广东省猪繁殖与呼吸综合征病毒ORF5 基因遗传变异分析
2022-06-07蒋智勇楚品品陈天宝李春玲蔡汝健
蒋智勇,楚品品,陈天宝,李春玲,蔡汝健
(广东省农业科学院动物卫生研究所/广东省畜禽疫病防治研究重点实验室/农业农村部兽用药物与诊断技术广东科学观测实验站,广东 广州 510640)
【研究意义】猪繁殖与呼吸综合征病毒(Porcine Reproductive and Respiratory Syndrome Virus,PRRSV)主要感染猪的呼吸系统和生殖系统,可引发高度传染性疾病,造成重大经济损失。PRRSV 自1987 年在美国发现以来,迅速蔓延至全世界频繁暴发。我国于1996 年分离到该病毒[1],2006 年夏秋季南方多个省区暴发高致病性PRRSV(HP-PRRSV)造成了大量猪只死亡,引起猪价大幅波动,对生猪产业的持续健康发展构成严重威胁。PRRSV 分子流行病学的调查与研究对制定以PRRSV 疫苗免疫和生物安全防控为基础的综合防控措施具有重要意义。
【前人研究进展】PRRSV 分为两个基因型,即欧洲型(PRRSV 1)和美洲型(PRRSV 2),它们感染猪的临床症状和病理变化相似,但抗原特性和遗传学特征却存在显著差异。目前我国的流行株主要为PRRSV 2。2010 年从广东地区使用MLV 疫苗的猪场中分离到属于Lineage 3 谱系毒株QYYZ 和GM2,重组分析表明GM2 是野毒株QYYZ 与疫苗毒株Resp/PRRS 的重组病毒[2]。2013 年周峰等[3]分离获得1 株Nsp2 区与2008年美国学者分离的NADC30 株[4]具有相同缺失特征的毒株,命名为NADC30-like,此后该毒株逐渐成为我国北方地区的主要流行毒株。2014 年以来我国报道了基因组出现明显变异和易重组的NADC30-like 毒株[5-6]。分子流行病学研究发现,我国流行的PRRSV 可分为4个谱系:Lineage 8(JXA1-like 和CH-1a-like)、Lineage 5(VR-2332-like)、Lineage 3(QYYZ-like)和Lineage 1(NADC30-like)[7]。
【本研究切入点】PRRSV 为单股正链RNA病毒,基因组长度约为 15 kb,其长度变化主要决定于Nsp2 区域缺失与插入片段长度。ORF2-7以亚基因组形式编码病毒的8 种结构蛋白,其中以GP5 变异最大,是PRRSV 最主要的保护性抗原蛋白,因此在病毒分子遗传变异分析中ORF5基因常作为基因分型依据用于 PRRSV 分子流行病学监测[8]。【拟解决的关键问题】为了监测PRRSV 在广东省的流行动态和进化趋向,本研究2021 年采集了广东地区规模猪场临床疑似PRRS症状的猪只组织样品,通过对ORF5基因测序进行遗传变异分析,探查广东省内各地区流行毒株的分布情况,为制定PRRSV 防控措施提供科学依据。
1 材料与方法
1.1 试验材料
病料为521 份肺脏组织样品,采于2021 年广东省不同地区37 个规模猪场疑似PRRS 症状的仔猪和流产胎儿。
病毒DNA/RNA 提取试剂盒(Magen)购自广州美基生物科技有限公司;PrimeScript™One Step RT-PCR Kit 购自宝生物工程(大连)有限公司(TaKaRa)。
1.2 引物设计
参照GenBank 中不同谱系PRRSV 代表株基因组序列设计1 对特异性引物扩增ORF5基因,具体序列如下,ORF5-F:5'-CATTTCATGACACCTGAGAC-3',ORF5-R:5'-CGGCATCTGGAGGTGATGA-3',预期扩增长度为970 bp,引物由生工生物工程(上海)股份有限公司合成。
1.3 PRRSV ORF5 基因的RT-PCR 扩增与测序
分别取少量肺脏加入500 μL PBS 匀浆、离心取200 μL 上清按照病毒DNA/RNA 提取试剂盒说明书提取病毒RNA,然后按照一步法RT-PCR 试剂盒配制RT-PCR 反应体系并进行RT-PCR 扩增,反应结束后将电泳检测为阳性的PCR 产物送生工生物工程(上海)股份有限公司测序。
1.4 PRRSV ORF5 基因及编码蛋白的序列比对与遗传进化树构建
对双向测序结果截取完整的ORF5基因,用DNAStar 与GenBank 中的16 株国内外代表株(表1)作为参考毒株,对其基因序列以及推导的氨基酸序列进行比对。使用ClustalX 2.1 和Mega 7.0软件基于 Neighbor-Joining 方法绘制系统遗传进化树。
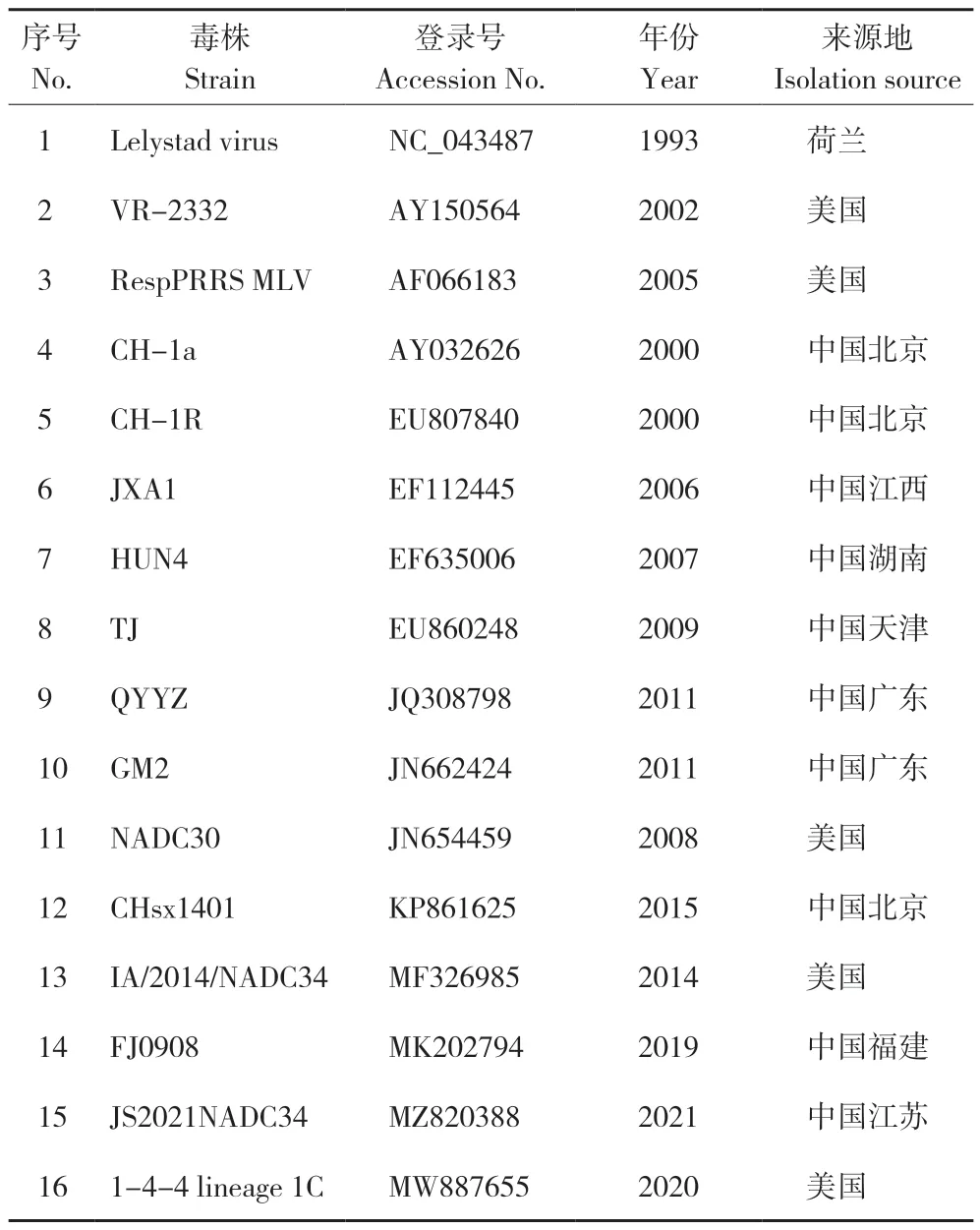
表1 PRRSV 代表毒株信息Table 1 Information of representative PRRSV strains
2 结果与分析
2.1 PRRSV ORF5 基因RT-PCR 扩增及序列测定
对供试的37 个猪场521 份疑似PRRS 症状的组织样品提取病毒RNA,经RT-PCR 扩增后,产物电泳结果见图1,扩增产物约1 000 bp,与预期大小一致。其中检出PRRSV 阳性样品127 份,来自17 个猪场,样品阳性率为24.4 %,猪场阳性率为45.9%。每个阳性样品双向重复测序2 次,测序结果表明同一阳性猪场的阳性样品测序结果一致,获得了来自17 个不同猪场的PRRSVORF5基因序列,其中有3 株(GD MM、GD FS 和GD LN)在同一位置缺失3 个碱基,其他ORF5基因全长均为603 bp,并根据样品来源地进行命名。

图1 PRRSV ORF5 基因的RT-PCR 产物Fig.1 RT-PCR products of PRRSV ORF5 gene
2.2 PRRSV ORF5 基因序列遗传进化分析
将17 株PRRSVORF5基因序列与表1 的16 株PRRSV 参考序列用Mega 7.0 构建遗传进化树,结果(图2)显示,17 株PRRSV 分别属于PRRSV 2 的3个谱系。Lineage 1 谱系分为2 个分支(NADC30-like 和NADC34-like),分别包含了近几年美国流行的ORF5RFLP 1-4-4 代表株1-4-4 lineage 1c 和ORF5RFLP 1-7-4 代表株IA/2014NADC34。本研究得到的8 株PRRSV(GD YC、GD FS、GD MM、GD YJ、GD ZQ、GD GZ、GD LN 和GD JM)属于 Lineage 1(NADC30-like)谱系,1株PRRSV(GD MZ)属于 Lineage 1(NADC34-like)谱系;3 株PRRSV(GD HY、GD HZ、和GD ZJ)属于 Lineage 3 谱系(QYYZ-like);5 株PRRSV(GD JM2、GD TS、GD YF、GD QY 和GD SH)属于Lineage 8(JXA1-like 和 CH-1a-like)谱系。17 株PRRSV 中Lineage 1、Lineage 8 和Lineage 3占比分别为52.94%、29.41%和17.65%,没有检测到Lineage 5(VR2332-like)谱系的毒株。
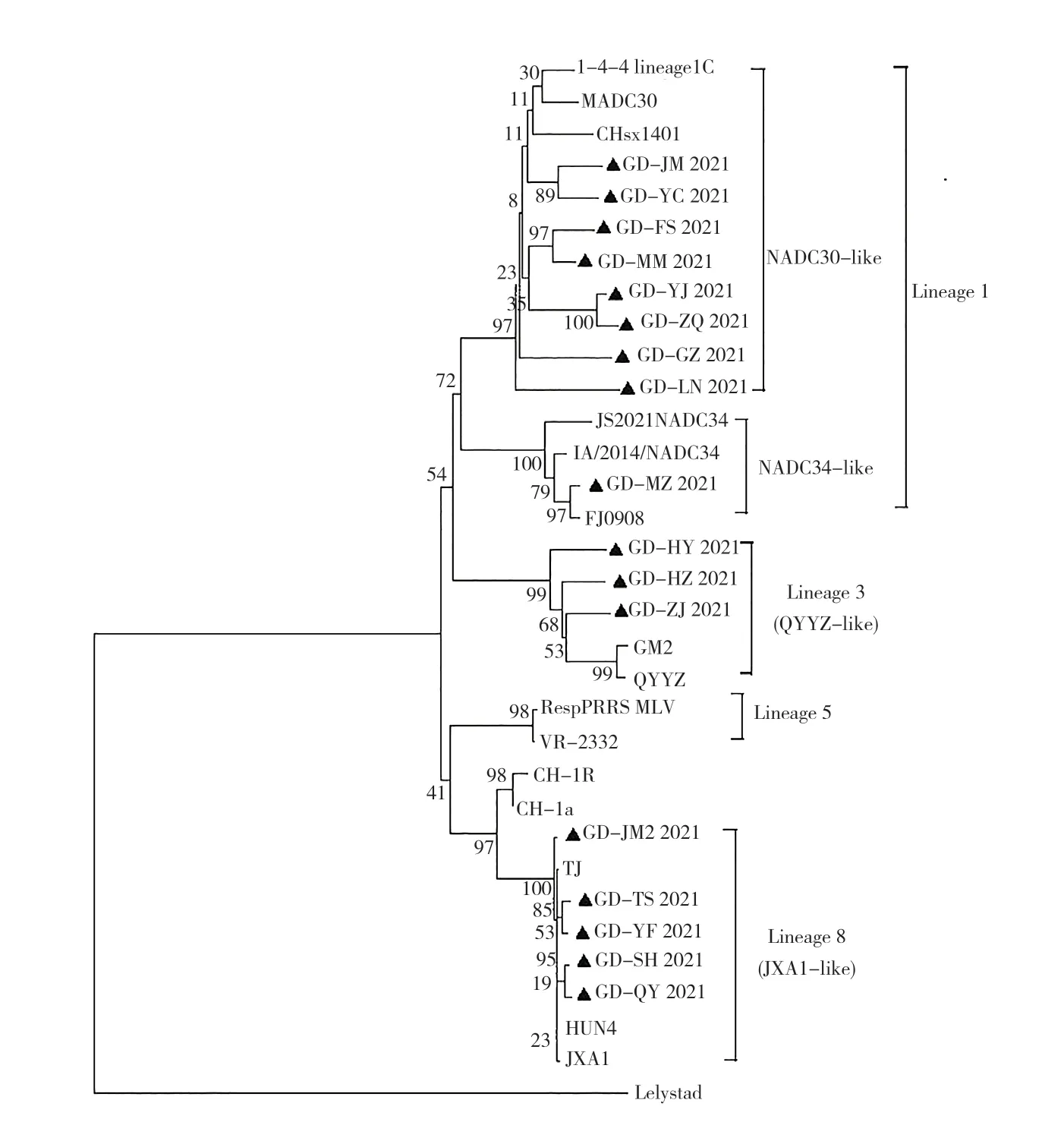
图2 PRRSV 基于ORF5 基因的系统遗传进化树Fig.2 Phylogenetic tree of PRRSV based on ORF5 gene
采用DNAstar 对17 株PRRSVORF5基因及参考序列进行核苷酸同源性分析,结果(图3)显示,ORF5基因核苷酸序列同源性为81.3%~99.5%;与HP-PRRSV 中的JXA 1 株、Hu4 株、TJ 株核苷酸同源性为81.1%~98.5%;与美国VR-2332 株的核苷酸同源性为83.1%~89.7%;与欧洲型LV 株的核苷酸同源性为61.8%~64.3%。属于 Lineage 1 (NADC30-like 和 NADC34-like) 谱系的9 株分离株之间的核苷酸同源性为84.8%~95.8%;属于Lineage 3(QYYZ-like)谱系的3 株分离株之间核苷酸同源性为92.9%~94.4%;属于Lineage 8(JXA1-like 和 CH-1a-like)谱系的5 株分离株之间核苷酸同源性为97.8%~99.5%,与JXA1 株核苷酸同源性为98.7%~99.5%。

图3 PRRSV ORF5 基因核苷酸序列同源性分析结果Fig.3 Results for homology analysis of nucleotide sequence of PRRSV ORF5 gene
2.3 PRRSV ORF5 基因推导的氨基酸序列分析
通过DNAstar 分析ORF5基因编码蛋白GP5 推导的氨基酸序列同源性,结果(图4)显示,17 株PRRSV GP5 氨基酸序列同源性为78.5%~98.5%;与Lineage 1 中NADC30 株的氨基酸同源性为84.6%~94.5%;与Lineage 3 中GM2 株的氨基酸同源性为77.5%~93.5%;与Lineage 5 中VR-2332株的氨基酸同源性为80.5%~88.1%;与Lineage 8中JXA 1 株、Hu4 株、TJ 株氨基酸序列同源性为83.0%~98.5%;与欧洲型LV 株的氨基酸同源性为50.7%~57.7%。属于 Lineage 1 (NADC30-like 和NADC34-like) 谱系的9 株分离株之间的氨基酸同源性为85.0%~95.0%;属于Lineage 3(QYYZlike)谱系的3 株分离株之间的氨基酸同源性为92.9%~94.4%。

图4 PRRSV ORF5 基因编码蛋白GP5 推导的氨基酸序列同源性分析结果Fig.4 Results for homology analysis of the deduced amino acid sequence of protein GP5 encoded by PRRSV ORF5 gene
ORF5基因编码蛋白GP5 推导的氨基酸序列比对结果如图5 所示,有3 株(GD MM、GD FS和GD LN)缺失了第33 位氨基酸,其氨基酸突变位点主要位于GP5 的信号肽(Signal peptide)、中和位点(PNE)、2 个高变区(HVR)、B 细胞抗原位点(B cell epitop)和2 个T 细胞抗原位点附近[9]。GP5 的N-糖基化位点在病毒感染、免疫逃避方面起重要作用,GP5 有2 个保守的糖基化位点(第44、51 位)[10],而位于高变区的第32、33、34、35 位N-糖基化位点容易突变,HP-PRRSV 第32 位突变为S,而第35 位又由S突变为N;Lineage 3 谱系第33 位N 突变为G。有些氨基酸位点的突变具有谱系特异性,Lineage 1 特异的突变位点有第47 位L 突变I、第57 位A突变N/R/D、第168 位E 突变D 和第192 位V 突变I;HP-PRRSV 特异的氨基酸位点第9、13、24、25、32、35、161、189、191 位分别为C、R、Y、L、S、N、V、L 和R;Lineage 3 特异的氨基酸位点第34、38、39、66、92、170 位分别为G、Y、S、C、S 和G。

图5 推导的GP5 氨基酸序列比对分析Fig.5 Alignment analysis of the deduced amino acid sequence of GP5
3 讨论
PRRSV 的聚合酶复制保真度差、点突变、重组和免疫压力选择促进PRRSV 不断演化,被认为是产生病毒变异性和多样性的潜在分子机制。自2006 年我国首次报道HP-PRRSV 毒株(以JXA1 为代表)以来,在随后的几年中,JXA1-like 毒株成为我国流行的优势毒株。但随着2010年和2013 年在我国大陆分别出现QYYZ-like 和NADC30-like 毒株,RPRSV 的多样性显著扩大,使得PRRS 的防控日趋复杂。同时多种弱毒活疫苗在全国范围内广泛使用,无疑加大了猪群的免疫选择压力,外来新毒株的入侵以及与疫苗株间重组进一步加剧了PRRSV 的多样化。
广东省内PRRSV 优势流行毒株与我国北方地区不一致,2014—2016 年从广东省内分离的毒株测序结果表明以Lineage 8(JXA1-like 和CH-1a-like)谱系和 Lineage 3(QYYZ-like)谱系为主[11];2019—2020 年监测结果表明广东省内PRRSV 以Lineage 3(QYYZ-like)谱系为主,其次为Lineage 8(JXA1-like 和CH-1a-like)谱系和 Lineage 1(NADC30-like)谱系[12]。本研究对2021 年广东省内不同猪场的PRRSV 测序分析结果显示,有52.94%(9/17)属于Lineage 1(NADC30-like 和NADC34-like)谱系、29.41%(5/17)属 于Lineage 8(JXA1-like 和CH-1a-like)谱 系和17.65%(3/17)属于Lineage 3(QYYZ-like)谱系。由此可见,2021 年广东省内PRRSV 流行毒株以Lineage 1 谱系为主,Lineage 3 谱系在广东省内流行比前几年大幅缩小,流行毒株的变化可能与近年来种猪的频繁调运有关。从临床PRRS症状检测属于Lineage 8(JXA1-like 和CH-1alike)谱系的毒株核苷酸序列与JXA1 株同源性为98.7%~99.5%,可能与疫苗使用不当有关,疫苗免疫以弱毒疫苗免疫后再以灭活疫苗加强免疫可以获得较好的免疫效果[13]。
2014 年首次报道在美国至少5 个州检测到RFLP 1-7-4 PRRSV,其代表毒株为IA/2014/NADC34 株,感染仔猪后可引起严重的临床症状[14]。2017 年在我国辽宁省首次监测到NADC34-like PRRSV[15],随后在黑龙江、福建、江苏和四川均有报道[16-19]。本研究从广东梅州地区分离的毒株GD MZ 2021 株ORF5基因具有RFLP1-7-4 模式特点,与IA/2014/NADC34 株同源性为97.3%,且NSP2 区域有100 aa 的连续缺失,还需要通过全基因组测序进一步验证。
2020 年10 月以来,美国猪场暴发由 PRRSV 1-4-4 Lineage 1C 新型变异株引起的疫情,该毒株具有更强的传播能力和致病性,且现有疫苗对其保护效果不佳[20]。美国PRRSV 1-4-4 Lineage1C 变异株与我国PRRSV NADC34-like 属于同一谱系分支(Lineage 1),其Nsp2 区域存在100 aa 连续缺失。鉴于NADC34-like 已成为美国主要流行株,有可能成为我国PRRSV 潜在流行株[21]。为了有效防控PRRS,需要对PRRSV 流行株分子变异进行持续监测,以期为临床诊断、新型疫苗的研发提供参考。
4 结论
为了监测广东省2021 年PRRSV 的分子流行动态,从省内生猪主产区采集37 个规模猪场521份疑似PRRS 患病猪的组织样本进行PRRSV 检测。对阳性样品进行ORF5基因的RT-PCR 扩增和测序,获得17 株来自不同地区规模猪场的PRRSVORF5基因序列并进行遗传进化分析,结果表明,所有毒株均属于PRRSV 2,在遗传进化树上分别属于3 个谱系即Lineage 1(NADC30-like和NADC34-like)谱系、Lineage 8(JXA1-like 和CH-1a-like)谱系和 Lineage 3(QYYZ-like)谱系,所占比例分别为52.94%、29.41%和17.65%,没有检测到Lineage 5(VR2332-like)谱系的毒株。与前几年的检测数据对比发现,2021 年广东地区Lineage 1 (NADC30-like)谱系的PRRSV 占比快速上升,而Lineage 3(QYYZ-like)谱系占比下降。17 株PRRSV 毒株ORF5基因之间核苷酸序列同源性为81.3%~99.5%,推导的GP5 氨基酸序列同源性为80.0%~98.3%,GP5 高变区的糖基化位点存在不同程度变异。
