树鼩永生化皮肤成纤维细胞株的建立及鉴定*
2022-05-28奎秀莹王文广金亮子仝品芬陆彩霞代解杰
奎秀莹,王文广,金亮子,邓 伟,仝品芬,陆彩霞 ,代解杰
(1.中国医学科学院,北京协和医学院 医学生物学研究所,云南 昆明 650031;2.昆明医科大学 医学生物学研究所,云南 昆明 650031;3.云南大学 医学生物学研究所,云南 昆明 650031)
皮肤成纤维细胞是网状真皮层中较为常见的细胞[1],能够在结缔组织中分泌和沉积大量整合素和细胞外基质(extracellular matrix,ECM),包括胶原蛋白、弹性纤维和结合蛋白等[2]。由于能够分泌一系列的ECM,皮肤成纤维细胞在皮肤的衰老、光老化[3-4]、炎症、烧伤或机械损伤[5]、瘢痕形成[6]和肿瘤及其他皮肤相关疾病等多种皮肤修复过程中起至关重要的作用。目前,已有研究者成功分离出小鼠皮肤成纤维细胞[7]、大鼠皮肤成纤维细胞[8]、人皮肤成纤维细胞[9]、裸鼹鼠皮肤成纤维细胞[10]和驴皮成纤维细胞[11],并基于不同物种的皮肤成纤维细胞开展了皮肤疾病相关研究。树鼩属攀鼩目小型哺乳动物,具有体型小、繁殖快的特点,进化上比啮齿类更接近非人灵长类。FAN 等[12]研究显示:树鼩在神经系统、视觉系统和免疫系统上与人相似,已被用于眼部[13]、视神经和脑部神经发育相关疾病以及多种肝炎病毒[14-17]、寨卡病毒[18]、单纯疱疹病毒[19]和流感病毒[20]等疾病的模型研究中。已有研究对树鼩进行肺纤维化诱导,建立了与临床患者出现相似症状的肺纤维化模型[21]。ZHANG 等[22]对树鼩皮肤结构进行解析,发现其与恒河猴及人的皮肤结构极为相似,有丰富的汗腺和皮脂腺,且汗腺分布在毛球顶部周围,与毛囊皮脂腺等共同组成与人类相似的附属皮肤单位,而小鼠汗腺和皮脂腺并不明显,表明树鼩是皮肤相关疾病研究的潜在非人灵长类替代实验动物。
课题组前期分离出原代树鼩皮肤成纤维细胞[23],然而,原代细胞往往只能够稳定增长几个代次,传代次数过多细胞便会出现各种问题,如细胞形态发生改变和细胞增殖变缓,试验过程中很容易错过细胞的最佳生长时期,阻碍了皮肤相关疾病细胞模型的构建和相关机制的研究。此外,对于需要反复验证的试验,原代细胞几乎无法再次重复,每一批次分离出的细胞难免会有差异,并且耗时长,试验效率会受到很大影响。因此,建立能够连续多次传代的永生化细胞可弥补原代细胞的不足。本研究通过优化分离树鼩原代皮肤成纤维细胞的方法,再利用携带SV40T的慢病毒转染细胞,经过50 代以上的传代,以期获得永生化树鼩皮肤成纤维细胞,为皮肤相关疾病的发病机制研究提供新材料。
1 材料与方法
1.1 试验动物和慢病毒
供试树鼩为滇西亚种中缅树鼩(Tupaia belangeri chinensis),由中国医学科学院医学生物学研究所树鼩种质资源中心提供,雄性,1 岁,生产许可证号为SCXK(滇)K2018-0002,使用许可证号为SYXK(滇)K2018-0002。本研究遵循试验动物使用3R 原则和福利伦理原则。
携带SV40T基因的慢病毒由汉恒生物科技(上海)有限公司构建,病毒滴度为1×109TU/mL。
1.2 树鼩成纤维细胞的酶消化法分离纯化
本试验在白剑等[23]的酶消化法基础上稍作改进。取约1 岁的树鼩腹腔注射3%戊巴比妥钠0.5 mL 麻醉后处死,用75%酒精进行皮肤消毒,刮除树鼩大腿内侧的毛发,剪下约2.5 cm×1.5 cm 的皮肤组织块,在含2%青霉素—链霉素(美国Gibco)的PBS(美国 Hyclone)中反复刮洗3次,将组织块剪碎为约1 mm3的块状,置于50 mL离心管中,加入0.1% Ⅰ型胶原酶(PBS 配制),于37 ℃恒温箱中消化45 min,每15 min 振荡1 次,使组织块与Ⅰ型胶原酶充分接触。消化完成后用100 目的钢网筛过滤细胞悬液,1 000 r/min离心10 min,弃上清后加入完全培养基[ 配方为:MEM 基础培养基(以色列Biological Industries)+10% FBS(以色列Biological Industries)+1%青霉素—链霉素+1% NaHCO3+1%非必需氨基酸(美国Gibco)]重悬细胞,1 000 r/min 离心10 min,用完全培养基再次重悬后转至6 孔板中,置于37 ℃、5% CO2培养箱中培养,每天观察细胞生长状况。待细胞铺满皿底时,用PBS 清洗原代细胞3 次,每孔加入0.25% Trypsin-EDTA 胰蛋白酶(美国Gibco)500 μL,室温消化3 min 后吸弃胰蛋白酶,加入完全培养基反复吹打,待变圆的皮肤成纤维细胞脱落后,将细胞悬液转至培养瓶中培养。
1.3 慢病毒转染树鼩成纤维细胞
将生长状态良好的皮肤成纤维细胞以1×105mL-1的密度接种于6 孔板,每孔2 mL,放入37 ℃、5% CO2培养箱中培养;待细胞汇合度达到80%以上时,用PBS 清洗2 次,加入含4 μg/mL助转剂的培养液1 mL,以感染复数(MOI)为30 接种携带SV40T基因的慢病毒,并设不接种的空白对照孔,置于37 ℃、5% CO2培养箱中培养4 h 后补加含4 μg/mL 助转剂的培养液1 mL,培养24 h 后更换为新鲜且不含助转剂的完全培养液继续培养24 h。之后,将培养液更换为含4 μg/mL嘌呤霉素的培养液继续培养,2 d 更换1 次培养液,直至空白对照孔的细胞完全死亡。
1.4 有限稀释法挑取单克隆
细胞汇合度达到80%以上时,用0.25%Trypsin-EDTA 胰蛋白酶消化并计数,将细胞稀释至密度为103mL-1,再进行10 倍梯度稀释,直至约10 mL-1,每个密度梯度接种16 孔(2 排)96 孔板,每孔100 μL,置于37 ℃、5% CO2培养箱中培养,每隔1 d 观察并标记由单个细胞增殖形成的单个细胞群。待细胞长满后,消化转至24 孔板中继续培养,并逐步扩大培养,每消化1 次记为1 代。
1.5 树鼩成纤维细胞传代
细胞汇合度达80%以上时,弃去培养液,PBS 清洗2 次,用0.25%胰蛋白酶消化,1.5 min后吸弃胰蛋白酶,待大部分细胞变圆、培养瓶底部变成毛玻璃状时加入培养液终止消化,以1∶6的比例传代培养,定期拍照保存。
1.6 树鼩永生化皮肤成纤维细胞生长曲线的测定
取第52 代细胞用胰蛋白酶消化后,稀释至密度为5×104mL-1,接种于96 孔板,每孔100 μL,按照CCK-8 试剂盒(上海东仁)说明书测定细胞生长曲线,检测6 d 的细胞增殖情况,并以6 d的吸光度值绘制细胞增殖动力曲线图。
1.7 免疫荧光鉴定
取第52 代细胞,经消化后将其稀释至密度为1×105mL-1,接种于24 孔板,每孔500 μL,同时用树鼩原代脑内皮细胞作对照。待细胞贴壁后,PBS 清洗2 次;用4 ℃预冷的4%多聚甲醛于室温固定细胞20 min,PBS 清洗3 次,每次5 min;0.5% Triton X-100(PBS 稀释)室温通透20 min,PBS 清洗3 次,每次5 min;用0.01 mmol/L PBS 稀释至20%的山羊血清封闭液封闭30 min,PBS 清洗3 次,每次5 min;加入1∶500 的鼠抗波形蛋白单克隆抗体和1∶50 的兔抗SV40T 单克隆抗体(一抗稀释液稀释,每个抗体2 个复孔),4 ℃孵育过夜,PBS 清洗3 次,每次5 min;分别加入1∶500 的羊抗鼠二抗和驴抗兔二抗(用0.01 mmol/L PBS 稀释),37 ℃避光孵育1 h,PBS 清洗3 次,每次5 min;加入5 μg/mL DAPI染色液,室温避光孵育5 min,对标本进行染核,PBS 漂洗4 次,每次5 min,在荧光显微镜下观察。
1.8 细胞核型分析
取第53 代细胞消化转至新的培养瓶中,置于37 ℃、5% CO2培养箱中培养24 h;加入4 ℃预冷的80 μg/mL 秋水仙素(美国Sigma)200 μL继续培养4 h,吸弃上清液,PBS 清洗3 次,每次5 min;用0.25%胰蛋白酶消化细胞并转至15 mL离心管中,1 000 r/min 离心5 min,弃上清;加入37 ℃预热的0.075 mol/L KCl 溶液7 mL,轻轻吹散细胞,室温低渗25 min,期间若有沉淀时需要轻轻吹打,使细胞与KCl 低渗液充分接触,1 000 r/min 离心5 min,弃上清;加入现配卡诺固定液(V甲醇∶V冰醋酸=3∶1)7 mL,固定5 min后沿管壁轻轻吹打,使沉淀的细胞翻转,固定5 min 后,轻轻吹散细胞,继续固定20 min,1 000 r/min 离心5 min,弃上清;再次以7 mL 固定液以同样方式固定25 min,1 000 r/min 离心5 min,弃去上清后重新加入适量固定液,重悬细胞。取4 ℃预冷的载玻片,吸管吸取混匀的细胞液悬空30 cm 垂直滴3 滴至载玻片上,斜靠载玻片流掉多余细胞液,自然风干12 h;将载玻片放入染缸中,加入Giemsa 染液(北京索莱宝)染色2 h,之后流水冲洗载玻片,自然风干12 h 后封片,在显微镜下用油镜观察。
2 结果与分析
2.1 不同代次的树鼩皮肤成纤维细胞
由图1 可知:树鼩皮肤成纤维细胞形态总体较均一,为梭形,较胖,细胞增殖速度较快。以1∶6 传代后,2 d 便长满培养瓶,可继续传代;培养2 d 后细胞上清中出现少许死细胞漂浮,45代后细胞增殖有所减慢,约3 d 可长满培养瓶,细胞状态也有所变化,部分细胞出现拉丝,细胞两端拉长,但大部分细胞仍能维持原有形态。

图1 不同代次的树鼩皮肤成纤维细胞(200×)Fig.1 Skin fibroblasts of Chinese tree shrew in different generations
2.2 生长曲线测定
由图2 可知:细胞数量在第2~3 天时明显增加,处于对数生长期,之后缓慢增长,在第5 天时达到高峰,进入平台期。
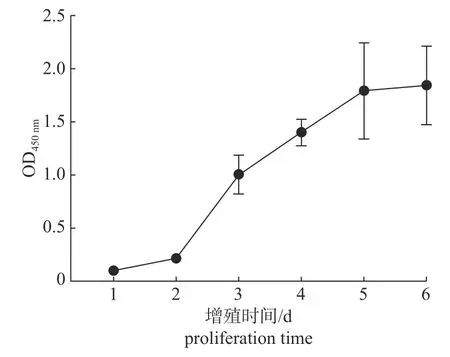
图2 第53 代树鼩皮肤成纤维细胞生长曲线Fig.2 The growth curve of Chinese tree shrew skin fibroblasts in the 53th generation
2.3 免疫荧光鉴定
由图3 可知:树鼩皮肤成纤维细胞细胞质中有明显的成纤维细胞特异性蛋白Vimentin 信号,而原代脑内皮细胞无该信号,结合树鼩原代皮肤成纤维细胞的分离方法及细胞形态学分析[23],判定分离得到的细胞是树鼩皮肤成纤维细胞。
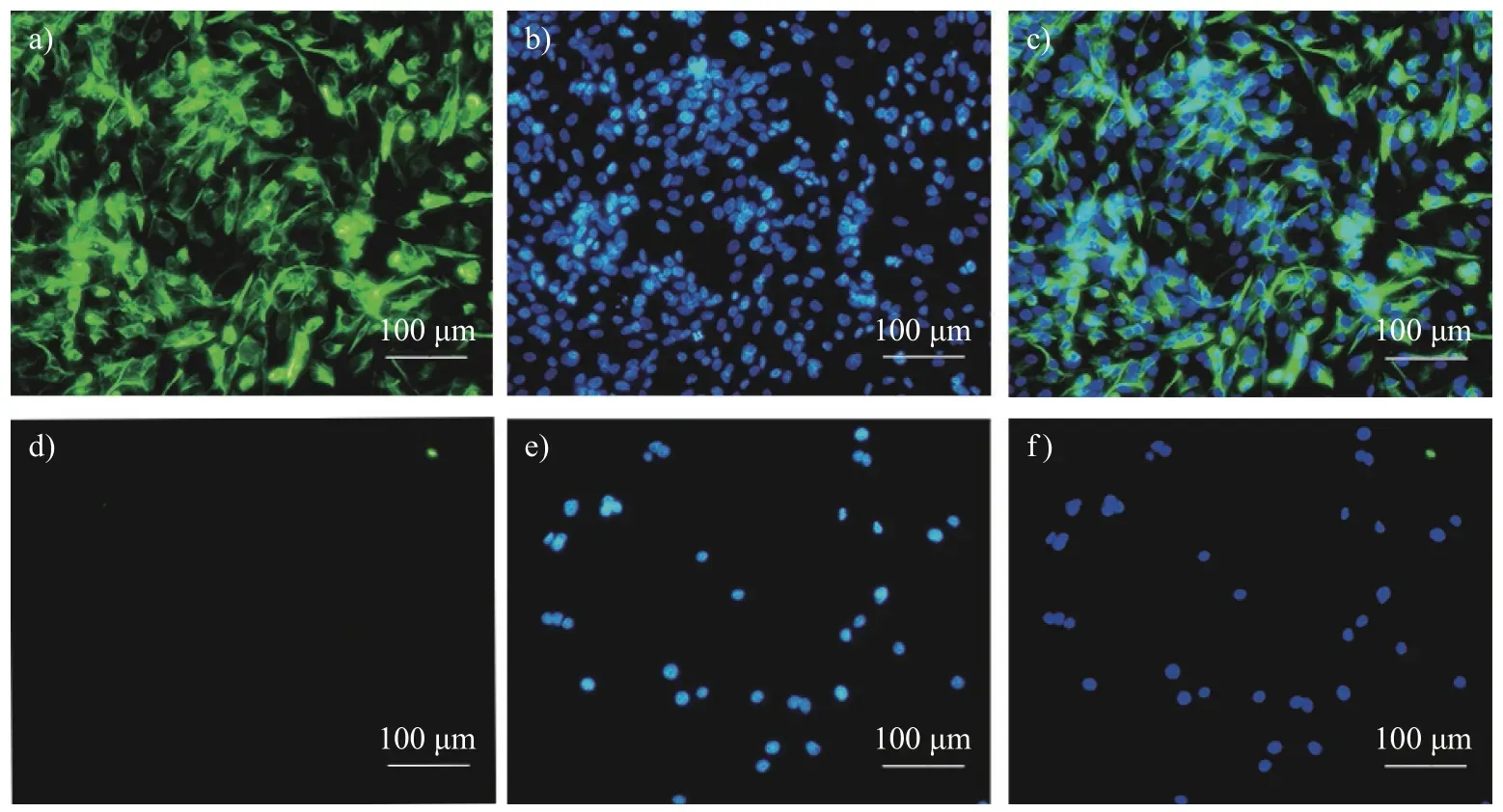
图3 第52 代树鼩皮肤成纤维细胞波形蛋白免疫荧光鉴定(200×)Fig.3 Vimentin identification of Chinese tree shrew skin fibroblasts in the 52th generation by immunofluorescence
由图4 可知:树鼩皮肤成纤维细胞细胞核处有明显的SV40T 信号,而原代脑内皮细胞无该信号,说明SV40T 已成功导入到所获得树鼩皮肤成纤维细胞中。

图4 第52 代树鼩皮肤成纤维细胞永生化基因SV40T 免疫荧光鉴定(200×)Fig.4 SV40T identification of Chinese tree shrew skin fibroblasts in the 52th generation by immunofluorescence
2.4 核型分析
选取53 代树鼩皮肤成纤维细胞进行核型分析,对细胞分裂相进行观察,结果(图5)显示:所获得的树鼩永生化皮肤成纤维细胞染色体数为62 条,与滇西亚种中缅树鼩的染色体数[24]一致,说明试验所得细胞是树鼩来源细胞。

图5 第52 代树鼩皮肤成纤维细胞核型分析Fig.5 Karyotype identification of Chinese tree shrew skin fibroblasts in the 52th generation
3 讨论
本研究参考白剑等[23]的酶消化法分离得到的细胞确定为树鼩皮肤成纤维细胞。本研究针对细胞培养基进行了改良优化,在基础培养基中加入10%血清、1%双抗、1%非必需氨基酸和1%NaHCO3,但没有添加低血清生长添加物。使用改良后的培养基细胞增殖能力仍然较强,生长状态也较好,即使数十次传代后细胞的生长状态也未出现差异,可提高试验的重复性、准确性和稳定性;此外,还避免了血清未知成分对细胞的损伤,提高了细胞质量和试验效率,一方面节省了细胞培养成本,另一方面减少了人为添加生长因子对细胞生长微环境的改变和影响。
本研究分离所得的细胞生长状态较好,不同代次之间细胞整体形态一致,均为梭形纤维状和多角纤维状结构,细胞间排列紧密,有典型的成纤维细胞特性。这与人及大鼠[25]永生化后的皮肤成纤维细胞形态类似,且与原代细胞没有明显区别。成纤维细胞特异性波形蛋白(Vimentin)鉴定结果证实所获得的细胞为成纤维细胞,与人体皮肤成纤维细胞的鉴定结果[9]一致;永生化基因SV40T检测证实已经将该基因成功导入成纤维细胞,在细胞传代培养过程中,其能够在维持细胞状态的同时无限增殖。永生化的成纤维细胞染色体数为62 条,与滇西亚种中缅树鼩[24]一致,证实所得细胞为树鼩源细胞。树鼩永生化皮肤成纤维细胞的建立为人类皮肤相关疾病的体外研究提供了新材料,也为体内皮肤治疗药物的研究奠定了基础。
目前广泛用于永生化细胞建立的方法主要有病毒转染诱导(包括人乳头瘤病毒、人类疱疹病毒和猴肾病毒SV40 等)、化学致癌物诱导射线、基因自发突变和端粒—端粒酶激活诱导产生。病毒诱导主要是病毒感染细胞后,通过改变细胞基因使细胞度过致死期,从而能够无限增殖,以成功诱导永生化细胞[26]。其中,SV40的应用机制为SV40基因被导入细胞核后,细胞内的抑癌基因P53和pRb及相应蛋白能够与其大T 抗原特定结合域结合,导致抑癌基因无法正常表达,细胞得以无限增殖并出现肿瘤相关的转变[27]。而病毒在感染细胞时,感染效率与细胞种类有极大的关系,慢病毒由于转染效率较高而被广泛应用。化学和射线诱导均是将细胞进行癌变诱导[28],而这个过程会导致细胞出现无法预测的其他基因突变,对构建目的细胞模型造成困扰。自发突变则是在细胞的培养过程中,自发的基因突变引起细胞不断增殖,出现细胞永生化现象,但这一方式随机性极强,突变后的基因型极不稳定,或许已经丢失了细胞的原本特性,用于试验时易出现多种不确定因素。端粒—端粒酶激活主要是通过活化细胞内的端粒酶,使其相对高水平地发挥催化端粒复制和维持端粒长度的功能[29],以保证细胞无限分裂。本研究采用慢病毒包装SV40T基因,在确保细胞基因能够往所需方向突变的基础上,大大提高了目的基因被导入细胞基因组的效率,对细胞的永生化建立起到很好的效果。
皮肤成纤维细胞是结缔组织的重要组成成分,对维持皮肤弹性和韧性具有重要作用。本研究获得的永生化树鼩皮肤成纤维细胞是可以稳定传代的细胞,试验时能够即取即用,省去了每次试验需要重新分离纯化细胞的步骤,并且不同代次间的细胞状态差别较小,试验重复性高,能够有效排除试验结果的偶然性,保证试验数据的可靠性。已有研究表明:树鼩皮肤结构与恒河猴及人类皮肤极为相似[22];但小鼠和人类皮肤结构明显不同,如人类皮肤表皮内陷,出现网脊,表皮和真皮层均较厚,而小鼠网脊不突出,皮层均较薄[30],且小鼠的汗腺皮脂腺不发达。因此,建立树鼩永生化的皮肤成纤维细胞系,并利用该细胞开展人类皮肤疾病的致病机理研究和相关药物研发具有重要意义。
4 结论
转染含SV40T永生化基因后的树鼩皮肤成纤维细胞,经过53 代以上的传代培养仍能维持原有状态,不同代次间细胞整体形态一致且排列紧密,有典型的成纤维细胞特性;细胞增殖能力旺盛且其永生化特性得以稳定表达;细胞的核型鉴定结果证明所得细胞确为树鼩源细胞。本研究成功建立1 株树鼩永生化皮肤成纤维细胞,为人类皮肤相关疾病的体外研究提供了新材料。
