甘肃黄河沿岸39份枣品种亲缘关系的SSR分析
2022-06-09胡秉芬李广宇孟亚雄陈翠莲张广忠张正振
胡秉芬,李广宇*,孟亚雄,刘 荟,陈翠莲,马 玮,张广忠,张正振
(1.甘肃省林业科学研究院,甘肃 兰州 730020;2.甘肃农业大学 农学院,甘肃 兰州 730070 ;3.平川区林业局,甘肃 白银 730900)
枣(Ziziphusjujuba)是原产我国的特有果树,栽培历史悠久,约有 900 多个品种[1-3]。甘肃是枣的原产地之一,枣资源丰富。在长期的栽培与驯化过程中,产生了许多变异类型,近年频繁引种,导致甘肃黄河沿岸枣区品种存在来源混淆、同物异名和同名异物的现象,品种间的遗传关系混乱,阻碍了实践生产和品种培育。传统的形态学鉴定费时费力,易受外界环境因素的影响,且难以区分性状相似的品种。SSR(simple sequence repeat)分子标记在国际植物新品种权保护联盟(UPOV)的BMT分子测试指南中被确定为构建 DNA 指纹数据库的标准标记方法[4],具有多态性高、易检测、重复性高、数量丰富、共显性等优点,非常适用于品种遗传多样性和亲缘关系的鉴定,已广泛应用于苹果、梨、桃、柑桔等各种经济树种的种质资源鉴定[5-6]、亲缘关系分析[7-8]、遗传多样性分析[9-11]、遗传图谱构建[12]、基因定位[12-13]和分子标记辅助育种[14]等方面。在枣树研究方面,北京林业大学利用SSR文库和PCR技术开发了301个多态性枣SSR标记[15],南京农业大学利用454高通量测序技术对枣基因组进行部分测序,找出15 036个微卫星重复序列,并对其特征进行了初步分析[16],河北农业大学利用全基因组数据并与近缘物种进行比较,全面分析了枣基因组SSR特征,共设计出30 565个SSR引物,公布了725对多态性SSR引物[17];麻丽颖等[18]利用SSR标记构建了枣指纹图谱,王斯琪等[19]、孙学超等[20]利用SSR标记进行了枣树亲本鉴定,殷晓等[21]用SSR标记研究了陕北54个枣品种遗传结构,刘秀云等[22]对来自全国22个省区的255个枣品种进行了SSR分析,揭示了它们的亲缘关系和群体遗传结构,喇菲菲等[23]利用SSR分析了84个枣品种的遗传多样性,刘琰玮等[24]用SSR标记技术鉴定了枣树多倍体种质资源。本文在前人研究的基础上,采用SSR分子标记技术分析甘肃黄河沿岸的39份枣品种的遗传多样性和亲缘关系,为枣种质资源的收集、保存和分子辅助育种提供理论依据。
1 材料与方法
1.1 试验材料
39份供试枣树叶样于2018年8月采自甘肃省沿黄灌区的景泰县、靖远县和平川区,其中景泰县五佛乡枣树品种园5份,平川区枣树示范园5份,靖远县甘肃省林业科学研究院枣树示范研究园27份,2份材料分别来自小口村和敦煌市鸣山村的老枣树(表1)。
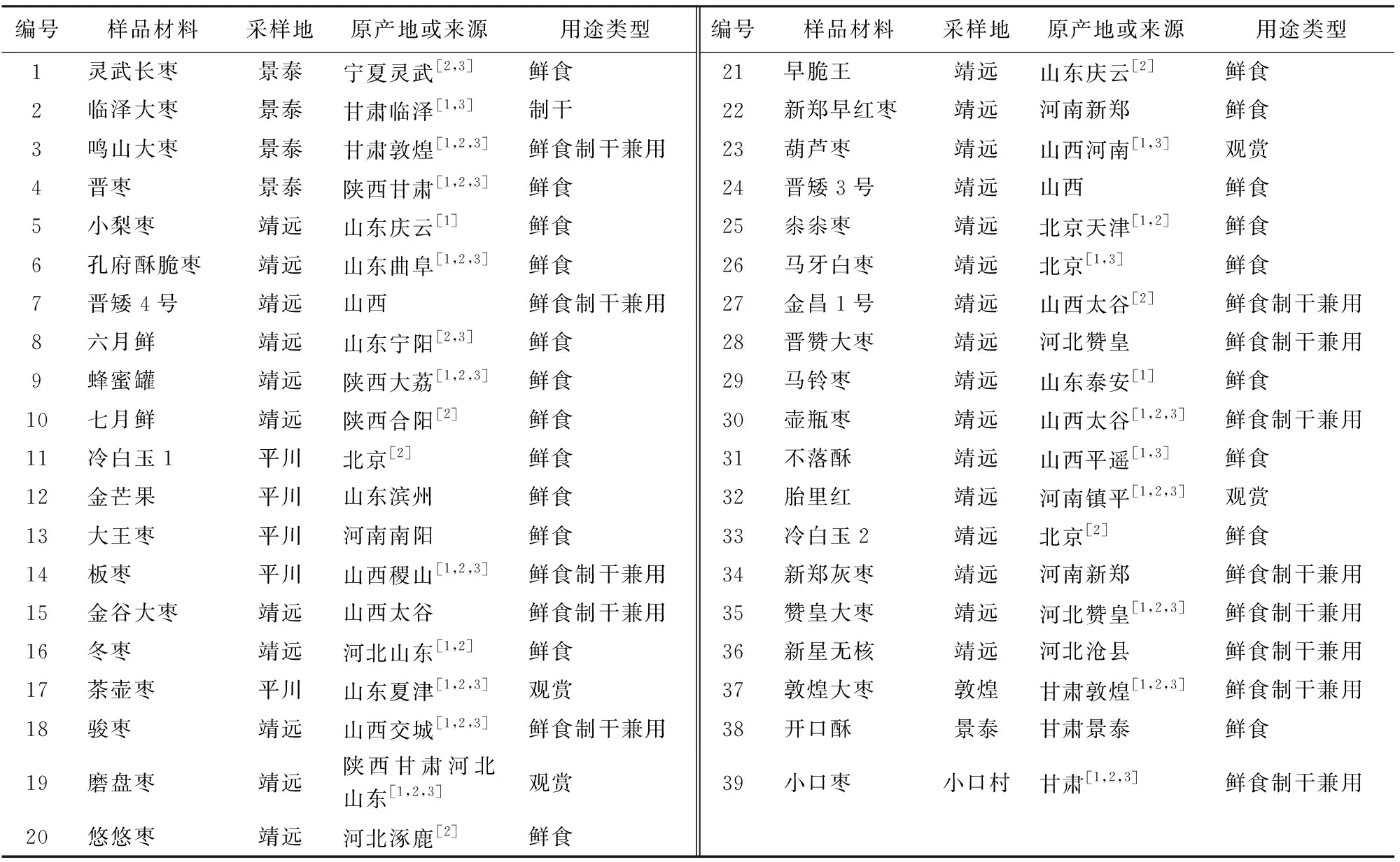
表1 39份枣树材料Table 1 Materials of 39 jujube cultivars
1.2 试验方法
1.2.1 枣树基因组DNA的提取 采集幼嫩、新鲜的枣树叶片,利用TIANGEN植物基因组DNA试剂盒提取样本DNA,IMPLEN纳米光度计检测DNA浓度和纯度,置-20 ℃冰箱保存待用。
1.2.2 SSR引物 20对SSR引物序列信息来源于《枣品种鉴定技术规程SSR分子标记法》[25]和公开发表的枣树文献[17,18,20,26]。引物均由北京六合华大基因科技有限公司合成(表2)。

表2 SSR引物序列与名称
1.2.3 PCR扩增与电泳检测 PCR 扩增体系为(10 μL):模板DNA 2.0 μL(约90ng);2×Taq PCR MasterMix 5.0 μL;正向Primer和反向Primer各0.5 μL(10 μM);ddH2O 2.0 μL。PCR反应程序为:94 ℃预变性3 min;94 ℃变性30 s;53~65 ℃退火30 s;72 ℃延伸1 min,30个循环;72 ℃延伸5 min,4 ℃保存。扩增产物用8%的聚丙烯酰胺凝胶电泳检测。电泳结束后用硝酸银染色观察结果,照相保存图片。DNA marker为pBR322。
1.2.4 数据统计及分析 依据扩增产物在电泳凝胶上的迁移率,分别以“1”和“0”表示同一位置谱带的有无,建立原始数据阵。用NTSYS-pc V2.10软件计算品种间遗传距离(GS),用SAHN程序和UPGMA方法进行聚类分析,并通过Tree plot模块生成聚类图。等位基因比率=(等位基因/n)×100%,n为每个SSR标记所测位点的等位基因总数。
2 结果与分析
2.1 SSR扩增结果
SSR分子标记扩增结果见表3。20对引物共扩增出142个清晰条带,均为多态性条带,多态性百分率为100%。每对引物扩增的条带数为3~13个,平均每对引物扩增的条带数为7.10个,扩增片段大小在70~300bp。引物BFU0308扩增的条带数最多,为13条,扩增条带最少的引物为JSSR198和JSSR289,各有3条。20对SSR引物在39份枣种质中均能获得清晰的条带。图1和图2是引物JSSR250和JSSR93扩增的结果。
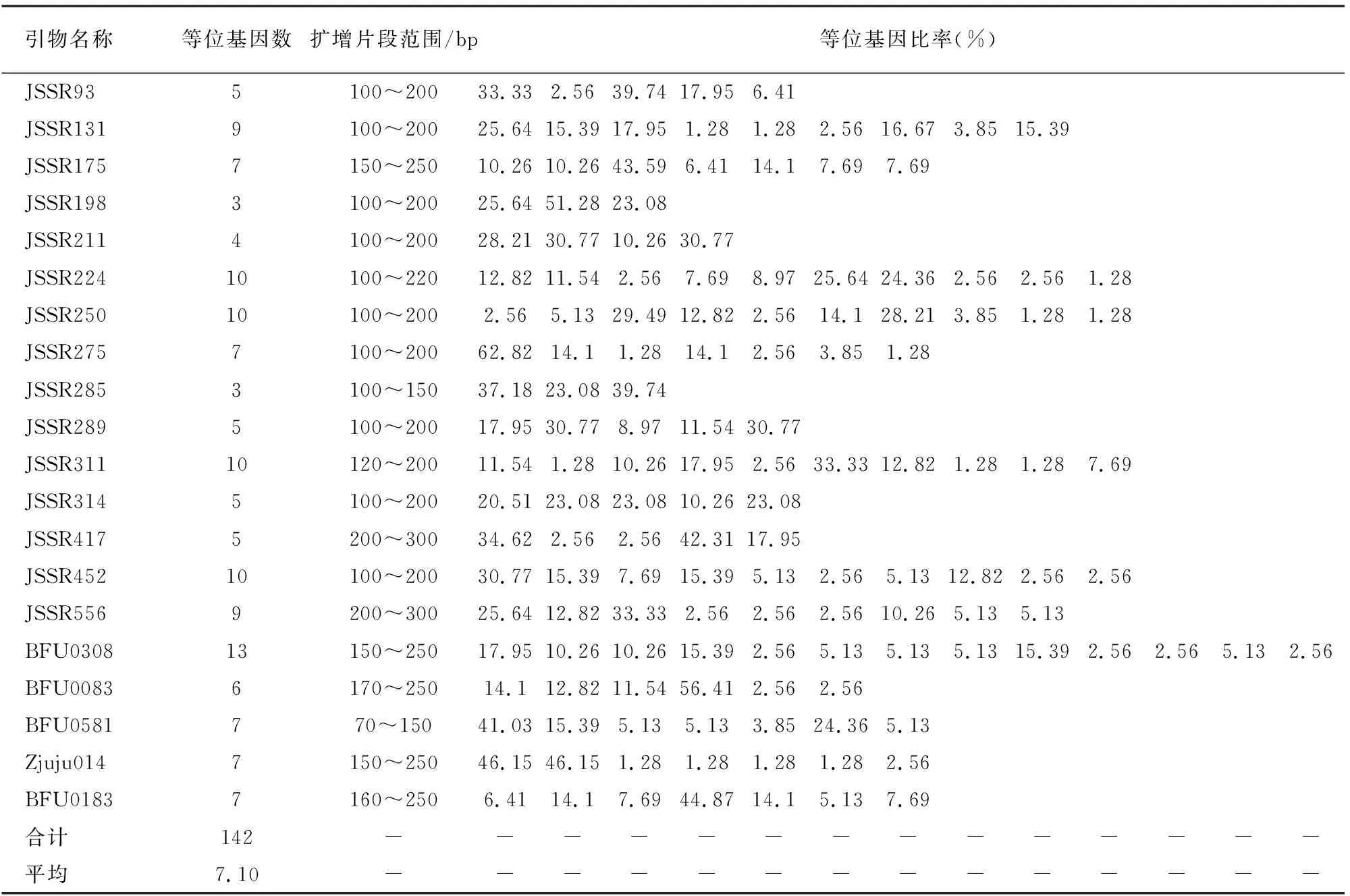
表3 SSR引物扩增情况
2.2 遗传多样性分析
群体中两两个体之间的遗传距离及平均遗传距离是度量群体遗传多样性指标之一[27]。本研究中39份供试枣品种间的遗传距离(GS)在0.000 0~0.993 8,平均遗传距离为0.558 7。其中原产山西稷山的板枣[1,2,3]与其他品种的遗传距离在0.408 9~0.993 8,平均遗传距离最大为0.729 8,表明板枣与其他供试品种亲缘关系最远;原产于北京的冷白玉1[2]次之,与其他品种的遗传距离在0.014 7~0.970 5,平均遗传距离为0.714 6;原产于甘肃的临泽大枣[1,3]与其他品种的遗传距离在0.054 8~0.814 7,平均遗传距离最小为0.457 2,表明临泽大枣与其他供试品种亲缘关系较近。根据两两供试品种之间的平均遗传距离,可以将每份试材与其他供试品种间的亲缘关系由远及近排序为板枣(0.729 8)、冷白玉1(0.714 6)、悠悠枣(0.692 5)、马牙白枣(0.689 0)、新郑灰枣(0.675 5)、七月鲜(0.664 1)、磨盘枣(0.640 4)、尜尜枣(0.597 6)、孔府酥脆枣(0.595 0)、新郑早红枣(0.594 4)、新星无核(0.587 4)、冬枣(0.585 5)、马铃枣(0.581 3)、葫芦枣(0.575 1)、金芒果枣(0.573 8)、早脆王(0.559 6)、晋枣(0.559 5)、冷白玉2(0.552 1)、灵武长枣(0.551 0)、小梨枣(0.549 9)、蜂蜜罐(0.543 7)、茶壶枣(0.539 7)、赞皇大枣(0.531 9)、胎里红(0.523 9)、晋赞大枣(0.517 4)、金谷大枣(0.517 4)、金昌1号(0.517 3)、骏枣(0.517 3)、壶瓶枣(0.514 4)、不落酥(0.504 0)、六月鲜(0.498 2)、敦煌大枣、小口枣、开口酥(0.494 2、0.494 2、0.494 2)、鸣山大枣(0.491 6)、晋矮3号(0.466 7)、大王枣(0.461 0)、晋矮4号(0.459 1)、临泽大枣(0.457 2)。遗传距离(GS)研究结果表明,黄河沿岸39份枣品种间的遗传差异性较大,遗传多样性丰富。
2.3 SSR聚类结果与分析
使用UPGMA法进行聚类分析,得到39份供试枣品种的遗传关系树状图(图 3)。在遗传相似系数0.660 0处,所有供试材料分为两大类群Ⅰ和Ⅱ。第Ⅰ类群只包括板枣,独立于其他供试材料,说明产于山西稷山的板枣[1,2,3]与其他供试品种亲缘关系较远,可能是独立起源的品种,与其他枣品种基因交流较少所致。
第Ⅱ类群在遗传相似系数0.696 0处分为4个亚群。
第1亚群包括悠悠枣、马牙白枣和磨盘枣。悠悠枣和马牙白枣的遗传相似系数为0.985 3,亲缘关系很近,这与它们的形态学观察一致,但两者原产地不同,悠悠枣原产于河北涿鹿[2],马牙白枣原产于北京[1,3]。
第2亚群只有赞皇大枣。赞皇大枣是河北省著名的土特产品,为自然三倍体[28]。图1和图2显示赞皇大枣为异源三倍体。
第3亚群包括灵武长枣(宁夏)、临泽大枣(甘肃)、晋矮4号(山西)、晋矮3号(山西)、鸣山大枣(甘肃)、六月鲜(山东)、敦煌大枣(甘肃)、小口枣(甘肃)、开口酥(甘肃)、不落酥(山西)、蜂蜜罐(陕西)、七月鲜(陕西)、金谷大枣(山西)、金昌1号(山西)、壶瓶枣(山西)、骏枣(山西)、晋枣(陕西、甘肃)等枣种质,原产于山西、陕西和甘肃的枣品种聚在了一起。
其中,晋矮4号和晋矮3号,鸣山大枣、敦煌大枣、小口枣和开口酥枣,骏枣、金谷大枣、金昌1号和壶瓶枣这3组试材分别在遗传相似系数0.993 0、0.995 5、0.953 0处各自相聚,其中金谷大枣和金昌1号的遗传相似系数是0.996 0,说明每组试材有极近的亲缘关系。鸣山大枣为敦煌大枣的变异单株[1],开口酥枣又名‘银枣 2 号’,是从甘肃小口枣中选育出的鲜食新品种[29];“金谷大枣”是从山西省太谷县优良种质资源中选育出的干鲜兼用新品种[30],金昌1号为山西省太谷县壶瓶枣的变异单株[2]。
第4亚群包括小梨枣(山东)、早脆王(山东)、新郑早红枣(河南)、金芒果枣(山东)、胎里红(河南)、孔府酥脆枣(山东)、葫芦枣(河南山西)、冷白玉1(北京)、尜尜枣(北京天津)、马铃枣(山东)、冬枣(山东河北)、新郑灰枣(河南)和新星无核(河北山东)等枣种质,原产于山东、河南和河北的枣品种聚在了一起。
其中,小梨枣和早脆王(0.988 0)聚在一起,两者亲缘关系很近。小梨枣是山东庆云[1,2]的早熟鲜食品种,枣果较小,早脆王是从山东庆云县引入山西的品种中选出的鲜食品种[2],两者果形枣核相似,果皮鲜红,完熟枣风味浓甜相同,是同一起源品种。马铃枣和尜尜枣在0.994 5处聚在一起,两者亲缘关系极近,表明它们起源相同。马铃枣产于山东陵县[1],而尜尜枣有原产北京的尜尜枣和天津尜尜枣之别[1,2]。根据形态学观察,来自林科院枣树示范园的“尜尜枣” 果形特征与天津尜尜枣相似,非北京的尜尜枣。
另外,宁夏灵武的灵武长枣[2,3],山西平遥等地的不落酥[1,3],陕西大荔等地的蜂蜜罐[1,2,3],陕西合阳的七月鲜[2],山东夏津等地的茶壶枣[1,2,3],陕西和甘肃的晋枣[1,2,3],河南新郑的新郑早红枣,河南镇平的胎里红[1,2,3],山东曲阜的孔府酥脆枣[1,2,3],北京的冷白玉1[2],山西河南各地的葫芦枣[1,3],河北和山东各地的冬枣[1,2],这12份试材在不同遗传相似系数(0.782 0、0.922 0、0.860、0.755 0、0.819 0、0.762 0、0.806 0、0.827 0、0.752 0、0.767 0、0.792 0、0.748 0)处各自独立为一支,说明它们彼此之间亲缘关系相对较远。
3 结论与讨论
利用20对SSR引物分析了39份枣品种的亲缘关系,结果表明,甘肃黄河沿岸枣品种间遗传差异较大,遗传多样性丰富。通过UPGMA聚类分析将39份枣品种分为两大类群Ⅰ和Ⅱ,第Ⅱ类群又分为4个亚群,聚类结果反映的品种间亲缘关系与其原产地有一定相关性。地理相邻的山西、陕西、宁夏和甘肃4省的枣类在0.735 0处聚在了一起,地理相邻的山东、河北和河南3省的枣类在0.724 0处聚在了一起(图3),这与前人研究结果基本一致,相同地理区域内亲缘关系近的枣品种被聚为一类[21,23]。本研究中,山西的晋矮4号和晋矮3号、山东庆云的小梨枣和早脆王等,均被聚到各自相同的类别。刘秀云等[22]用SSR分子标记分析255个枣品种亲缘关系,聚类和群体结构2种分析结果均表明,大部分枣品种的亲缘关系与来源区域基本相符。但枣亲缘关系与地理分布并非完全相同。六月鲜原产于山东宁阳[2,3],本研究中与甘肃的鸣山大枣(0.998 0)聚在一起。六月鲜与鸣山大枣表型差异较大,但遗传关系极近。新星无核是从河北沧州金丝小枣资源中选育出来的变异优良新品种[31],但是与河南新郑灰枣在0.927 0处聚在一起,说明它们有较近的亲缘关系。
通过SSR遗传分析,揭示了供试材料间的遗传多样性和复杂的亲缘关系,从分子水平厘清品种命名混乱的情况。本研究中,“大王枣”与晋矮4号、晋矮3号被聚为一类,说明“大王枣”与晋矮4号、晋矮3号起源相同,而不可能源于河南大王枣,应该是引种错误。“晋赞大枣”与壶瓶枣被聚为一类,图1和图2又显示赞皇大枣是三倍体,“晋赞大枣”是二倍体。而文献报道称晋赞大枣是从赞皇大枣变异优株中选育出的新品种[32],刘琰玮等[24]用16个位点的SSR标记分析晋赞大枣和赞皇大枣的亲缘关系,结果两者具有相同的基因型。这就说明本研究中的“晋赞大枣”与赞皇大枣非近亲品种,而是与壶瓶枣有极近亲缘关系的同源大果型品种。再如,本研究中的“冷白玉2”,果形特征与平川区枣树示范园的金芒果枣相似,两者的遗传相似系数是0.964 1。说明“冷白玉2” 和冷白玉1(北京)非近亲品种,而是与金芒果枣亲缘关系较近的冬枣品种。
本研究基于SSR标记技术从分子水平上揭示了甘肃黄河沿岸枣品种丰富的遗传多样性和复杂的亲缘关系,为有效解决枣品种命名混乱问题提供了依据,也为进一步研究和利用这些枣种质资源提供了依据。
