定性和脆弱性的表现。如Mao等[48]对呼伦湖鱼类群落功能多样性对水环境变化的响应研究发现, 功能多样性从2005年至2016年均呈现出先下降后上升的波动趋势, 其主要原因是人为的长时间干扰导致河流中鱼类栖息地丧失和破坏从而造成了河流生态系统功能的不稳定性和脆弱性。嘉陵江中游蓬安段鱼类群落结构在年际上也存在显著波动和不稳定, 这种趋势反映出人为活动和环境变化对蓬安段水生生态系统的影响是一个长期和不稳定的过程。
3.2 嘉陵江中游蓬安段鱼类群落功能多样性与水环境因子的关系
水环境对于鱼类聚集及群落分布具有重要影响。当鱼类生存环境发生改变时, 适应该环境的鱼类会随之出现, 不适应该环境的鱼类会随之消失[49]。水位、温度、流速和透明度等物理环境因子主要是通过影响鱼类饵料资源分布和栖息地环境从而间接影响鱼类群落分布和资源利用格局, 这些非生物因子对鱼类群落的影响在其他流域中都较为明显。如Baptista等[7]认为造成葡萄牙南、北海岸鱼类群落结构和组成差异的原因是鱼类对水温的适应性选择结果; Eros等[50]发现芬兰125个湖泊的水位和水体透明度与其鱼类群落功能多样性呈现显著正相关关系。本研究与以上研究结果相似, 洪、枯期各功能多样性指数与物理环境因子均具有显著相关关系, 其中透明度与各功能多样性指数显著负相关。而透明度是衡量水体浮游生物和藻类含量的指标, 透明度较高的水体水较瘦[52], 不利于如似鳊(P. simoni) 等以藻类和植物碎屑为食的鱼类生长。该结论证明枯水期鱼类群落能利用的饵料资源减少会导致鱼类对其他食物资源的竞争加剧, 导致生态系统功能不稳定[19,45]。水位、温度和流速对功能多样性指数存在显著影响, 可能原因是枯水期处在寒冷的冬季, 温水性鱼类如似鳊(P.simoni)和鳜(S. chuatsi)等相对丰度减少。再加上枯水期水位较低, 干流和支流及独立湿地、江心岛间的连通性降低, 且水流速度较慢缩小了上层鱼类如飘鱼(P. sinensis)和䱗(H. leucisculus)的栖息地空间和饵料资源导致功能多样性下降[47,52]。另外, 导致洪、枯期功能多样性指数与环境因子相关性程度呈现较大差异的另外一个可能原因是, 洪水期水流速度相对较快, 对小型鱼类影响较明显, 且有相关研究证实, 适宜的流速刺激能促进鱼体快速生长[53];而枯水期则相反, 水流较缓, 有利于小型鱼类的生存, 从而导致鱼类群落较脆弱, FEve值降低。
鱼类的生存往往也受到水体化学环境因子的影响[54], 水体化学条件的变化会影响鱼类生长、食物资源分布及栖息地进而造成鱼类群落的生产力和抵抗力稳定性。影响功能多样性的化学环境因子因群落类型和水域的不同而有差异。如Mao等[48]发现太湖水体鱼类群落FDis与水中pH显著负相关;Gusmao等[55]发现除了pH外, 总氮、总磷对潮间带大型底栖无脊椎动物功能多样性也具有显著影响。水体酸碱度是影响鱼生长的重要因素, 蓬安段洪水期水体呈现弱碱性, 而弱碱性水体更适合鱼类生长[44], 枯水期水体偏中性, 鱼类群落体型偏向小型化使该时期鱼类运动能力较弱, 影响鱼类寻找食物的能力进而导致功能多样性值显著降低。蓬安段流域总氮和总磷的值虽然并未超过《地表水环境质量标准》限值, 但枯水期总氮和总磷均显著高于洪水期, 而总磷和总氮的升高会导致水体趋于富营养化造成水中溶氧量降低[56], 进而影响鱼类生存和功能性状分布, 导致枯水期FRic和FEve值显著下降, 这一结论与Paker[8]的研究结果一致。
