海南南渡江上游海南半鲿繁殖生物学研究
2022-05-25梁智策郭志强申志新蔡杏伟
梁智策,潘 帅,刘 飞,郭志强,申志新,蔡杏伟
(1.海南省海洋与渔业科学院,海口 571126;2.南海海洋资源利用国家重点实验室,海南大学海洋学院,海南大学生命科学学院,海口 570228)
海南半鲿(Tchang)为海南岛珍稀鱼类,隶属于鲇形目(Siluriformes)鲿科(Bagridae)半鲿属()。其濒危等级为数据缺乏(DD)。上个世纪中期,海南半鲿在海南岛南渡江水系、昌化江水系以及万泉河水系均有分布。但随着海南省社会和经济的发展,人们对岛内河流生态环境的干扰不断增大,如河道采砂挖石、河道梯级水电开发、水体污染、酷渔滥捕等,使得河流生境严重破碎化、水域环境污染、自然生境退化甚至丧失。人类活动导致的河流生态环境的一系列变化将会对鱼类产生一定的影响。目前,关于海南半鲿的研究仍停留在形态学分类、分布记录和生物学参数研究等方面,而对其基础生物学及资源动态方面的研究极其匮乏。有关海南半鲿的基础研究严重滞后,相关野外实地调查数据缺乏,导致无法对其受濒危程度进行评估。因此,开展海南半鲿有关基础生物学方面的研究,积累相关方面数据迫在眉睫。
繁殖是鱼类生活史研究的重要内容之一,包括生殖腺的发育、排卵、受精,以及精卵结合孵化仔鱼。鱼类通过摄食等途径为繁殖活动准备物质和能量资源,并将这种资源繁殖传递给后代,以此保证种群可持续繁衍和发展。掌握繁殖生物学信息是研究鱼类种群动态变化的基础之一,是成功进行资源评估和管理的关键。繁殖力反映了物种或种群对环境因子变化的适应能力,关系到种群的补充,其变化体现了环境对种群的影响和种群相对应的适应能力,对于科学有效地渔业管理非常重要。本研究以南渡江上游海南半鲿为研究对象,通过揭示海南半鲿基础繁殖生物学特征,包括性、性腺发育程度、繁殖季节、性成熟年龄和大小以及繁殖力等,旨在为海南南渡江上游海南半鲿的种群资源恢复和保护提供基础资料,从而实现南渡江上游海南半鲿资源的可持续发展。
1 材料与方法
1.1 研究区域与数据获取
南渡江为海南岛最大的河流,发源于白沙黎族自治县南峰山,干流斜贯海南岛中北部,于海口市美兰区流入琼中海峡。南渡江上游山高坡陡,支流众多,生境多样性较高,具有丰富的鱼类种类,是鱼类良好的栖息地。根据历史资料记载,上世纪80年代南渡江共记录有鱼类152种。余梵冬等于2018年调查了南渡江鱼类种类组成及分布,结果表明南渡江鱼类共计136种,保存相对完整。
本研究于2019年1-12月对南渡江上游渔获物按月度进行调查(图1)。每月15-17日在日落之前放置3条多网目复合刺网(长:30 m;高:1.5 m;网目2a分别为:1.0,6.0和12.5 cm)和地笼(长:4 m;网目:2a=0.4 cm),次日6:00收网,收集网内所有的渔获物,并及时放入带有冰块的鱼箱内保存,带回实验室测量每尾样本的体长(,1 mm)和体质量(,0.1 g)之后,解剖,通过性腺发育情况辨别性别。性腺发育到Ⅱ期及以上的样本即可判别性别。取出性腺并称重(,0.001 g),最后称量去内脏和性腺后的体重(,0.1 g)。

图1 南渡江上游海南半鲿采样点布置图
初步发现海南半鲿的左右卵巢是同步发育且大小没有显著差异,因此取左卵巢用于组织学分析。根据组织学和外部特征,六期性腺分期标准用于本研究性腺分期。
1.2 研究方法
1.2.1 体长-体质量关系
其体长与体质量关系式=由幂函数拟合,式中为体质量(g);为体长(mm);为肥满度;为幂指数系数。
1.2.2 性体指标
性体指标(GSI)的计算公式如下:GSI=/。
1.2.3 50%性成熟体长
在渔业资源评估中,一般采用50%性成熟体长作为鱼类性成熟体长的参数,本研究以10 mm为体长组的组距,将用于繁殖生物学特征分析的全部雌性个体用来拟合性成熟曲线并估算50%性成熟体长()。海南半鲿的性腺成熟度百分比与体长组的关系用logistic曲线回归:
=1/[1+e-(-)]
式中:为成熟度增长系数;为50%性成熟体长。
1.2.4 繁殖力估算
成熟(性腺发育至IV期或V期)雌性个体的卵巢用来计算绝对繁殖力()和相对繁殖力()。从右侧卵巢的前部、中部和后部各取约0.3 g卵粒混匀并称重(,0.001 g),保存在5%的福尔马林中。将全部卵粒()计数。绝对繁殖力和相对繁殖力的计算方法如下:
=×/;
=/。
2 结果
2.1 个体大小分布
研究分析了体长()为42~289 mm的491个样本(图2)。在总样本中共鉴定出雌性个体146尾,体长72~208 mm,平均体长为143.42 mm;雄性个体105尾,体长91~289 mm,平均体长为180.54 mm(Mann-Whitney检验,=4 570.5,<0.001);其它为性腺未发育个体。雌性和雄性个体平均体长具有极显著差异,因此海南半鲿具有雌雄异形现象。

图2 南渡江上游海南半鲿的体长分布图
2.2 体长-体质量关系
通过拟合南渡江上游海南半鲿的雌性、雄性和性别未分辨群体的体长和体质量幂指数曲线(图3),其体长和体质量关系回归方程为:(1)全部样本:=0000 024288(=098;=491);(2)雌:=0000 012303(=097;=146);雄:=0000 050273(=098;=105);(3)性别未分辨:=0000 017295(=098;=240)。

图3 南渡江上游海南半鲿雌雄体长与体质量关系图
2.3 性腺组织切片
本研究总样本的雌雄性比为1.39:1,146个雌性样本全部用于GSI分析和组织切片分析。通过观察其卵巢组织,对146尾雌性样本的卵巢发育情况进行分期,具体结果如下:
第II期:不成熟期。性腺比较小,通常有脂肪包裹,肉眼几乎看不到卵粒。组织切片观察发现:卵黄发生前的卵母细胞主要出现在不成熟期的卵巢中。包括周边核仁期(peri-nucleolus;PN),核仁分布在细胞核膜内缘周围(图4-a),产卵板期(图4-b)和单个卵黄囊期(图4-c)。
第Ⅲ期:发育期。性腺稍微膨大,大部分被脂肪包裹,肉眼可见极少数卵粒。组织切片观察发现:卵母细胞中出现滤泡层和多个卵黄囊。包括早期的卵黄囊(图4-d),中期的卵黄囊(图4-e)和晚期的卵黄囊(图4-f)。
第IV期:成熟期。性腺饱满,少量或没有脂肪包裹。肉眼能看到全部卵粒,但是卵粒不透明,挤压腹部几乎没有卵粒从泄殖孔排出。组织切片观察发现:卵母细胞的细胞质中开始出现卵黄颗粒和油球(图4-g),之后卵黄颗粒的大小增大且数量不断增加(图4-h),在末期几乎充满整个细胞质(图4-i)。
第V期:饱满期(图4-j)。此时期特点为:卵巢体积及重量明显增大,手指轻轻按压鱼体腹部,有成熟的卵细胞排出。通过组织切片观察发现:与成熟期相比,卵黄颗粒的数量增加且大小增大,几乎占据了整个卵母细胞。同时,卵母细胞的大小也显著增加。油球的数量减少但是体积显著增大。细胞核逐渐向动物极一端迁移。

图4 南渡江上游海南半鲿性腺不同发育期的卵巢组织学切片
2.4 繁殖时间和50%性成熟体长
GSI值在4-5月迅速上升,最高的GSI均值出现在6月,GSI均值从6月开始逐渐下降(图5)。
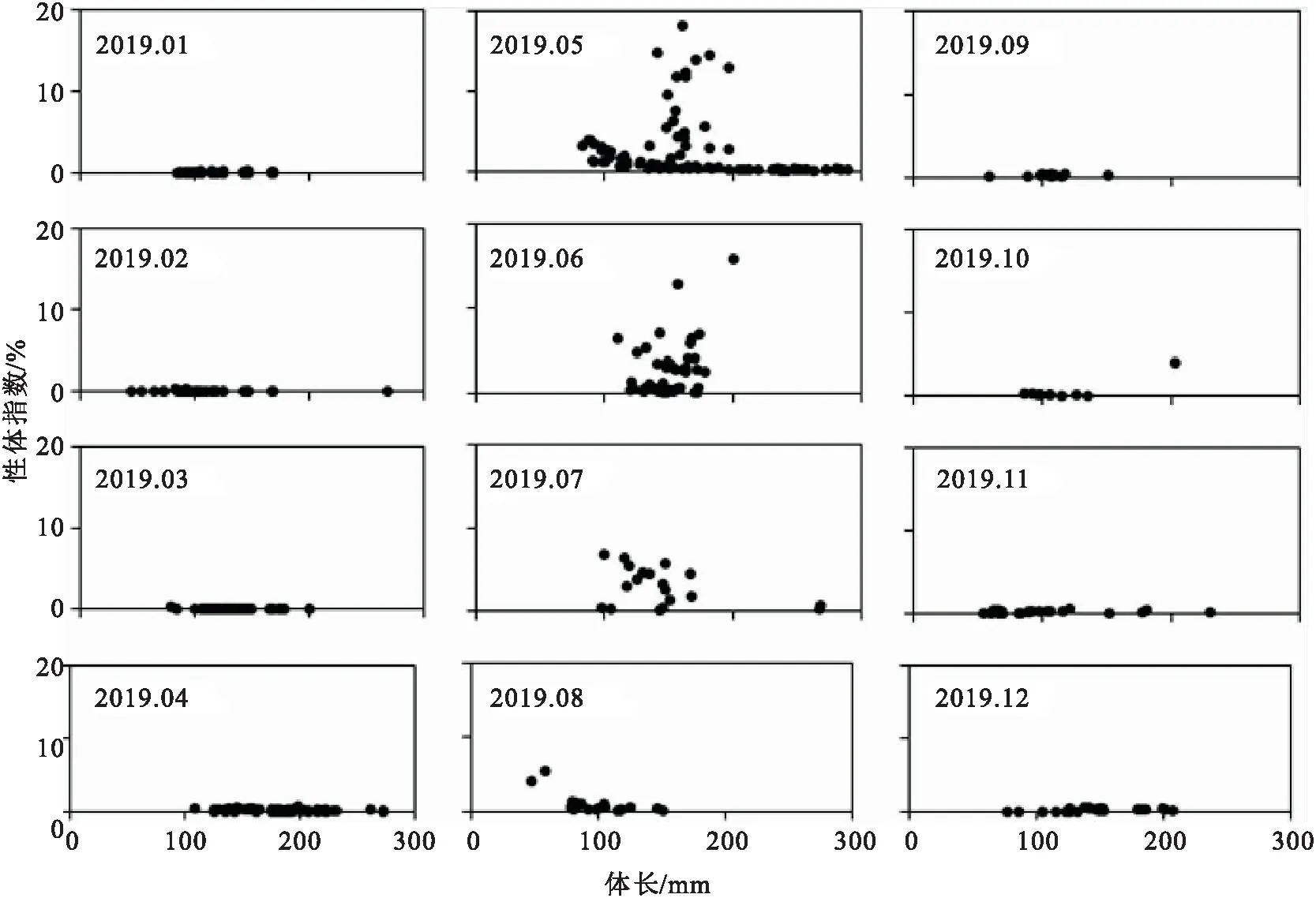
图5 南渡江上游雌性海南半鲿GSI值的月变化
南渡江上游海南半鲿为140.95 mm,通过logistic曲线回归法估算而得。海南半鲿的性腺成熟度百分比与体长组的关系为:=1/[1+e-0.093 9(-140.95)](=0.986;图6)。

图6 南渡江上游雌性海南半鲿性成熟比例与体长关系
2.5 繁殖力
本研究以未受精成熟卵和发育期受精卵为数据源,统计怀卵量。共采集到23尾海南半鲿成熟个体(性腺发育阶段处于Ⅳ期和Ⅴ期),体长为125~199 mm,体质量为33.56~129.13 g。海南半鲿绝对繁殖力波动于20~1 157粒/尾,平均(511±339)粒/尾;相对繁殖力波动于0.66~30.91粒/g,平均(6.89±4.26)粒/g。
拟合南渡江上游海南半鲿的绝对繁殖力和相对繁殖力与体长和体质量的关系,结果显示,海南半鲿的绝对繁殖力与体长和体质量呈现正相关性,拟合方程如下:=10.741-1 300.7(=0.394;<0.01);=9.035-185.94(=0.481 8;<0.01),而相对繁殖力在一定范围内波动,并无明显的变化趋势。
2.6 成熟卵径
对23个Ⅳ期和Ⅴ期卵巢样本的卵径进行测量,结果显示南渡江上游海南半鲿的卵粒较大,卵径分布范围较广,由图7可知,海南半鲿的卵径分布为单峰型,海南半鲿为一次性产卵鱼类。

图7 南渡江上游海南半鲿卵径的分布
3 讨论
3.1 雌雄异形
本研究中海南半鲿表现出明显的雌雄异形现象,雄性群体的最大个体和平均个体大小均高于雌性群体。雄性个体大于雌性的雌雄异形现象主要来源于性别选择。当大个体雄性吸引的雌性个体数量大于小个体雄性时,个体的增大会增加雄性繁殖的成功率,称为配偶选择;当大个体雄性具有更强的竞争能力,包括更强的筑巢、护巢以及护幼能力时,这些雄性个体也会获得更高的繁殖成功率,称为同性选择。半鲿属鱼类具有挖巢、护巢和护幼行为,并由雄性个体负责,直到巢内孵化的仔鱼能自由游动为止,大个体的雄性将更有利于掘巢及护幼等行为,从而增加仔鱼的成活率。我们推断同性选择是海南半鲿雌雄异形的主要成因。
3.2 繁殖特征
南渡江上游海南半鲿具有高GSI值(大于0.02),成熟雌性个体(性腺发育处于IV期和V期)以及产后雌性个体(性腺发育处于VI期)主要出现在5月至7月。并且根据卵巢组织学的观察,性腺发育处于IV期和V期的成熟雌性个体在5月和6月所占的比例较高,因此,南渡江上游海南半鲿的繁殖高峰期为5-6月。王德寿等和朱成科等根据大鳍半鲿()性腺、成熟系数的周年变化,推断嘉陵江水系和长江水系大鳍半鲿的繁殖季节均为5-7月,繁殖高峰期为6-7月。而杜孝田等认为元江多辐半鲿繁殖期为4-7月,高峰期为5月。另外,阴双雨研究发现澜沧江丝尾半鲿()性体指标具有两个高峰(4月和8月),预示丝尾半鲿可进行分批产卵。造成这种差异的原因可能是栖息地环境(水温、流速、水质等)不同导致繁殖活动所需外源因子的到达时间不同,导致同一鱼类不同地理种群的繁殖时间发生变化。
性比是种群繁殖力的重要决定因素之一,体现了物种对环境长期适应性。澜沧江丝尾半鲿、元江多辐半鲿()和涪江、渠江大鳍半鲿的雌性个体数均多于雄性,而对于大鳍半鲿,王德寿等和朱成科等认为其雌性个体数远小于雄性,本研究中南渡江上游海南半鲿雌性个体显著多于雄性个体。造成这种差异可能是由于生境差异化导致的,因为鱼类在其他条件相似的情况下,雌性占优势是维持和增加种群数量的主要手段之一。
在渔业资源评估中,鱼类繁殖力体现了鱼类的繁殖潜能。绝对繁殖力反映了物种或种群对环境变化的适应特征,而相对繁殖力则反映了鱼类的繁殖策略。南渡江上游海南半鲿的绝对繁殖力()平均值为(511±339)粒/尾,相对繁殖力()平均值为(6.89±4.26)粒/g。本研究的相对繁殖力比报道的其他半鲿属鱼类都要低,而卵径则普遍大于其他半鲿属鱼类。相对较低的繁殖力意味着鱼所怀的卵较大,数量较少,含有较多的卵黄,因此孵化的仔鱼可以利用较多的营养,从而增加了每个卵发育成成鱼的机会。另外,环境因子是造成鱼类繁殖力变化的关键因素之一,在众多环境因子中,最重要的因子为营养条件。生境食饵丰度大,鱼类体长和体质量生长速率快,相对丰满度大,性腺发育快,繁殖力高;相反,性腺发育不良,萎缩,甚至退化、最后被吸收,繁殖力就会下降。目前,南渡江保存了历史大部分鱼类,河流整体健康状况良好,但鱼类总种数相较20世纪80年代下降了10.5%。推测威胁南渡江上游海南半鲿种群生存、繁殖的因素可能有以下两方面:(1)水利工程建设是威胁海南岛淡水鱼类多样性的最大因素,南渡江上游建有18个小型滚水坝,可能造成海南半鲿生境碎片化,影响其产卵、摄食场所,降低种群整体的遗传多样性。(2)外来鱼类入侵是导致本土鱼类种群丰度下降的最大因素,南渡江上游齐氏罗非鱼()、花身副丽鱼()等外来入侵物种的大量出现,会与海南半鲿形成生态位重叠,竞争食饵,甚至直接捕食海南半鲿的鱼卵及幼鱼等,对其生存繁殖造成严重的威胁。鱼类繁殖力变动的主要原因之一为生境条件的改变。这种发生在自然环境下的繁殖策略变化需要进一步监测,并与资源评估相结合,这将有利于了解鱼类种群结构的变动趋势。
