外源ALA促进亚低温胁迫下番茄幼苗营养元素积累与分配
2022-06-02袁路乔贾媛婕张政达薛建康胡晓辉
康 珍,袁路乔,贾媛婕,张政达,薛建康,胡晓辉,3*
(1.西北农林科技大学园艺学院,陕西 杨凌 712100;2.农业农村部西北设施园艺工程重点实验室,陕西 杨凌 712100;3.陕西省设施农业工程技术研究中心,陕西 杨凌 712100)
番茄(Solanum lycopersicum)是我国主要设施蔬菜作物之一[1]。我国现有设施环境调控能力弱,北方冬春季设施生产中的番茄常遭遇亚低温胁迫[2]。亚低温影响番茄植株叶绿素合成、营养吸收与分配和生长发育,导致植株营养不良等生理障碍[3-4]。低温对植株生长发育的阻碍程度与叶片内矿质元素含量减少的程度呈显著正相关[5]。
植物需矿质元素维持自身正常生理生化活动[4],通过生理或生化代谢作用于植株生长,其中氮、磷、钾、钙和镁元素既可作为植物体内重要化合物组分构成植物体结构,又可作为酶激活剂和渗透调节剂参与能量代谢过程,保证植株生理生化活动[6-7],氮、镁和铁元素可参与植物叶绿素合成、光合和呼吸作用等,在植物生命活动中起重要作用[8]。在低温和盐碱等逆境下植物细胞常常积累一些无机离子以调节渗透势,增强自身抗逆性[9-10],植株各器官中矿质元素含量对番茄设施栽培具有重要意义[11]。5-氨基乙酰丙酸(5-aminolevu⁃linic acid,ALA)作为一种非蛋白类氨基酸,广泛存在于动植物、真菌和细菌中[12]。叶面喷施ALA可增强亚低温下细胞清除活性氧能力,提高渗透调节能力,保护玉米幼苗叶片细胞膜结构和功能[13],促进恒定亚低温下黄瓜幼苗根系和叶片中养分积累[14],调节番茄幼苗内源激素水平和叶绿素合成[12],增强植物低温抗性。
关于外源ALA 增强番茄亚低温耐性的研究主要围绕植株光合作用、氧化胁迫及内源激素等方面,而关于非恒定亚低温状态下,植物不同营养元素积累与分配的研究较少。本研究以养分利用为切入点,研究外施ALA 对番茄幼苗主要矿质元素累积和分配的影响,以期明确ALA 对亚低温下番茄元素代谢的影响,为调节亚低温胁迫下番茄养分平衡及优质高产提供合理有效技术参考和理论依据。
1 材料与方法
1.1 试验材料
供试番茄品种为‘金棚一号’(低温敏感型[3])。选用商品性基质(购于山东省寿光市天丰园艺材料厂)栽培,其理化性质为硝态氮6179 mg·kg-1,铵态氮535 mg·kg-1,速效磷68 mg·kg-1,速效钾631.23 mg·kg-1,pH 4.88,电导率(EC)值1 012 μS·cm-1。
1.2 试验设计
本试验于2020年2~10月在陕西省杨凌区西北农林科技大学设施农业生物与环境工程实验室进行(34°28′N、108°07′E)。选择完整饱满、均匀一致番茄种子经消毒、浸种、催芽、露白后,采用72孔穴盘育苗,育苗环境为温度(25.0±4.0)/(20.0±2.0)℃(昼/夜)、光照280~400/0 μmol·m-2·s-1(昼/夜)、空气相对湿度(RH)60%±10%/70%±10%(昼/夜)。子叶展平后适时、适量浇灌1/4 倍Hoagland营养液。选取健壮、长势一致4叶1心幼苗定植于营养钵中(7 cm×7 cm,每钵1 株),并置于智能人工气候箱(DWRG-1000D4,合肥右科仪器设备有限公司)中培养,培养环境条件:温度,16~28 ℃(昼/夜);RH,65%±5%;光照,12 h [光量子通量密度(PPFD),350 μmol·m-2·s-1]/12 h(PPFD,0 μmol·m-2·s)-1(昼/夜)。待番茄长至5叶1心时按照50 mL·plant-1浇1 次1/2 剂量Hoagland 营养液,缓苗3 d后开始试验处理。模拟昼夜温度变化规律设置常温(16~28 ℃/16~18 ℃,昼/夜)和亚低温(8~18 ℃/8~12 ℃,昼/夜)范围,光周期为12 h/12 h。除温度外其他环境条件均与幼苗培养条件相同。叶面预喷施6 mL·plant-1蒸馏水(0 mg·L-1ALA)或6 mL·plant-125 mg·L-1[3]ALA 溶液,常温培养12 h后,开始不同常温或亚低温处理,分别为清水+常温(CK)、ALA+常温(ALA)、清水+亚低温(LT)和ALA+亚低温(LTA),每个处理15 株,并重复3次。为保证基质营养充足,于处理第6天时所有处理浇灌等量营养液。亚低温处理第3、6、9、12和15天时测定番茄第3片真叶叶绿素含量,处理15 d时收集植株根、茎和叶,测定生物质量和各元素含量。
1.3 测定项目及方法
1.3.1 番茄幼苗生物质量测定
清洗干净植株根、茎、上位叶(第4 片真叶及以上叶片)和下位叶(第5 片真叶及以下叶片)四部分分别称量鲜重后于105 ℃杀青15 min,再75 ℃烘至恒重,称量干重。
1.3.2 番茄幼苗叶片叶绿素含量测定
叶绿素含量采用乙醇浸提法测定[15]。
1.3.3 植株元素含量测定
将样品用微型粉碎机粉碎后过筛(0.5 mm)。样品经浓HNO3-H2O2消煮后用原子吸收分光光度计测定(Z-5000型,美国珀金埃尔默企业管理有限公司)钙含量,采用原子吸收光谱法测定镁、铁元素含量。样品浓H2SO4-H2O2消煮后采用高分辨自动化学分析仪(AA3 型,德国SEAL)测定全氮、全磷和全钾含量。
1.3.4 相关指标计算
依据张忠学等[16]和杜少平等[17]试验方法,适当调整得出以下指标计算公式:
养分积累量(mg·plant-1)=干物质量×养分含量;
干物质分配率(%)=各器官干物质量/植株总干物质量×100;
各器官养分分配率(%)=各器官养分积累量/植株总养分积累量×100。
1.4 数据处理与分析
采用Microsoft Excel 2007记录、整理数据并制作图表,SPSS 25.0 作单因素方差分析,Ducan法作显著性检验(P<0.05)。图表中数据为平均值±标准差。
2 结果与分析
2.1 外施ALA对亚低温胁迫下番茄幼苗鲜重和干重的影响
常温下喷施ALA 可促进番茄幼苗根、茎生长,但对叶片生长无促进效果。与CK相比,单独亚低温胁迫(LT)15 d 番茄幼苗茎、上位叶、下位叶、叶总鲜重和植株总鲜重分别显著降低35.3%、34.2%、41.1%、34.2%和31.2%,相应器官干重和植株总干重分别显著降低23.3%、19.1%、30.8%、26.7%和23.5%。与LT 处理相比,低温下预喷施ALA(LTA)处理显著增加番茄幼苗相应器官及植株总鲜重和干重,分别为17.3%、19.0%、26.8%、23.3%、19.0%和11.4%、19.1%、20.6%、20.0%、17.0%。表明亚低温胁迫显著抑制番茄幼苗各器官生物量增加,外施ALA 显著缓解亚低温胁迫对番茄幼苗生长的抑制(见图1)。
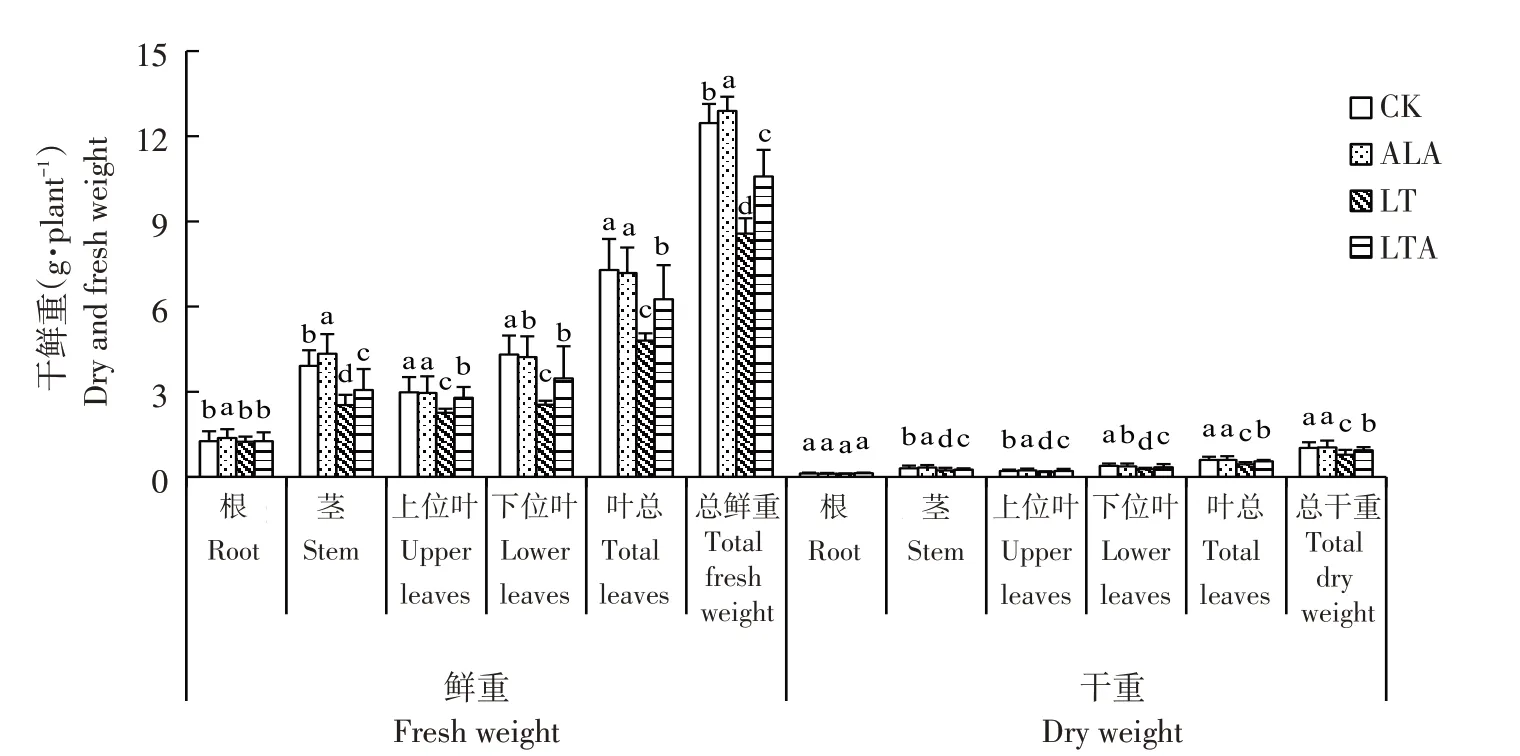
图1 外施ALA对亚低温胁迫下番茄幼苗不同器官鲜重和干重的影响Fig.1 Effects of exogenous ALA on the fresh and dry weight of tomato seedling organs under sub-low temperature stress
2.2 外施ALA对亚低温胁迫下番茄幼苗叶片叶绿素含量的影响
常温下喷施ALA 短时间内(3 d)促进番茄幼苗叶绿素含量增加,但随植株生长促进效果消失。亚低温显著降低番茄幼苗叶片叶绿素含量,胁迫6 d时表现显著抑制现象,随胁迫时间增加抑制作用愈加明显,胁迫15 d 时抑制作用最为显著,此时LT 处理番茄幼苗叶片叶绿素含量较CK 显著降低32.2%,而LTA 处理叶绿素含量与LT 处理相比显著增加21.5%。胁迫期间,外源ALA对因亚低温造成的叶绿素含量下降均呈缓解作用,在胁迫后期尤其显著(见图2)。

图2 外施ALA对亚低温胁迫下番茄幼苗叶片叶绿素含量的影响Fig.2 Effects of exogenous ALA on chlorophyll content of tomato seedling leaves under sub-low temperature stress
2.3 外施ALA对亚低温胁迫下番茄幼苗各器官氮磷钾元素积累量及分配率的影响
常温下喷施ALA 显著降低番茄幼苗根系中钾素和茎中氮、磷素分配率及根系中氮、磷、钾元素,茎中氮、磷元素积累量,增加上位叶和叶片总氮、总磷素分配率;对番茄幼苗叶片和全株氮、磷、钾元素积累均无促进作用(见表1)。
与CK 相比,LT 处理番茄幼苗上位叶、下位叶、叶片总氮素和全株氮素积累量分别显著降低22.9%、43.2%、35.3%和28.6%,根系氮素分配率增加58.7%,下位叶和叶片总氮分配率分别显著降低20.0%和8.8%。与LT处理相比,LTA处理番茄幼苗相应器官中氮素积累量显著增加89.2%、57.1%、72.0%和55.2%,均与CK相近,而番茄幼苗根系和茎中氮素积累量均无显著差异;LTA 与LT 处理幼苗各器官中氮素分配率变化规律相反,表现为显著降低番茄根系氮素分配率,显著增加叶片总氮分配率。亚低温胁迫下外施ALA 通过调节氮素在番茄幼苗不同器官中的积累与分配显著缓解因低温造成的番茄幼苗叶片中氮素积累量下降(见表1)。
与CK相比,LT处理番茄幼苗根系、茎、下位叶、叶片总磷素积累量和全株磷素积累量分别显著减少24.6%、48.5%、46.7%、25.2%和31.7%,同时茎和下位叶磷素分配率分别显著降低25.5%和21.4%,上位叶和叶总磷素分配率分别显著增加63.3%和10.0%;而LTA 处理显著缓解亚低温对番茄幼苗磷素积累的负面影响,表现为茎、下位叶、叶片总磷素积累量和全株磷素积累量分别比LT 处理显著增加94.3%、82.3%、60.8%和61.4%。外施ALA 仅降低根系磷素分配,对其他器官磷素分配无显著影响(见表1)。
与CK 相比,LT 处理番茄幼苗茎中钾素积累量与分配率分别显著降低38.1%、19.3%,上位叶和叶片总钾素分配率分别显著增加38.4%、12.2%,而LTA处理番茄幼苗茎中钾素积累量比LT处理显著增加72.6%,与CK 接近,各器官钾素分配率与LT处理均无显著差异(见表1) 。
2.4 外施ALA对亚低温胁迫下番茄幼苗各器官钙镁铁元素积累量及其分配率的影响
常温下喷施ALA 显著增加番茄幼苗根系钙素、茎中钙镁素和叶片总铁素分配,降低根系铁素、上位叶钙素和叶片总铁素分配;显著增加根系中钙素、茎中钙和镁素及全株总钙素积累量,降低根系中铁素和上位叶钙素积累量,对番茄幼苗下位叶和叶片总钙、镁、铁素积累及上位叶和全株镁、铁素积累均无促进作用(见表2)。

?

?
与CK 相比,LT 处理番茄幼苗上位叶、下位叶、叶片总钙素和全株钙素积累量分别显著降低77.4%、45.1%、54.7%和52.7%,同时上位叶及总叶片钙素分配率分别显著降低51.9%和3.8%,茎及下叶位钙素分配率显著增加137.7%、16.8%;而LTA处理显著缓解低温对番茄幼苗钙素积累的负面影响,表现为LTA 处理番茄幼苗相应器官中钙素积累量与LT 处理相比分别显著增加90.5%、91.3%、91.2%和93.9%,均与CK 接近。外施ALA仅增加茎中钙素分配,对其他器官钙素分配无影响(见表2)。
与CK相比,LT处理番茄幼苗下位叶、叶片总镁素和全株积累量分别显著减少51.4%、36.1%和32.5%,同时下位叶镁素分配率显著降低27.40%、根系和上位叶镁素分配率分别显著增加48.0%和48.7%;而LTA处理显著缓解低温对番茄幼苗钙素积累的负面影响,表现为LTA 处理相应器官中镁素积累量与LT 处理相比分别显著增加77.6%、53.5%和56.7%,同时根系中镁素分配率显著降低26.1%,均与CK 接近。外施ALA 除增加茎中镁素分配外对其他器官镁素分配均无显著影响(见表2)。
与CK相比,LT处理番茄幼苗根系、上位叶和全株铁素积累量分别显著下降57.1%、42.9%和27.6%,下位叶铁素积累量显著增加60.0%,同时根系和上位叶铁素分配率分别显著降低33.4%和29.6%,下位叶和叶总铁素分配率分别显著增加112.5%和29.9%;LTA处理显著缓解低温对番茄幼苗钙素积累的负面影响,表现为LTA 处理番茄幼苗上位叶和全株铁素积累量与LT 处理相比分别显著增加150.0%和57.1%,均达到CK 水平,同时根系和下位叶铁素分配率分别显著降低34.2%和44.2%,上位叶铁素分配率显著增加64.1%,在LT与LTA 两处理间番茄幼苗叶片总铁素分配率无差异情况下,ALA 促进铁素由下叶位向上叶位转运,增加铁素在上位叶的分配与积累。
3 讨论
外源施用ALA 可通过提高植物抗氧化和光合能力减轻亚低温胁迫伤害[18-19]。本研究发现,亚低温胁迫显著降低番茄幼苗地上部组织鲜重、干重和叶片叶绿素含量,抑制番茄幼苗生长;而外施ALA 促进低温胁迫下幼苗形态建成和叶绿素合成(见图1 和2),缓解亚低温胁迫对番茄幼苗生长的限制,与其他物种研究结果类似[20-22]。矿质元素在植物体各器官中分配比例反映该元素在植株体内分布及其在各器官间迁移规律[23],其受温度影响[24-25]。本试验中,亚低温胁迫显著增加氮镁元素在根系、磷钾镁元素在上位叶、钙铁元素在下位叶和磷钾铁元素在总叶片中分配率,而各器官中相应元素积累量均未增加且全株各元素积累总量均降低(见表1 和2)。亚低温胁迫造成根系皮层厚度增加进而增大离子运输阻力[11],同时植株根系中相关离子转运蛋白基因表达下调、根系对离子的吸收量降低[24],导致氮素由根系向叶片、钙素由茎及老叶向新叶、铁素由老叶向新叶和镁素由根系向叶片的转运受阻。
亚低温可能通过抑制叶片氮素同化过程中关键酶活性和磷酸盐等转运蛋白基因表达导致氮素同化量减少,显著降低番茄幼苗叶片氮磷积累量(见表1),与前人研究结果一致[11,24]。上位叶中氮、磷素对亚低温更为敏感,亚低温胁迫下植株自身率先保证新叶氮、磷素尤其是磷素积累,通过茎和上位叶中磷素向新叶转运从而维持新叶磷素积累,促进植株在低温下能量代谢,为生殖生长存储和传递遗传信息[25]。与氮、磷素不同,亚低温胁迫增加番茄幼苗下位叶铁素积累量与分配率,降低上位叶铁素积累量与分配率,说明亚低温抑制铁素由老叶向新叶转运,这是因为铁元素在植物体内移动性较差,当亚低温下铁素吸收受抑时,植株整体处于缺铁状态,而铁又难以从老叶中转移出来进入幼叶供幼叶生长,致使新叶中铁素积累量显著降低。由于低温刺激诱发钙信号跨膜传递,随处理时间增加茎中钙积累量呈逐渐上升趋势,减少向叶片中运输量,致使叶中钙素积累量减少,故亚低温胁迫下番茄幼苗上位叶钙素、下位叶钙镁素积累量显著降低,与张佩茹根区低温对番茄幼苗矿质元素吸收运输影响研究结果一致[11]。低温也影响镁转运蛋白酶活性[11],导致根系对镁素吸收量减少,同时茎对镁素运输量减少,造成叶片中镁素积累量降低。研究表明,在植株遭遇非生物胁迫初始阶段,植株为抵抗逆境大量吸收K+[26],而本试验中15 d亚低温胁迫可能使植株产生抗性惰性,根系、叶片及单株钾素积累量不再显著增加(见表1),抗逆能力下降。与单纯亚低温胁迫处理相比,外施ALA 显著促进番茄幼苗上位叶、下位叶和单株氮素积累;亚低温条件下番茄植株对氮素转运能力降低,氮在根系中积滞,而ALA 通过缓解亚低温下氮素在根系中的积滞,促进氮素由根系向叶分配,缓解低温伤害(见表1),ALA 促进氮素吸收并由根向叶转运的效果在黄杨中也得到证实[26],可能是因为ALA调节亚低温胁迫下植株新、老叶氮代谢中硝酸还原酶和谷氨酰胺合成酶等相关酶活性[27],促进番茄上、下位叶片中氮素积累。相比ALA 对氮素转运作用,本试验发现亚低温下番茄幼苗各器官中磷钾素分配情况未受ALA影响(见表1),表明外施ALA主要是通过促进亚低温下番茄幼苗对磷、钾素吸收缓解低温伤害,其原因可能是外施ALA 增加低温下番茄幼苗根系活力、上调根系中相关元素通道基因和转运蛋白基因表达[28],促进亚低温下植株体内磷钾素积累。氮、磷、钾元素在植物生长发育、光合作用和物质代谢等方面均发挥极其重要作用[11],外施ALA 通过缓解亚低温引起的番茄幼苗氮、磷、钾元素吸收障碍,促进亚低温下植株中各元素积累,促进亚低温下番茄幼苗生长。
与磷钾素类似,外施ALA 显著缓解亚低温对叶片钙、镁素积累的抑制作用,使番茄幼苗叶片及植株总钙、镁积累量维持在正常水平,但ALA对叶片中钙、镁分配率无显著影响(见表2),说明ALA 主要通过促进钙、镁元素吸收增加叶片中钙、镁积累。钙作为第二信使可偶联胞外信号与胞内生理生化反应,亚低温下ALA 可能诱发钙信号跨膜传递从而促进钙素积累,调控细胞生长、促进亚低温下番茄幼苗叶片生长;镁主要依赖蛋白酶以主动运输方式进入根系[7,11],结合各器官中镁素分配情况可知,亚低温下番茄幼苗下位叶及植株总镁素含量降低可能是长时间亚低温影响镁转运蛋白酶活性,根系对镁素总吸收量减少所致,ALA 可能通过调节镁转运蛋白酶活性促进镁素吸收利用。本试验结果表明,亚低温抑制铁素由老叶向新叶转运,亚低温下ALA 预处理的番茄幼苗中铁素在下位叶的分配率与单纯亚低温相比显著下降且铁素积累量无显著变化,上位叶铁素积累量与分配率显著增加,均保持在正常水平,同时叶片和全株总铁素积累量显著增加,说明ALA通过促进亚低温下植株对铁素的吸收并促进铁素由老叶向新叶转运实现番茄幼苗上位叶铁素的积累,缓解叶片失绿进而缓解低温伤害。但当前针对该生理效应的内在机理研究尚不深入,关于外施ALA 如何作用于铁素在叶片中转运的机制有待进一步探究。除铁元素外,氮镁元素减少也导致叶绿素含量下降[11,17],影响植物光合效率[25],而外施ALA 增加亚低温下叶片中氮、镁、铁元素积累量并缓解氮、铁元素转运障碍,缓解叶片叶绿素含量降低,提高植株光合作用,促进番茄幼苗生长。本研究通过模拟自然亚低温动态变化,发现外施ALA 可缓解亚低温对番茄幼苗元素利用的抑制作用,与前人在其他作物上研究类似[13-14]。但关于ALA 在亚低温下对作物营养元素吸收和转运发挥作用的具体机理仍需深入探究。
4 结 论
亚低温胁迫限制番茄幼苗对氮、磷、钾、钙、镁和铁元素的吸收,抑制磷钾铁元素在根系、磷钾元素在茎和氮磷钙镁铁元素在叶片的积累,抑制氮素由根系向叶、铁素由下位叶向上位叶转运,并造成叶片叶绿素含量下降。亚低温胁迫下,外施25 mg·L-1ALA 促进番茄幼苗对氮、磷、钾、钙、镁和铁元素的吸收及积累并促进氮、铁元素转运,缓解亚低温胁迫对番茄幼苗各器官中营养元素积累与分配的不利影响和叶绿素含量降低,促进番茄幼苗生长。
