昼夜温度波动对麦长管蚜生理生态表型影响及其相关性研究
2022-06-03张树明张建珍
邢 鲲,张树明,赵 飞*,张建珍
(1. 山西大学应用生物学研究所,太原 030006;2. 山西农业大学植物保护学院,太原 030031)
近几年,全球气候变化现象日益显著,不仅平均温度增加明显,而且昼夜或季节性温度波动幅度也增加明显(Vázquezetal., 2017)。温度波动幅度(temperature fluctuation)作为气候变化主要特征之一,日益受到生物学及生态学家关注。已有研究表明,温度波动影响着多种生物种群的生理、生态、适应性,甚至对一些物种进化都产生显著影响(Xingetal., 2019; 2021a;Noeretal., 2020),如植物(Quisthoudtetal., 2012)、微生物(Longoetal., 2017)、鱼类(Oligny-Hébertetal., 2015)、蛇类(Geistetal., 2011)、龟类(Micheli-Campbelletal., 2012)、等。
对温度敏感的昆虫而言,变温幅度对昆虫的生态特征、生理特点、分子机理等影响尤为明显。从生态角度来说,较大变温幅度能够抑制小菜蛾Plutellaxylostella卵、幼虫的发育,降低卵与幼虫的存活率(Xingetal., 2015; 2021b);减少麦长管蚜Sitobionavenae寿命(Zhaoetal., 2014);降低地中海果蝇Ceratitiscapitata生殖量(Terblancheetal., 2010),甚至降低黑腹果蝇Drosophilamelanogaster种群内禀增长率(Clavijo-Baquetetal., 2014)。从生理角度来说,较大变温幅度可以降低黑腹果蝇温度耐受性范围(Bozinovicetal., 2016),提高地中海果蝇热耐受性(Terblancheetal., 2010);降低埃及伊蚊Aedesaegypti传毒能力(Lambrechtsetal., 2011);从分子机理角度来说,较大变温幅度提高了Drosophilagaucha热激蛋白的表达量(Franciscaetal., 2016);影响黑腹果蝇体内热相关基因表达,并且这种改变是独立于对平均温度变化的反应(Srensenetal., 2016)。
麦长管蚜Sitobionavenae(Fabricius)作为小麦种植区主要害虫之一,广泛分布于东非、亚洲、欧洲、北美等世界各个地区(Winderetal., 2010)。麦长管蚜由于体型小、热传导快、代谢速率高,对环境温度变化非常敏感(Zhaoetal., 2014),是研究环境温度变化影响的理想物种。大部分麦长管蚜与温度的相关性研究,主要以恒温梯度变化为主(尹青云等, 2003; 许乐园等, 2014),也有一些研究表明短时高温或波动温度对麦长管蚜的发育、存活(Zhaoetal., 2014)、寿命与繁殖(Zhaoetal., 2019)、热耐受性(Zhaoetal., 2017)、种群动态(Caoetal., 2018)等有显著的影响。目前,麦长管蚜对变温幅度的生态学响应研究比较有限,并且尚无变温幅度对麦长管蚜温度耐受性与呼吸代谢方面的研究。充分了解变温幅度对麦长管蚜生理生态表型的影响,以及两者之间的相关性,不仅可以为气候变化下害虫发生与为害的风险评估提供指导,还能为合理的制定害虫综合防控措施提供数据支撑。
为此,本研究选取平均温度为22℃下3种变温幅度(±0℃、±6℃、±12℃),研究在模拟自然界24 h波动温度中变温幅度对麦长管蚜发育、存活、寿命、繁殖、种群参数以及热耐受性、冷耐受性、呼吸速率等方面的影响,以期全面阐明气候变化背景下昼夜变温幅度对麦长管蚜生理与生态学的影响。
1 材料与方法
1.1 供试昆虫
2016年5月山西省农科院小麦试验田(116°16′N,35°55′E)采集麦长管蚜。采回后将麦长管蚜置于养虫室内(温度20±1℃,相对湿度50%~60%,光周期16 L ∶ 8 D)进行饲养。先将麦长管蚜放置在10~20 cm冬小麦幼苗(每周更换一次)上进行饲养,然后将冬小麦放置于养虫笼内(60 cm×60 cm×60 cm),最后养虫笼置于养虫室内。试验前麦长管蚜在室内已经继代饲养至少4年。
1.2 温度设置
本研究模拟2012-2020年中国北方小麦产区(临汾)小麦快速生长时期5月份的气温数据。通过气象数据进一步分析发现,临汾地区5月份小麦生长期内日均温在22℃左右,昼夜变温幅度最大为±12℃。因此,本研究选取平均温度为22℃,昼夜温差分别设置为±0℃、±6℃和±12℃(图1-A)。采用人工气候箱内模拟昼夜温度变化。试验过程中,人工气候箱内使用Datalogger(U23-001, Hobo Ltd., USA)记录温度,并且每20 min自动记录1次人工气候箱内实际温湿度(图1-B)。人工气候箱内光暗周期为固定的L ∶ D=16 ∶ 8,其中每天05 ∶ 00-21 ∶ 00为光照阶段,21 ∶ 00-05 ∶ 00为黑暗阶段,湿度控制在RH 40%~60%。实验室内温度由中央空调控制在22℃左右。

图1 试验设计温度(A)与连续三天内不同变温幅度处理下实际记录温度(B)Fig.1 Experimental design temperature (A) and actual recorded temperature in three consecutive days under different temperature fluctuations (B)
1.3 相关试验测定
生活史测定:选取4 h内新生若蚜144头,采用饲养管(Caoetal., 2018)进行单头饲养,分别置于22±0℃、22±6℃、22±12℃人工气候箱内进行饲养,每处理48头新生若蚜。每日08 ∶ 00检查蚜虫个体死亡及产仔情况,并将死亡蚜虫及新生若蚜去除,直至供试蚜虫全部死亡,试验调查结束。为保证充足的营养供应,饲养管内小麦叶片每隔2 d更换1次。
温度耐受性测定:(1)临界高温(critical maximum temperature,CTmax)测定,在22±0℃、22±6℃、22±12℃人工气候箱内,随机选取9日龄的体型大小基本一致的成蚜,放入Huber(Ministat230-cc-NR,德国)(图2-A)的外接反应釜中。先将成蚜(每处理30头)单个放入蜂窝板(长度×宽度:80×80 mm)的小孔(直径 5 mm,深度5 mm)中,两侧分别由尼龙纱布(200目)覆盖,防止蚜虫逃逸。将蜂窝板垂直放置于反应釜内,反应釜内温度在22℃平衡5 min后,温度从22℃以0.5℃/min的速率上升到30℃,然后以0.1℃/min上升到40℃,停止温度程序。随着温度升高,反应釜内蚜虫会出现不能正常行走、痉挛、附肢突然停止颤动,最终蚜虫昏迷。CTmax值即反应釜内蚜虫附肢停止颤动时的温度(Zhaoetal., 2017)。(2)过冷却点(supercooling point,SCP)及冰点(freezing point,FP)测定,在22±0℃、22±6℃、22±12℃人工气候箱内,随机选取9日龄的体型大小基本一致的成蚜。采用热电偶测定法进行测定,每处理30头蚜虫。因成蚜体较小,温度探头断插入自制固定器内,将成蚜放入固定器并与热敏电阻探头充分接触,用脱脂棉轻轻固定,然后将固定器快速放入低温-35℃冰箱,此时,成蚜体温迅速降低。热敏电阻开始显示成蚜体表温度的变化,观察并记录仪成蚜过冷却点与冰点温度。
呼吸代谢测定:在22±0℃、22±6℃、22±12℃人工气候箱内,随机选取9日龄的体型大小基本一致的成蚜用于呼吸速率测定。每处理30头成蚜。采用呼吸测定仪(Li-7000,美国)(图2-B),将成蚜按照不同处理分别放入第2到第4个呼吸室(第1个呼吸室作为基线)内。成蚜在呼吸室内呼吸1 h后开始测量,成虫呼出的CO2进入CO2流速分析仪,并用Expedata软件进行呼吸分析。试验在室温度下(约22℃)进行,重复3次。

图2 温度耐受性测定仪器(A)与呼吸测定仪器(B)Fig.2 Temperature tolerance instrument (A) and respiration instrument (B)
1.4 统计分析
利用Shapiro-Wilk检验与Levene检验分别检验数据的正态性与方差均质性,发现大部分数据不符合正态分布且方差不齐。因此,采用广义线性模型(GLM)中Gamma分布模型分析变温幅度(单因素)对麦长管蚜发育、寿命、繁殖、温度耐受性、呼吸速率等性状的影响,不同变温幅度处理间采用LSD进行多重比较。麦长管蚜存活采用Kaplan-Meier进行生存曲线分析,并采用Log Rank法进行不同处理间差异显著性比较,生存分析中去除非正常死亡与逃逸蚜虫数据,剩余数据均在SPSS 20.0中进行分析,并用Graphpad 9.0作图。种群参数,即每种处理的净繁殖率(R0)、世代周期(G)和内禀增长率(Rm)由以下方程计算:
其中x是麦长管蚜的日龄,Lx是麦长管蚜在x日龄时的存活率比例,Mx是每头麦长管蚜的生殖总量。采用R中bootstrap procedure方法分析麦蚜种群内禀增长率的平均值以及95%置信区间,并且不同变温幅度处理之间采用R的“agricolae”数据包中“Kruskal”函数进行Kruskal-Wallis多重比较。结果以平均值±标准差(Mean±SD)表示,P<0.05为差异显著。
2 结果与分析
2.1 不同变温幅度对麦长管蚜生理表型的影响
2.2 不同变温幅度对麦长管蚜生态表型的影响


图3 不同变温幅度对于麦长管蚜成蚜临界高温、过冷却点、冰点和呼吸速率的影响Fig.3 Effect of different temperature fluctuations on CTmax, SCP, FP and respiration of Sitobion avenae adult注:图中柱上不同小写字母表示不同处理间差异显著(P<0.05)。Note: Different letters above the bar indicated significant differences (P<0.05).
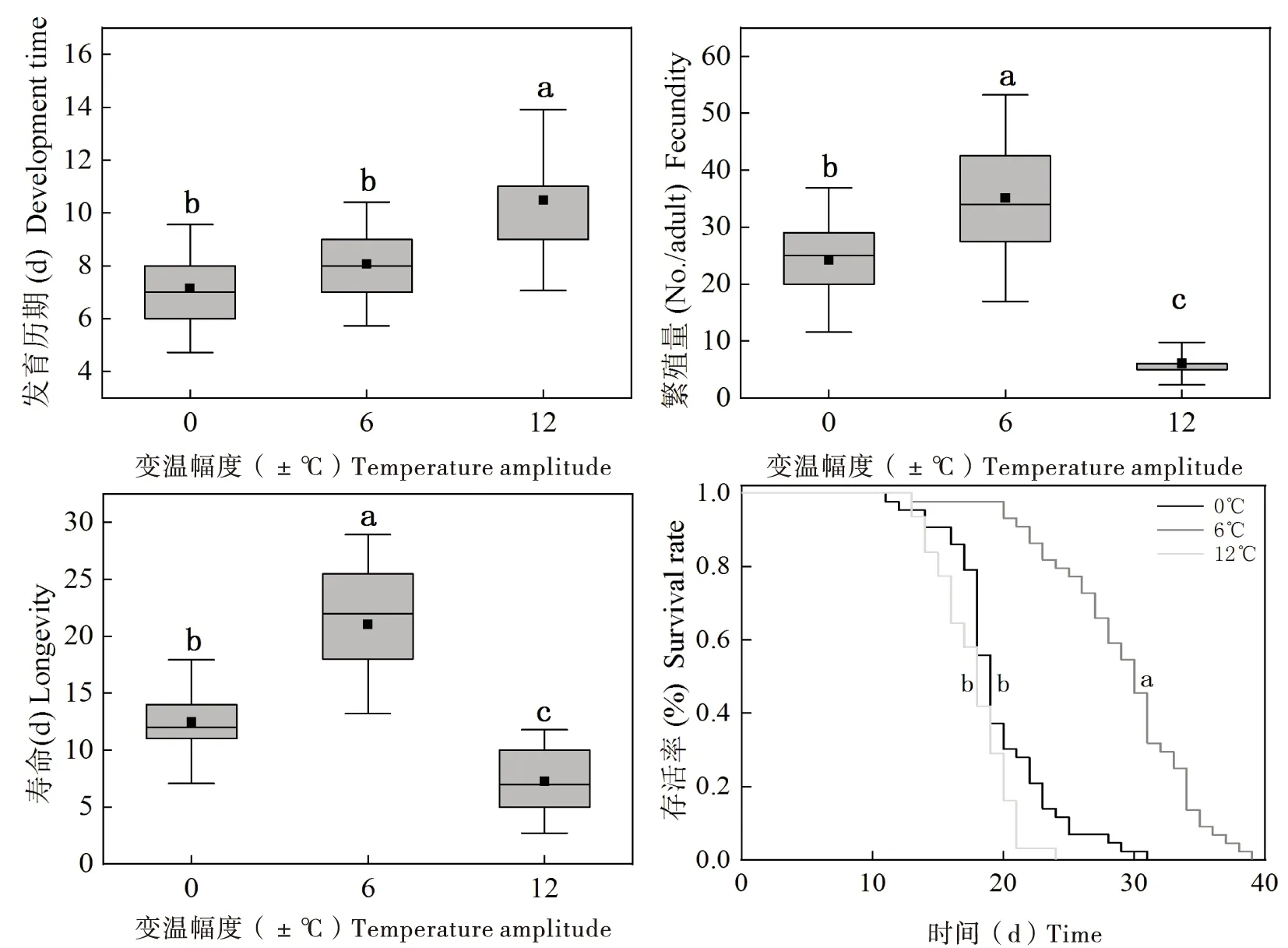
图4 不同变温幅度对麦长管蚜若虫发育历期、寿命、繁殖和存活情况的影响Fig.4 Effect of different temperature fluctuations on nymph development time, adult longevity, fecundity and survival rate of Sitobion avenae注:图中不同小写字母表示不同处理间差异显著(P<0.05)。Note: Different letters above the bar indicated significant differences (P<0.05).
不同变温幅度对麦长管蚜种群内禀增长率影响显著(χ2=263.11,df=2,P<0.01)(图5)。恒温(±0℃)与变温幅度(±6℃)下麦长管蚜种群内禀增长率分别为0.25±0.01、0.21±0.01,显著高于变温幅度(±12℃)下内禀增长率0.08±0.011。不同变温幅度对麦长管蚜种群净增值率影响显著(χ2=263.12,df=2,P<0.01)(图5)。变温幅度(±0℃、±6℃、±12℃)下麦长管蚜种群净增值率分别为21.74±1.58、31.90±2.42、3.21±0.48。不同变温幅度对麦长管蚜种群世代周期影响显著(χ2=263.12,df=2,P<0.01)(图5)。麦长管蚜种群世代周期由高到低分别是16.34±0.27(±6℃)、13.94±0.39(±12℃)、12.21±0.25(±0℃)。

图5 不同变温幅度对于麦长管蚜种群内禀增长率、净增值率和世代周期的影响Fig.5 Effect of different temperature fluctuations on nymph development time, adult fecundity and adult longevity of Sitobion avenae注:图中柱上不同小写字母表示不同处理间差异显著(P<0.05)。Note: Different letters above the bar indicated significant differences (P<0.05).
2.2 麦长管蚜表型之间相关性分析
对麦长管蚜生理生态表型之间进行相关性分析,以探索麦长管蚜表型之间可能的关联(见表1)。临界高温与呼吸速率(R2=0.653,P=0.401)、过冷却点(R2=0.358,P=0.592)、冰点(R2=0.940,P=0.160)、寿命(R2=0.507,P=0.661)、繁殖(R2=0.712,P=0.361)相关性不显著,过冷却点与呼吸速率(R2=0.508,P=0.855)、冰点(R2=0.004,P=0.960)、繁殖(R2=0.910,P=0.194)相关性不显著,冰点与呼吸速率(R2=0.418,P=0.553)、寿命(R2=0.102,P=0.793)、繁殖(R2=0.543,P=0.473)相关性不显著,呼吸速率与寿命(R2=0.561,P=0.461)、繁殖(R2=0.900,P=0.204)相关性不显著,寿命与繁殖(R2=0.941,P=0.156)相关性不显著。但是,过冷却点与寿命(R2=0.995,P=0.043)之间存在很强的相关性。

表1 麦长管蚜生理生态表型之间线性回归分析
3 结论与讨论
经历较大变温幅度后,麦长管蚜成蚜临界高温显著升高。这与以往短时热激与麦蚜耐热性研究结果相一致(Zhaoetal., 2017)。这可能是与较大变温幅度中短时高温诱导热激蛋白表达(蒋健等, 2013)以及山梨醇和甘露醇等热保护物质升高(Chownetal., 2010)有关。研究还发现经历较大变温幅度后成蚜过冷却点显著降低,并且成蚜临界高温与过冷却点的变化之间无显著线性关系。这说明,成蚜耐热性(临界高温)与耐寒性(过冷却点)的变化是相对独立(Bozinovicetal., 2016),这可能是由于成蚜温度耐受性对波动温度中热阶段与冷阶段的响应是不同的,可能存在不同的温度调节或响应机制(Colinetetal., 2015)。
本研究首次发现,在较大变温幅度下,麦长管蚜成虫呼吸速率显著提高。昆虫在适宜的温度环境范围内,呼吸速率处于平稳状态,仅有小范围变化;当不处于适温范围时,呼吸将放生较大变化(王冬梅等, 2014)。试验结果发现,较大变温幅度下,成蚜的呼吸速率明显提高。已有研究表明,昆虫呼吸速率随温度变化会出现升高或降低的不同状态(戈峰, 1991; 钱雪等, 2016)。这说明,昼夜变温幅度中短时日高温作为一种生理胁迫,已经对成蚜呼吸产生显著影响。并且,成蚜呼吸速率与变温幅度呈现正相关关系。由此可见,麦长管蚜在经历较大变温幅度时,可以通过提高自身呼吸代谢,改变对温度胁迫的适应能力。
昼夜变温幅度显著影响麦长管蚜若虫发育。当变温幅度为±0℃、±6℃时,麦长管蚜若虫发育之间无显著差异,但是当变温幅度为±12℃时,若虫发育历期显著延长。已有研究表明,发育和环境温度之间关系符合Jensen’s不等式中Sharpe-De Michele模型(Georgesetal., 2005)。在适宜温度附近且变温幅度较小时,昆虫发育与温度变化为线性模型(Escobaretal., 2012; Xingetal., 2014)。但是,当较大变温幅度中日高温接近或超过昆虫发育温度上限时,生物发育受到显著抑制(Martinetal., 2008)。试验中,变温幅度为±12℃时,日最高温度达到了34℃,且超过了麦长管蚜若虫发育上限温度(30℃)达到5 h。因此,较大变温幅度明显延长了麦长管蚜若虫的发育,这可能是由较大变温幅度中日高温胁迫造成若虫体内保幼激素水平变化造成的(Goodmanetal., 1990)。
与变温幅度±0℃、±6℃相比,较大变温幅度±12℃下麦长管蚜若虫存活增加,成虫寿命与生殖显著降低。这结果与以往恒温研究并不一致。恒温10~25℃范围内,麦长管蚜若虫存活变化较小(91.8%~97.0%),寿命在15.1~24.7 d之间;但温度升高至30℃时,麦长管蚜则无法存活与生殖(Dean, 1974)。试验中,在变温幅度±12℃下不仅麦长管蚜若虫可以发育,而且成虫还可以存活与生殖,这可能是因为,1)在日高温后,适宜恢复温度对麦长管蚜若虫与成虫均有热损伤的修复作用(Zhaoetal., 2014)。2)活动能力较强的成虫,由于频繁的移动,可以获得调节体温能力,从而躲避高温带来的损伤,提高自身耐热性(Maetal., 2012)。并且,还发现变温幅度±6℃时,麦长管蚜若虫存活、成虫寿命与生殖达到最大值,这可能由于变温幅度±6℃时,麦长管蚜若虫与成虫在日高温(28℃)并未对其产生致命损伤,但是经历适宜的夜间温度不仅修复了日高温造成的损伤,而且积累较多的保护性物质,进一步提高了若虫存活,也延长了成虫寿命与促进了生殖。
昼夜温差在一定程度上降低了昆虫种群的适合度,抑制种群增长。试验结果表明,变温幅度±12℃时麦长管蚜种群的内禀增长率明显低于变温幅度±0℃、±6℃。麦长管蚜种群增长最适宜温度在20~25℃之间,变温幅度±12℃时由于种群在白天日高温(34℃)显著抑制了生殖率,从而降低了种群增长。由此可见,当环境温度波动变化足够高时引起昆虫体内产生某种生理胁迫,破坏了体内生理平衡,降低了昆虫种群的适合度,从而使种群的增长受到一定的抑制。
随着变温幅度增加,麦长管蚜耐寒性(过冷却点)与寿命显著负相关(R2=0.995,P=0.043)。这说明,经历较大变温幅度的锻炼对麦长管蚜耐寒性提高是有益处的,但是带来的成本也是显著的,麦长管蚜寿命的降低。已有研究表明,多种生物经历环境温度波动锻炼(temperature acclimation)后,都会明显表现出一种成本与利益(cost and benefit)的权衡关系,如,长红锥蝽Rhodniusprolixus(Rolandi, 2018)、黑腹果蝇(Schouetal., 2015);苹果蠹蛾Cydiapomonella(Chidawanyika and Terblanche, 2011)。由此可见,经历较大温度波动锻炼后,虽然麦长管蚜通过某种表型的提高,增加其对环境变化的生理适应能力,但这种高能量的消耗会产生较大的成本,这种过程可能通过降低其它表型来实现。随着全球气候变化下温度变异性的增加,这种权衡现象将增加昆虫表型的变化情况。
以往多数种群表型及数量动态预测模型适用的温度均是恒温(Samanthaetal., 2014),那么昼夜温度波动中白天几个小时的高温对昆虫的作用往往容易被忽视。因而,昼夜变温幅度也必须纳入到种群表型及动态预测模型中,这样不仅能真正的全面阐明自然界复杂变温模式对麦长管蚜种群生理生态学的影响,而且能提高麦蚜田间发生预测预报的准确性以及提升气候变化对生物风险评估的精确性。
