桂竹叶片解剖结构研究
2022-06-09贺沁文张亦嘉顾延生
贺沁文,郑 敏,张亦嘉,顾延生
(中国地质大学 环境学院,湖北 武汉 430078)
桂竹(Phyllostachysreticulata)为刚竹属的模式种[1]。刚竹属(Phyllostachys)隶属于禾本科(Gramineae)竹亚科(Bambusoideae),属下有约60种,为我国竹林最普遍的栽培种类[2]。该属研究近年来进展较大,但其属下关系有待澄清,与竹亚科其他属的关系尚存在争议[2]。研究表明,植物的解剖结构是研究植物系统分类的重要依据[3],分子生物学在应用于竹亚科系统发育研究时也需要形态学数据的补充和验证[4]。有关刚竹属植物的报道显示该属叶片的泡状细胞、短细胞、叶肉细胞、梭形细胞、气孔器、维管束、表皮毛、下表皮乳突等解剖结构的形态特征在不同种间存在变化,对揭示竹亚科植物的系统分类有重要意义[2,5-15]。其中,毛竹(Phyllostachysedulis)和黄槽竹(Phyllostachysaureosulcata)泡状细胞、气孔器、硅细胞等的形态,可作为竹类系统分类的依据[8];毛竹(Phyllostachysedulis)、刚竹(Phyllostachysviridis)、淡竹(Phyllostachysglauca)之间长细胞、短细胞、气孔器的形态特征存在差异[14];桂竹、毛竹、淡竹之间主脉维管束和梭形细胞的形态特征存在差异[15]。目前,刚竹属下叶片解剖结构被报道的种有5个,约占该属的10%,已有报道局限于少数形态指标,尚无全面的描述,刚竹属下各物种叶片解剖结构的共同特征为:梭形细胞不发达或无梭形细胞[15],短细胞单个或成对生长于脉上或脉间,气孔器集中分布于脉的每侧,乳突成列分布于下表皮长细胞表面,微毛分布于脉间,刺毛分布于脉间和脉上,大毛分布于脉上[16]。本研究对桂竹叶片解剖结构进行了全面的描述、测量及分析,建立了横切面结构与表皮结构的对应关系以填补本种叶解剖学研究的空白,并为桂竹及刚竹属的系统分类研究提供科学依据。此外,桂竹抗性较强,适生范围大[17],在我国黄河流域及其以南地区广泛分布,被大量用于建筑、纸张、食用、观赏等,为一重要经济种。我国拥有全世界最大规模的竹产业[18],桂竹为我国第二大竹材,用途之广仅次于毛竹[17],因而拥有重要的经济价值。目前对于桂竹的报道主要集中在基因组研究方面,桂竹的完整叶绿体基因组于2020年被初次报道[19],除此以外,对桂竹叶片的专门研究有限,尤其是在解剖学方面。本文对桂竹叶解剖结构进行详细描述,可为桂竹资源的进一步认识和可持续开发提供参考。
1 材料与方法
1.1 试验材料及仪器
桂竹叶片于2020年10月采自中国地质大学(武汉)未来城校区内(30°27′31″N,114°36′41″E)。选取长势良好的桂竹10株,从每株中部的不同枝条取完全舒展、无病虫害的成熟叶片4片。采摘后剪取叶片中部,立即置于FAA固定液中做固定处理。
主要仪器:Kedee KD-1508型切片机、烘箱、通风橱、Olympus BX51光学显微-拍摄系统。
1.2 样品制作方法
1.2.1 桂竹叶横切材料处理方法 取桂竹叶20片,置于25%氢氟酸中脱硅48 h[20];使用纯水漂洗1 d;经50%(1.5 h)、70%(1.5 h)、85%(1.5 h)、95%(1.5 h)、100%(0.5 h)叔丁醇梯度脱水及透明化处理;脱水及透明化后的材料浸入体积比为1∶1的石蜡-叔丁醇溶液,置于55 ℃烘箱中处理6 h,再转入体积比为3∶1的石蜡-叔丁醇溶液,置于57 ℃烘箱中处理6 h。烘干后的材料转入100%石蜡溶液,置于59 ℃烘箱中处理12 h,每4 h更换一次石蜡溶液。最后用石蜡包埋、Kedee KD-1508型切片机切片,切片厚度8~10 μm,中性树胶封片,Olympus BX51显微镜观察。
1.2.2 桂竹叶表皮材料处理方法 取桂竹叶20片,叶片上表面朝上置于载玻片上,使用蒸馏水润湿,用刀片轻轻刮去上表皮和叶肉组织,直至取得1 cm2完全透明而不破损的下表皮组织;后翻转叶片,使用同样方法在叶中部另一位置刮取1 cm2上表皮组织。甘油浸润后使用中性树胶封片,Olympus BX51显微镜观察。
1.3 参数测量方法
每个切片和刮片选取5个视野,在400倍显微镜下随机观察维管束、泡状细胞、长细胞、短细胞、表皮毛细胞、气孔、叶肉细胞、气腔、薄壁细胞、厚壁细胞。在所有切片和刮片中,对每级维管束各选取10个进行测量,对其他每类细胞各选取50个进行测量。其中,薄壁细胞、厚壁细胞在主脉和平行脉周围均有分布。在测量横切面高时,将这2种细胞近似视作圆形,不再因其分布位置加以区分。使用SPSS Statistics 26软件分析数据。
2 结果与分析
2.1 叶片表皮系统
2.1.1 上表皮 桂竹叶片上表皮细胞沿垂直于叶脉方向呈现“维管束-长(短)细胞-泡状细胞-长(短)细胞-维管束”的交替带状结构(图1A,图2A)。
在叶片上表面,泡状细胞带分布于2个长(短)细胞带之间,宽2~3行。泡状细胞呈现近圆形,其表面分布有少量矩形的硅质凸起(图1A:bc)。叶片横切面上,泡状细胞成组存在,每2个维管束之间分布1组(图1C、1D,图3)。每组有3个,中间一个体积较大,呈现典型的扇形,深入到叶肉内部,其两侧的泡状细胞呈现近矩形。
在叶片上表面,长细胞带位于维管束带的两侧,宽5~6行(图1A)。长细胞大致呈现矩形,角质层向外突起,突起长度约2 μm(图1A:lc)。在横切面上,长细胞呈现近圆形,分布于上表面泡状细胞和维管束之间(图1C、1D,图2)。短细胞成对或单个分布于2个长细胞之间。当成对出现时,向叶尖一侧的为硅细胞(图1A:sc1),向叶基一侧的为栓细胞(图1A:cc)。硅细胞呈现边缘不规则的近椭圆形,栓细胞呈现马鞍形。二者长宽相近。短细胞对数与长细胞个数之比约为1∶2。
复杂鞍形硅细胞生长于维管束近轴面和远轴面两侧的表皮上,两侧表皮上各分布有一行,呈现近矩形或哑铃形,(图1A:sc2,图1B:sc2)。复杂鞍形硅细胞广义上属于一种短细胞,本研究另列出以作区分。上表皮复杂鞍形硅细胞排列较为稀疏,每2个之间距离约10 μm。
桂竹叶的气孔器为横列型气孔器,在叶片上表皮分布稀疏,每个400倍视野(面积约84 800 μm2,下文亦同)下可见0~6个。气孔夹于2个长细胞之间生长,其长轴方向与长细胞相同。保卫细胞较为细小,形态特征不易观察。副卫细胞呈现典型的圆顶型(图2A)。
2.1.2 下表皮 由于不存在泡状细胞,下表皮呈现“维管束-长(短)细胞-维管束”的交替带状结构。与上表皮相比,下表皮细胞小而密集,表皮毛发达(图1B,图2C)。
在叶片下表面,长细胞分布于2个维管束之间,宽10~12行,长度波动较大。其角质层突起较上表皮长细胞更为明显,突起长度约3 μm(图1B:lc)。下表皮长细胞在远轴面一侧生长有5~6个乳突(图1C、1D:pp)。下表皮短细胞分布方式与上表皮相同,大小相近(图1B:sc1、cc)。下表皮短细胞数量远少于上表皮,短细胞对数与长细胞个数之比约为1∶10。
下表皮复杂鞍形细胞分布规律与上表皮基本相同(图1B:sc2),呈现哑铃形,大小与上表皮相接近。
桂竹下表皮细胞乳突发育密集,分布于长细胞表面,在下表皮呈现平行于维管的线形分布,排布较为规律(图1B:pp)。每2个乳突间距约3 μm,每个长细胞根据其大小生长有2~8个乳突。桂竹下表皮毛有大毛、刺毛2种。大毛密集分布于叶片下表面的脉上,每个400倍视野下可见约25个,长度变化幅度大(图1B:ma)。相较于分布在其他部位的大毛,分布在维管束附近的大毛更加粗壮(图1B:ma)。每根大毛的朝向一致,从叶基指向叶尖方向(图1E)。刺毛在下表面稀疏分布于脉间和脉上,每个400倍视野下可见3~5个(图1B:mi)。
下表皮气孔器分布方式与上表皮相近,数量远多于上表皮,400倍镜视野下可见约50个(图1B)。大小与上表皮接近。
2.2 叶片基本组织系统
2.2.1 叶肉细胞 桂竹叶肉细胞层数随部位变化。上表皮长细胞下方、气腔上方的叶肉最厚,约3~4层;泡状细胞下方的叶肉最薄,约2~3层(图2B)。靠近上表皮的第1层叶肉细胞排列最为规则,呈现略微弯曲的指状,第2~4层叶肉细胞排布较为不规则,呈现近放射状(图1D)。
2.2.2 梭形细胞 梭形细胞位于每个维管束两侧,无色透明,形态近于矩形(图1D)。
2.3 叶片维管组织系统
桂竹主脉位于叶片中央位置,其两侧各有6~7条一级平行脉,每2条一级平行脉之间有6~8条二级平行脉(图2)。桂竹的主脉和1级平行脉维管束具“V”字形结构。各级维管束鞘由一层较大的薄壁细胞和一层较小的厚壁细胞构成。维管束周围无“花环形”结构特征出现,为C3植物(图3)。
2.3.1 主脉 主脉在所有叶脉中最为发育,结构最为复杂,木质部和韧皮部最为分化。主脉与多个小型维管束形成复合维管系统,体积较大,横截面宽度约200 μm(图3:mv)。中央的一个维管束形态与一级平行脉近似,近轴面和远轴面处均有发达的机械组织与表皮相连,其余的小维管束鞘状结构不完整,薄壁细胞和厚壁细胞排列不规则。主脉两侧的梭形细胞最为发达,所形成的气腔结构体积最大。
2.3.2 一级平行脉 一级平行脉宽46.5~77.3 μm,木质部与韧皮部分化明显,维管束被薄壁细胞、厚壁细胞包围(图1C)。木质部发育有3个大型导管,近轴面一侧较小的导管为原生导管,远轴面一侧2个大小、形态相近的导管为后生导管,三者呈现“V”字形排列。近轴面一侧有少量机械组织与表皮相连(图3:1st pv)。
2.3.3 二级平行脉 二级平行脉宽27.7~31.4 μm,木质部与韧皮部分化不明显,木质部不发育大型导管(图1D)。维管束鞘细胞中,外侧的薄壁细胞较为发达,排列整齐,内侧的厚壁细胞数量较少。近轴面一侧有少量机械组织与表皮连接(图3:2nd pv)。
2.4 叶片解剖结构变异分析
对桂竹叶片的形态指标做统计分析,结果见表1~3。各个形态指标的变异系数差别较大,范围为4.9%~37.8%。其中,上表皮气孔、上表皮复杂鞍形细胞、二级平行脉的形态最为稳定,各项指标的变异系数均小于10%;下表皮长细胞、叶肉细胞的指标波动幅度最大,变异系数可达25%以上。通过对上、下表皮形态指标的对比得出,下表皮细胞的形态指标变异系数普遍大于上表皮细胞,说明桂竹叶片上表皮细胞形态较为稳定。

表1 桂竹上表皮形态指标统计Table 1 Morphological indices upper epidermis of P.reticulata

表2 桂竹下表皮形态指标统计Table 2 Morphological indices of lower epidermis of P.reticulata
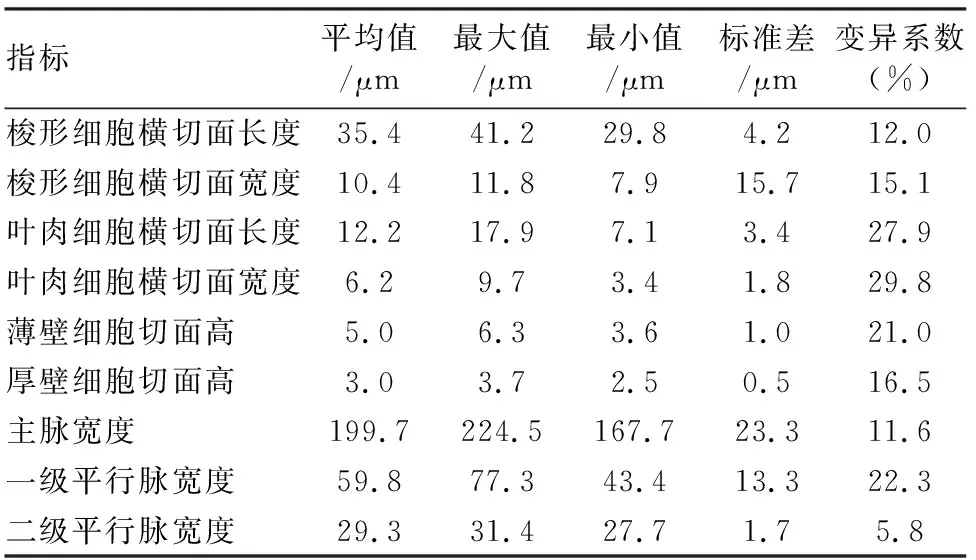
表3 桂竹叶肉及叶脉形态指标统计Table 3 Morphological indices of veins and mesophyll of P.reticulata
3 结论与讨论
桂竹是刚竹属的模式种,研究桂竹叶片的解剖结构,可以为研究其他刚竹属植物的系统分类和演化趋势提供参考。
桂竹叶片表面,长细胞间的短细胞多数以马鞍形栓细胞和椭圆形硅细胞成对的形式生长。国产17属33种竹材中,仅有刚竹属叶片同时存在椭圆形硅细胞与马鞍形栓细胞,为刚竹属的一个明显特征[14]。因此,上表皮马鞍形栓细胞和椭圆形硅细胞的成对出现可作为识别刚竹属的依据。对于生长于维管束两侧的短细胞,即复杂鞍形硅细胞,早期学者指出,其分布和形态是辨别竹亚科植物的重要特征[21]。禾本科各亚科植物泡状细胞形态学参数的概率分布函数显示,各亚科之间泡状细胞的形态参数存在显著性差异,因此泡状细胞的形态可作为分辨禾本科不同亚科植物的参考依据[6]。桂竹叶片表皮泡状植硅体的横向长度、纵向长度等形态学参数对于禾本科植物的分类定种有一定参考价值[22]。与竹亚科其他种的叶细胞相比,桂竹的梭形细胞窄而薄,叶肉细胞的分化水平较高(赵惠如等[8]认为,叶细胞的分化程度与梭形细胞的宽度、厚度呈负相关性。丁雨龙等[15]认为,桂竹叶片无梭形细胞,这与本研究的观察结果相矛盾。桂竹气孔的副卫细胞类型为圆顶型,这与桂竹的生境有关系。圆顶型副卫细胞是禾本科植物副卫细胞中较为进化的一类型,常于偏干、寒的温带环境中出现[23]。而圆顶型副卫细胞如何提高桂竹对偏干、寒环境的适应性,则有待进一步研究。桂竹的下表皮乳突呈现平行于维管的线形分布,这一分布方式有较强的规律性,为木本竹所特有[9],且在禾本科中为演化水平较高的物种所有[16,23]。近年,对于竹亚科筇竹属筇竹(Qiongzhueatumidinoda)[24]、慈竹属慈竹(Neosinocalamusaffinis)[25]叶解剖结构的全面描述亦被报道,将筇竹、慈竹、桂竹叶解剖结构做对比,得出桂竹叶表皮长细胞较筇竹、慈竹更发达;薄壁细胞、厚壁细胞较筇竹、慈竹不发达;泡状细胞大小与筇竹相近,较慈竹不发达;梭形细胞较筇竹不发达[24-25]。
综上所述,依照已有的研究结论,桂竹窄而薄的梭形细胞、分化水平较高的叶肉细胞、圆顶型的气孔副卫细胞、线形排布的下表皮乳突等特征均属于演化上比较进化的类型[8,16,23]。本文重点描述了桂竹叶片的表皮及横切面解剖结构,可用于桂竹的鉴定,为竹亚科的系统分类学研究提供资料。需要注意的是,本研究并未在不同竹种间做具体的解剖结构比较。将本研究结论应用于竹亚科的系统分类学时,有必要将桂竹叶解剖结构的观察结果、各个结构的变异分析结果与其他竹种进行比较研究[15]。
