朱红毛斑蛾和花胸姬蜂对虫害诱导榕树挥发物的触角电位及行为反应
2022-06-07管维康王小云陆温张志林郑霞林
管维康 王小云 陆温 张志林 郑霞林




摘要 朱紅毛斑蛾Phauda flammans是一种寡食性害虫,仅取食小叶榕Ficus concinna和垂叶榕F.benjamina。花胸姬蜂Gotra octocinctus是其幼虫的优势寄生性天敌。研究朱红毛斑蛾及花胸姬蜂对朱红毛斑蛾为害小叶榕和垂叶榕后诱导产生的挥发物的触角电位及行为反应可为实施“推-拉”策略防控朱红毛斑蛾提供基础资料。基于气相色谱-质谱联用(gas chromatography-mass spectrum, GC-MS)和气相色谱-触角电位联用(gas chromatography-electroantennagraphic detection, GC-EAD),从受朱红毛斑蛾幼虫为害的小叶榕和垂叶榕虫害叶挥发物中筛选出对朱红毛斑蛾和花胸姬蜂雌雄成虫有电生理活性的物质,基于Y形嗅觉仪,测定朱红毛斑蛾和花胸姬蜂雌雄成虫对虫害叶挥发物成分的行为反应。结果表明,朱红毛斑蛾为害小叶榕和垂叶榕后,虫害叶中共分离出57种挥发物成分,其中,共有27种能引起朱红毛斑蛾雌雄虫的触角电位反应,在小叶榕和垂叶榕虫害叶片挥发物中同时存在的有11种。其中,两种榕树虫害叶特有的5种挥发物(β-月桂烯、β-罗勒烯、(-)-α-蒎烯、β-石竹烯和α-金合欢烯)均能引起花胸姬蜂雌雄成虫的触角电位反应。与健康叶挥发物混合物相比,受朱红毛斑蛾幼虫为害后释放的挥发物成分混合物对朱红毛斑蛾和花胸姬蜂雌雄成虫均具有更强的吸引作用。结果提示在生产上可使用朱红毛斑蛾为害后诱导的榕树挥发物成分混合物实施“推-拉”防控策略。
关键词 虫害诱导挥发物;朱红毛斑蛾;花胸姬蜂;触角电位;行为反应
中图分类号: S763.3
文献标识码: A
DOI: 10.16688/j.zwbh.2021115
Abstract Phauda flammans (Walker) (Lepidoptera: Phaudidae) is an oligophagous pest, which only feeds on Ficus concinna Miq. and F.benjamin L. Gotra octocinctus Ashmead is the dominant parasitoid of P.flammans larvae. Understanding the function of plant volatiles induced by P.flammans larvae from the two Ficus species can provide the basis for implementation of the “push-pull” control strategy against this moth. Gas chromatography-mass spectrum (GC-MS) and gas chromatography-electroantennographic detection (GC-EAD) were used to screen electrophysiological active substances from infested leaves of F.concinna and F.benjamina. Based on the Y olfactometer, the behavioral responses of male and female adults of P.flammans and G. octocinctus to plant volatiles from infested leaves were measured. A total of 57 compounds were isolated from F.concinna and F.benjamina leaves after damaged by P.flammans larvae. Male and female P.flammans exhibited electroantennographic responses to 27 compounds, of which 11 compounds were shared by F.concinna and F.benjamina. The five volatiles (β-myrcene, β-ocimene, (-)-α-copaene, β-caryophyllene and α-farnesene) specific to the leaves of two Ficus species damaged by P.flammans larvae could elicit electroantennographic response of male and female G. octocinctus. Compared with the volatile mixture from healthy Ficus leaves, the mixture from damaged leaves showed stronger attraction to both male and female adults of P.flammans and G. octocinctus. The results suggested that the volatile mixtures from damaged leaves of the two Ficus species have the potential for implementing the “push-pull” control strategy against P.flammans.
Key words herbivore-induced plant volatiles;Phauda flammans;Gotra octocinctus;electroantennogram;behavioral response
植物-植食性昆虫-天敌三者经过长期的协同进化,不仅形成了以能量流动为基础的食物链关系,还形成了以化学信息为基础的复杂的种间联系[1-2]。其中,植物挥发物在三者间的关系中发挥了重要作用。一方面,植食性昆虫可利用寄主植物挥发物进行寄主定位[3-5];另一方面,寄主植物受植食性昆虫为害后,也会释放能吸引昆虫前去取食、产卵、滞留的化学信息物质,这类物质可被植食性昆虫利用,以搜寻寄主植物,也可被天敌利用,以定位寄主昆虫生境[1, 6]。研究这三者间的关系,既能为深入探索其协同进化奠定基础,也可为研发新的害虫防控策略提供技术支撑,具有重要的理论和应用价值。
朱红毛斑蛾 Phauda flammans(Walker)(鳞翅目Lepidoptera毛斑蛾科Phaudidae)是一種寡食性害虫,幼虫仅以小叶榕Ficus concinna Miq.和垂叶榕F.benjamina L.叶片为食[7],在南亚和东南亚国家已有其暴发为害的报道[8-10]。在我国主要分布于广西、广东、福建和海南等地[11-14]。其低龄幼虫啃食叶肉,留下白色膜状上表皮,而高龄幼虫取食后造成叶片缺刻或孔洞,甚至将整株叶片全部食光,食物短缺时还会取食嫩枝韧皮部,造成枝干枯死,连续多年为害可导致整株榕树死亡[11],严重影响城市景观、生态效益和社会效益。目前,针对该虫的研究主要集中在各虫态外部形态特征[11]、成虫和幼虫触角感器类型及分布[15-16]、生殖系统构造[17]等形态学方面;食性[7]、发生规律[11-14]、越冬场所[18]、发育起点温度和有效积温[19]、抗寒力[20]和潜在地理分布[21]等生物生态学方面;化学药剂[22-23]、核型多角体病毒[24]、性信息素[25]、球孢白僵菌[26-28]和寄生性天敌[29]对其防效等防控技术方面。作者前期研究结果表明,小叶榕和垂叶榕健康叶中的D-柠檬烯、β-罗勒烯和β-石竹烯是吸引朱红毛斑蛾雌雄虫寄主定位的主要植物挥发物活性成分[30]。然而,当朱红毛斑蛾取食为害小叶榕和垂叶榕后,寄主植物释放的主要植物挥发物活性成分及其含量是否会发生改变及其对朱红毛斑蛾和主要寄生性天敌花胸姬蜂Gotra octocinctus Ashmead(膜翅目Hymenoptera姬蜂科Ichneumonidae)寄主定位的影响均尚不明确。作者假设小叶榕和垂叶榕受朱红毛斑蛾为害后释放的挥发物在朱红毛斑蛾和花胸姬蜂寄主定位中发挥积极作用。为验证该假设,采用动态顶空吸附法收集小叶榕和垂叶榕虫害叶片释放的挥发物,并通过气相色谱-质谱联用(gas chromatography-mass spectrum,GC-MS)、气相色谱-触角电位联用(gas chromatography-electroantennographic detection,GC-EAD)和Y形嗅觉仪技术,研究了朱红毛斑蛾和花胸姬蜂对小叶榕和垂叶榕虫害诱导挥发物的触角电位和行为反应。
1 材料与方法
1.1 试验材料
1.1.1 供试虫源
2019年5月,在广西壮族自治区南宁市东葛路(22°49′ N,108°20′ E)垂叶榕上采集朱红毛斑蛾幼虫,带回室内置于塑料盒(直径10 cm,高 15 cm)内,每盒10头,在室温条件下饲养,每日更换新鲜的垂叶榕叶片直至化蛹。根据茅裕婷等[17]的方法鉴别雌雄蛹,并分开保存在不同的塑料盒内(直径10 cm,高 15 cm),置于室温下。待成虫羽化后,随机选取1日龄健康的未交配雌雄成虫作为供试虫源。
将野外采集的朱红毛斑蛾幼虫在室内用垂叶榕叶片饲养,直至化蛹,每日09:00、15:00和21:00观察是否有花胸姬蜂羽化,随机选取1日龄健康的花胸姬蜂未交配雌雄成虫作为供试虫源。
1.1.2 仪器与试剂
气相色谱-质谱联用仪(7890B-5975C,安捷伦科技有限公司,美国)、气相色谱-触角电位联用仪(7890B-IDAC4,安捷伦科技有限公司,美国;Syntech公司,德国)、Y形嗅觉仪(自制,主臂长30 cm,侧臂长20 cm,主臂和侧臂内径4 cm,侧臂间夹角70°)。
D-柠檬烯(CAS号:7705-14-8;纯度90%;百灵威科技有限公司,中国)、β-月桂烯(CAS号:123-35-3;纯度98%;成都埃法生物科技有限公司,中国)、β-罗勒烯(CAS号:3779-61-1;纯度90%;南宁国拓生物科技有限公司,中国)、(-)-α-蒎烯(CAS号:3856-25-5;纯度95%;成都埃法生物科技有限公司,中国)、β-石竹烯(CAS号:87-44-5;纯度98%;成都埃法生物科技有限公司,中国)、α-金合欢烯(CAS号:502-61-4;纯度99%;上海伊卡生物技术有限公司,中国)。
1.2 试验方法
1.2.1 小叶榕和垂叶榕虫害叶片挥发物的收集与鉴定
选取广西大学校园(22°50′ N,108°17′ E)内长势良好、未被朱红毛斑蛾为害且未喷洒化学药剂的小叶榕和垂叶榕植株作为供试植物,用网罩(长 × 宽=50 cm × 50 cm)将5头4~5龄饥饿处理12 h的朱红毛斑蛾幼虫限制在垂叶榕或小叶榕侧枝顶端约20片叶片上,取食12 h后取出幼虫,作为虫害叶处理,并采用动态顶空吸附法收集虫害叶挥发物。
收集挥发物前,使用聚对苯二甲酸乙二酯无味透明塑料袋套住榕树顶端虫害叶片,袋的两端留进气口和出气口,使用聚四氟乙烯管连接,进气流量为300 mL/min,采气流量为250 mL/min。进气端连接1.0 g吸附剂(PoraPak QS 80-100)过滤空气,采气端装有与进气端同等型号的吸附剂吸附植物挥发物。采集时间为9:30~15:30,同时采集3组,为3个重复,采集结束后用2.5 mL的色谱纯正己烷洗脱吸附管,并将洗脱液保存于-18℃低温冰箱,检测前在通风橱中用N2吹扫,挥发浓缩至100 μL,待用。
GC-MS检测条件:色谱柱为HP-5毛细管柱(30 m × 0.25 mm × 0.25 μm),载气为高纯氦气(99.99%),进样量为2 μL,恒流不分流模式,流速为1 mL/min;柱箱升温程序:起始温度50℃,保持1 min,5℃/min升温至95℃,1℃/min升温至99℃,5℃/min升温至115℃,1℃/min升温至119℃,5℃/min升温至170℃,15℃/min升温至250℃结束;EI离子源,电子能量70 eV,质子扫描范围50~550 amu,四极杆温度为150℃,离子源温度为230℃。各成分通过核对标准谱库(NIST 11.0)标准化合物的质谱图进行校对,参考夏尚文等[31]报道的榕树挥发物成分,确定各色谱峰所对应的物质,根据峰面积归一法进行定量。
1.2.2 朱红毛斑蛾和花胸姬蜂对榕树虫害叶挥发物的GC-EAD反应
测试样品为小叶榕和垂叶榕植株虫害叶释放的挥发物洗脱液。参照马艳等[32]的方法,用手术剪将待测试虫的触角从基部剪下,用昆虫针挑起触角,将其两端连接在涂有导电胶的电极上,将触角放置在距离气味管出口约1 cm处,持续气流为300 mL/min。气相色谱的条件同1.2.1。色谱柱出口接分流器,分流比为1∶1,分流的化合物一部分进入氢焰离子化检测器(FID),另一部分经过恒温加热套管吹向触角,利用GC-EAD软件,同步记录气相色谱和触角电位图,通过对化合物的峰形和保留时间进行对比,鉴定出朱红毛斑蛾雌雄虫触角产生电位反应的物质。每头成虫仅使用1根触角,每根触角仅测试1次,重复测试10根触角。
参照夏尚文等[31]的研究对挥发物进行初步筛选。选取对朱红毛斑蛾雌雄成虫具有GC-EAD反应的榕树虫害叶特有的挥发物成分,用正己烷分别稀释到10-4,研究朱红毛斑蛾雌雄成虫对其的GC-EAD反应。花胸姬蜂对榕树虫害叶挥发物的GC-EAD反应研究方法同朱红毛斑蛾。柱箱升温程序:起始温度50℃,保持1 min,再以20℃/min升温至250℃结束。每头成虫仅使用1根触角,每处理重复3次。
1.2.3 朱红毛斑蛾和花胸姬蜂对榕树虫害叶挥发物成分混合物的行为反应
将小叶榕和垂叶榕特有的5种虫害叶挥发物活性物质参照其相对含量按照一定比例混合,作为处理混剂A(β-月桂烯∶β-罗勒烯∶(-)-α-蒎烯∶β-石竹烯∶α-金合欢烯=1∶100∶10∶100∶100,m/m);健康叶中5种活性物质根据其相对含量按照一定比例混合,作为对照混剂B(D-柠檬烯∶β-月桂烯∶β-罗勒烯∶ (-)-α-蒎烯∶β-石竹烯=1∶1∶100∶10∶100,m/m)[30]。参照江丽容等[33]和Roy等[34]的方法,利用Y形嗅觉仪研究朱红毛斑蛾及花胸姬蜂雌雄成虫对虫害叶挥发物的行为反应。测试时,用10 μL移液枪吸取5 μL 混剂A和混剂B分别滴在1 cm × 2 cm 的定性滤纸上,并分别置于两臂连接球形陷阱中,以混剂A为处理,混剂B为对照。气体流速控制在300 mL/min。每次仅释放1头朱红毛斑蛾成虫,试验时先于引虫口适应30 s后释放,每头成虫观察5 min,进入侧臂一半以上则计为选择,否则记为不选择并更换成虫重新测试。每10头为1组,重复3组。每测试5头,调换两侧管臂位置,以消除环境影响。每测试1组,用98%无水乙醇清洗装置后,用蒸馏水冲洗并烘干后使用。花胸姬蜂对榕树虫害叶挥发物成分混合物行为反应的研究方法同朱红毛斑蛾。
1.3 数据统计
使用SPSS 19.0软件进行数据统计。小叶榕和垂叶榕健康/虫害叶挥发物相对含量间的比较及朱红毛斑蛾和花胸姬蜂对虫害叶挥发物成分混合物的行为反应均采用独立样本t测验检验(P < 0.05)。统计分析前,百分率数据均进行了平方根转换。
2 结果与分析
2.1 小叶榕和垂叶榕虫害叶片挥发物的收集与鉴定
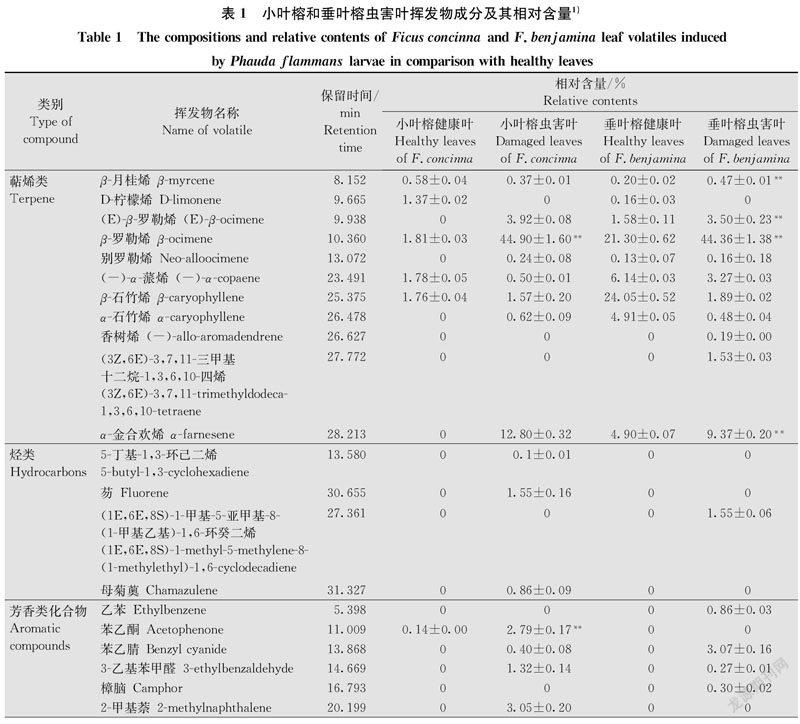
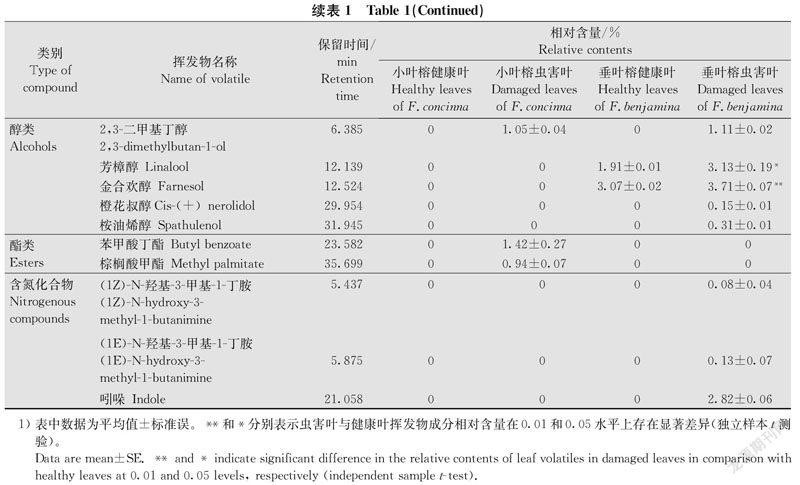
从小叶榕和垂叶榕虫害叶中共分离出57种挥发物成分,其中小叶榕虫害叶中分离出33种,垂叶榕虫害叶中分离出37种,两种榕树虫害叶挥发物中相同成分为14种。与小叶榕健康叶挥发物成分相比,小叶榕虫害叶中新增挥发物成分14种;虫害叶相对含量较高的是β-罗勒烯和α-金合欢烯,分别占挥发物总量的(44.90±1.60)%和(12.80±0.32)%;虫害叶中β-罗勒烯(t=26.967,df=4,P < 0.01)和苯乙酮(t=15.432,df=4,P < 0.01)的相对含量显著增加(表1)。
与垂叶榕健康叶挥发物成分相比,垂叶榕虫害叶中新增挥发物成分13种;虫害叶相对含量较高的是β-罗勒烯和α-金合欢烯,分别占挥发物总量的(44.36±1.38)%和(9.37±0.20)%;虫害叶中β-月桂烯(t =12.211,df=4,P < 0.01)、(E)-β-罗勒烯(t=7.703,df=4,P < 0.01)、β-罗勒烯(t=15.234,df=4,P < 0.01)、芳樟醇(t=6.487,df=4,P < 0.05)、金合欢醇(t=9.307,df=4,P < 0.01)和α-金合欢烯(t=21.254,df=4,P < 0.01)的相对含量显著增加(表1)。
2.2 朱红毛斑蛾和花胸姬蜂对榕树虫害叶挥发物的GC-EAD反应
朱红毛斑蛾雌雄虫对小叶榕和垂叶榕虫害叶片挥发物有触角电位反应的物质共有27种,其中小叶榕21种(图1),垂叶榕18种(图2),相同物质有11种。有触角电位反应的27种物质分别为甲苯、乙苯、对二甲苯、邻二甲苯、蒎烯、β-月桂烯、(E)-β-罗勒烯、β-罗勒烯、苯乙酮、壬醛、芳樟醇、金合欢醇、别罗勒烯、3-乙基苯甲醛、4-乙基苯甲醛、萘、水杨酸甲酯、4-异丙基苯甲醇、3-乙基苯乙酮、4-乙基苯乙酮、吲哚、(-)-α-荜澄茄油烯、(-)-α-蒎烯、β-荜澄茄油烯、β-石竹烯、α-金合欢烯和石竹素(图1,图2)。相同的11种物质分别为乙苯、β-月桂烯、(E)-β-羅勒烯、β-罗勒烯、3-乙基苯甲醛、4-异丙基苯甲醇、3-乙基苯乙酮、4-乙基苯乙酮、(-)-α-蒎烯、β-石竹烯和α-金合欢烯。
花胸姬蜂雌雄成虫对受朱红毛斑蛾幼虫为害的小叶榕和垂叶榕叶片特有的挥发物成分β-月桂烯、β-罗勒烯、(-)-α-蒎烯、β-石竹烯和α-金合欢烯均有明显的GC-EAD反应(图3)。
2.3 朱红毛斑蛾和花胸姬蜂对虫害叶挥发物成分混合物的行为反应
与健康叶挥发物成分混合物相比,虫害叶挥发物成分混合物对朱红毛斑蛾雌虫(t=5.657,df=4,P < 0.01)和雄虫(t=2.828,df=4,P < 0.05)均
有显著的吸引作用(图4)。
与健康叶挥发物成分混合物相比,花胸姬蜂雌虫(t=7.071,df=4,P < 0.01)和雄虫(t=2.828,df=4,P < 0.05)对受朱红毛斑蛾为害后榕树特有的5种挥发物成分混合物具有显著的趋向行为反应(图5)。
3 讨论
本研究结果表明,与小叶榕和垂叶榕健康叶片挥发物相比,受朱红毛斑蛾为害后其释放的挥发物对朱红毛斑蛾及其天敌花胸姬蜂的吸引力更强,说明虫害诱导的榕树挥发物在朱红毛斑蛾和花胸姬蜂的寄主定位中发挥积极作用,支持本文提出的假设。
虫害诱导挥发物有助于植食性昆虫的寄主定位。虫害诱导挥发物的成分主要是萜类化合物,且机械损伤不能诱导这些物质的产生[35]。作者前期研究明确了榕树健康叶挥发物的成分及朱红毛斑蛾雌雄成虫对其的行为反应[30]。本研究发现,受朱红毛斑蛾取食为害后,小叶榕和垂叶榕叶片挥发物中的β-罗勒烯等萜烯类物质的含量显著增加,并增加了新的挥发物α-金合欢烯,且虫害诱导的挥发物成分混合物对朱红毛斑蛾雌雄虫的吸引效果显著高于健康叶挥发物成分混合物。对于幼虫活动能力较弱的蛾类而言,找不到合适的寄主植物的代价可能是饿死或被天敌捕食。所以,鉴于幼虫搜索寄主能力较弱和新寄主植物适口性不确定的双重因素,部分蛾类成虫更喜欢选择受同种类为害的植株产卵[36]。如,苹果蠹蛾Cydia pomonella(L.)[37]和草地贪夜蛾Spodoptera frugiperda(J. E. Smith)[36]。
寄生性天敵对寄主昆虫的搜寻通常取决于寄主昆虫和与寄主相关的环境等因素。通常情况下,大多数寄主昆虫以聚集形式分布即“斑块”分布,而不是均匀或随机分布[38]。因此,相比于随机搜索,利用化学信号搜索猎物的效率要更高。就信号可信度而言,来自于寄主昆虫的化学信息物质最为可靠,而植物所释放的挥发物的可信度要相对较低,但对寄主昆虫释放的化学信号其识别距离通常较短,一般用于近距离精准定位,而植物所释放化学信号通常量较大,更易吸引天敌在寄主生境滞留[1-2]。
对于寄生蜂雌蜂而言,如何利用寄主植物挥发物来准确定位寄主,是其种群繁衍的关键因素之一。本研究中,与榕树健康叶的挥发物相比,朱红毛斑蛾幼虫为害诱导的挥发物对花胸姬蜂雌虫有显著的引诱效果,可能是由于小叶榕和垂叶榕经朱红毛斑蛾幼虫取食后,其挥发物成分以及相对含量发生了变化,导致其“化学指纹图谱”发生了改变,从而更利于花胸姬蜂识别寄主。其他类群昆虫的研究也得到相类似的结果。例如,受小菜蛾Plutella xylostella为害的卷心菜Brassica oleracea var. capitata L.和胡萝卜Daucus carota L. var. sativa Hoffm.挥发物更容易吸引其天敌菜蛾盘绒茧蜂Cotesia vestalis(Halliday)雌蜂[39];与健康桑Morus alba L. 枝相比,受桑天牛Apriona germari Hope为害后桑树枝条挥发物更容易吸引桑天牛的寄生性天敌长尾啮小蜂Aprostocetus fukutai Miwa et Sonan雌蜂,其吸引作用甚至比桑天牛的卵更强[40]。本研究还发现虫害诱导的挥发物对花胸姬蜂雄虫也有显著的引诱效果。寄主植物挥发物对于寄生蜂雄蜂来说也有重要的生态学意义,因为其是雄蜂定位雌蜂从而能顺利完成两性交配的关键因素[41]。例如,中红侧沟茧蜂Microplitis mediator(Haliday)雄蜂主要依靠被甘蓝夜蛾Mamestra brassicae L.为害的芜青Brassica rapa L.挥发物来定位配偶[42]。对于花胸姬蜂雄虫而言,可能朱红毛斑蛾幼虫为害诱导的榕树挥发物在其觅偶过程中发挥了引导作用。
研究表明,植物挥发物在寄生蜂远距离寄主定位生境过程中发挥主要作用。而对其寄主的近距离定位,仍需多种化学物质的共同作用,如寄主唾液[43]、虫粪挥发物[44]、虫体挥发物[45]和卵表面化合物[46]等,才能显著提高寄生蜂定位寄主的准确度与效率。本文仅研究了寄主植物挥发物对花胸姬蜂行为反应的影响,有关寄主植物挥发物对花胸姬蜂寄主定位的影响及朱红毛斑蛾的卵、幼虫和虫粪等挥发物对花胸姬蜂行为反应和寄主定位的影响尚不明确;这些化合物在其寄主定位过程中是否存在协同作用也需进一步研究。
总之,受朱红毛斑蛾幼虫为害后的榕树叶片挥发物更容易吸引朱红毛斑蛾及其天敌花胸姬蜂。说明虫害诱导的榕树挥发物在榕树-朱红毛斑蛾-花胸姬蜂系统中,对植食性昆虫和寄生蜂的寄主定位均具有积极的作用。在生产上可使用朱红毛斑蛾为害后诱导的榕树挥发物成分混合物,“拉”来朱红毛斑蛾成虫集中诱杀,或“拉”来花胸姬蜂,发挥其自然天敌的控害作用,从而减轻或避免榕树受害。然而,榕树挥发物对于朱红毛斑蛾和花胸姬蜂的调控机制尚不明确,还有待进一步的深入研究。此外,已有研究表明昆虫性信息素通常和寄主植物挥发物协同作用于植食性昆虫的寄主定位[47-48],对朱红毛斑蛾而言,虽然已明确了其性信息素组分及诱捕效果[25],但其是否会和榕树挥发物协同作用,从而更有利于朱红毛斑蛾或花胸姬蜂的寄主定位,也需深入研究。
参考文献
[1] TURLINGS T C J, ERB M. Tritrophic interactions mediated by herbivore-induced plant volatiles: Mechanisms, ecological relevance, and application potential [J]. Annual Review of Entomology, 2018, 63: 433-452.
[2] TAKABAYASHI J, SHIOJIRI K. Multifunctionality of herbivory-induced plant volatiles in chemical communication in tritrophic interactions [J]. Current Opinion in Insect Science, 2019, 32: 110-117.
[3] 钦俊德, 王琛柱. 论昆虫与植物的相互作用和进化的关系[J]. 昆虫学报, 2001, 44(3): 360-365.
[4] BRUCE T J, WADHAMS L J, WOODCOCK C M. Insect host location: a volatile situation [J]. Trends in Plant Science, 2005, 10(6): 269-274.
[5] DUDAREVA N, NEGRE F, NAGEGOWDA D A, et al. Plant volatiles: Recent advances and future perspectives [J]. Critical Reviews in Plant Sciences, 2006, 25(5): 417-440.
[6] 王琛柱, 钦俊德. 昆虫与植物的协同进化: 寄主植物-铃夜蛾-寄生蜂相互作用[J]. 昆虫知识, 2007, 44(3): 311-319.
[7] 刘俊延, 马仲辉, 吴塞逸, 等. 朱红毛斑蛾嗜食性的研究[J]. 环境昆虫学报, 2016, 38(5): 924-930.
[8] NAGESHCHANDRA B K, RAJAGOPAL B K, BALASUBRAMANIAN R. Occurrence of slug caterpillar Phauda flammans (Walker) (Lepidoptera: Zygaenidae) on Ficus racemosa L. in South India [J]. Mysore Journal of Agricultural Science, 1972, 6(2): 186-189.
[9] VERMA T D, DOGRA G S. Occurrence of Phauda flammans (Walker) (Lepidoptera: Zygaenidae) on Ficus species in Himachal Pradesh [J]. Journal of Tree Science, 1982, 1(1/2): 130-132.
[10]ROBINSON G S, ACKERY P R, KITCHING I J, et al. Hosts-a database of the world’s Lepidopteran hostplants [EB/OL]. [2021-02-26].The Natural History Museum, UK, 2010. http:∥www.nhm.ac.uk/hosts.
[11]刘俊延, 何秋隆, 魏航, 等. 朱红毛斑蛾生物学特性研究[J]. 植物保护, 2015, 41(3): 188 -192.
[12]何桂玲, 廖富林. 朱红毛斑蛾危害梅州市区榕树的研究[J]. 嘉应学院学报, 2015, 33(11): 72-74.
[13]黄志嘉, 茅裕婷, 朱映, 等. 朱红毛斑蛾在广东地区的爆发程度调查[J]. 河北林业科技, 2018(3): 47-49.
[14]林奇田, 林卉, 谢长炜, 等. 厦门地区朱红毛斑蛾的发生情况[J]. 亚热带植物科学, 2020, 49(4): 307-311.
[15]LIU Junyan, ZHANG Yujing, HUANG Zongyou, et al. Ultrastructural observations of antennal sensilla in Phauda flammans (Walker) (Lepidoptera: Zygaenidae) [J]. Journal of Entomological Science, 2018, 53(3): 281-294.
[16]劉俊延, 黄宗优, 张玉静, 等. 朱红毛斑蛾幼虫头部感受器扫描电镜观察[J]. 植物保护学报, 2018, 45(6): 1314-1320.
[17]茅裕婷, 贾蕊娟, 朱诚棋, 等. 朱红毛斑蛾雌雄鉴别与生殖系统解剖观察[J]. 浙江林业科技, 2017, 37(6): 87-92.
[18]刘俊延, 何秋隆, 苏莎, 等. 榕树食叶害虫朱红毛斑蛾蛹的调查方法[J]. 中国植保导刊, 2014, 34(6): 51-53.
[19]刘俊延, 何秋隆, 魏航, 等. 室内自然变温下朱红毛斑蛾发育历期、起点温度和有效积温[J]. 植物保护, 2015, 41(1): 137-140.
[20]ZHENG Xialin, LIU Junyan, HUANG Zongyou, et al. Cold hardiness of Phauda flammans (Lepidoptera: Zygaenidae) larvae [J]. Entomologica Fennica, 2017, 28(1): 9-15.
[21]卢小雨, 蒋露, 叶奕优, 等. 基于MaxEnt模型的朱红毛斑蛾在我国的潜在地理分布[J]. 环境昆虫学报, 2019, 41(6): 1268-1275.
[22]蒋露, 王林聪, 郭强, 等. 四种药剂对朱红毛斑蛾幼虫室内药效测定[J]. 中国森林病虫, 2018, 37(6): 41-44.
[23]HUANG Ailing, MENG Lingyu, ZHANG Wei, et al. Effects of five pesticides on toxicity, detoxifying and protective enzymes in Phauda flammans (Walker) (Lepidoptera: Zygaenidae) [J]. Pakistan Journal of Zoology, 2019, 51(4): 1457-1463.
[24]刘清浪, 吴若光, 曾陈湘. 朱红毛斑蛾核型多角体病毒[J]. 广东林业科技, 1985(4): 30.
[25]ZHENG Xialin, LIU Junyan, ZHANG Zhilin, et al. Diel rhythms of sexual behavior and pheromone responses in Phauda flammans (Walker) (Lepidoptera: Zygaenidae) [J]. Pest Management Science, 2019, 75(11): 3070-3075.
[26]陈学敏, 王小云, 陆温, 等. 球孢白僵菌对朱红毛斑蛾幼虫蛋白质含量、保护酶和解毒酶活力的影响[J]. 植物保护学报, 2020, 47(6): 1379-1380.
[27]陈学敏, 王小云, 陆温, 等. 4种虫生真菌对朱红毛斑蛾幼虫的室内毒力测定[J]. 广西植保, 2020, 33(2): 1-5.
[28]CHEN Xueming, WANG Xiaoyun, LU Wen, et al. Use of Beauveria bassiana in combination with commercial insecticides to manage Phauda flammans (Walker) (Lepidoptera: Phaudidae): Testing for compatibility and synergy [J]. Journal of Asia-Pacific Entomology, 2021,24(2): 272-278.
[29]ZHENG Xialin, LI Jun, SU Li, et al. Ecological and morphological characteristics of parasitoids in Phauda flammans (Lepidoptera: Zygaenidae) [J/OL]. Parasite, 2015, 22: 36. DOI: 10.1051/parasite/2015036.
[30]管维康, 王小云, 陆温, 等. 朱红毛斑蛾对榕树挥发物的触角电位及行为反应[J]. 森林与环境学报, 2020, 40(4): 442-448.
[31]夏尚文, 陳进. 昆虫取食和人工损伤处理对五种榕树挥发物释放的影响[J]. 云南植物研究, 2007, 29(6): 694-700.
[32]马艳, 史黎央, 赵艺, 等. 星天牛不同为害状态下山核桃挥发物成分的GC-EAD和行为反应[J]. 应用昆虫学报, 2019, 56(3): 530-538.
[33]江丽容, 刘守安, 韩宝瑜, 等. 7种寄主和非寄主植物气味对茶尺蠖成虫行为的调控效应[J]. 生态学报, 2010, 30(18): 4993-5000.
[34]ROY N, BARIK A. Long-chain free fatty acids from sunflower (Asteraceae) leaves: Allelochemicals for host location by the arctiid moth, Diacrisia casignetum Kollar (Lepidoptera: Arctiidae) [J]. Journal of the Kansas Entomological Society, 2014, 87(1): 22-36.
[35]娄永根, 程家安. 虫害诱导的植物挥发物: 基本特性、生态学功能及释放机制[J]. 生态学报, 2000, 20(6): 1097-1106.
[36]CARROLL M J, SCHMELZ E A, MEAGHER R L, et al. Attraction of Spodoptera frugiperda larvae to volatiles from herbivore-damaged maize seedlings [J]. Journal of Chemical Ecology, 2006, 32(9): 1911-1924.
[37]LANDOLT P J, BRUMLEY J A, SMITHHISLER C L, et al. Apple fruit infested with codling moth are more attractive to neonate codling moth larvae and possess increased amounts of (E, E)-alpha-farnesene [J]. Journal of Chemical Ecology, 2000, 26(7): 1685-1699.
[38]WILLIS A J, BEGON M, HARPER J L, et al. Ecology: individuals, populations and communities [J]. Bioscience, 1996, 38(3): 424-424.
[39]YONEYA K, UEFUNE M, TAKABAYASHI J. Parasitoid wasps’ exposure to host-infested plant volatiles affects their olfactory cognition of host-infested plants [J]. Animal Cognition, 2018, 21(1): 79-86.
[40]仇兰芬, 孙绪艮, 孙守家, 等. 桑天牛长尾啮小蜂的寄主搜索行为[J]. 昆虫天敌, 2003, 25(1): 24-27.
[41]XU Hao, TURLINGS T C. Plant volatiles as mate-finding cues for insects [J]. Trends in Plant Science, 2018, 23(2): 100-111.
[42]XU Hao, DESURMONT G, DEGEN T, et al. Combined use of herbivore-induced plant volatiles and sex pheromones for mate location in braconid parasitoids [J]. Plant Cell and Environment, 2017, 40(3): 330-339.
[43]BABENDREIER D, BIGLER F, KUHLMANN U. Methods used to assess nontarget effects of invertebrate biological control agents of arthropod pests [J]. Biological Control, 2005, 50(6): 821-870.
[44]楊元, 张凤娟, 李继泉, 等. 桑枝和桑天牛虫粪挥发物对桑天牛长尾啮小蜂的引诱作用[J]. 蚕业科学, 2007, 33(3): 360-366.
[45]李伟. 蚧虫虫体挥发物对天敌瓢虫的引诱效应[D]. 太原: 山西大学, 2016.
[46]李继泉, 王树香, 杨元, 等. 桑天牛长尾啮小蜂产卵及寄主识别行为的观察与研究[J]. 蚕业科学, 2006, 32(4): 447-452.
[47]REDDY G V P, GUERRERO A. Interactions of insect pheromones and plant semiochemicals [J]. Trends in Plant Science, 2004, 9(5): 253-261.
[48]FANG Yulei, ZENG Rui, LU Shufang, et al. The synergistic attractiveness effect of plant volatiles to sex pheromones in a moth [J]. Journal of Asia-Pacific Entomology, 2018, 21(1): 380-387.
(责任编辑:杨明丽)
